
Potential of Dietary HDAC2i in Breast Cancer Patients Receiving PD-1/PD-L1 Inhibitors

Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
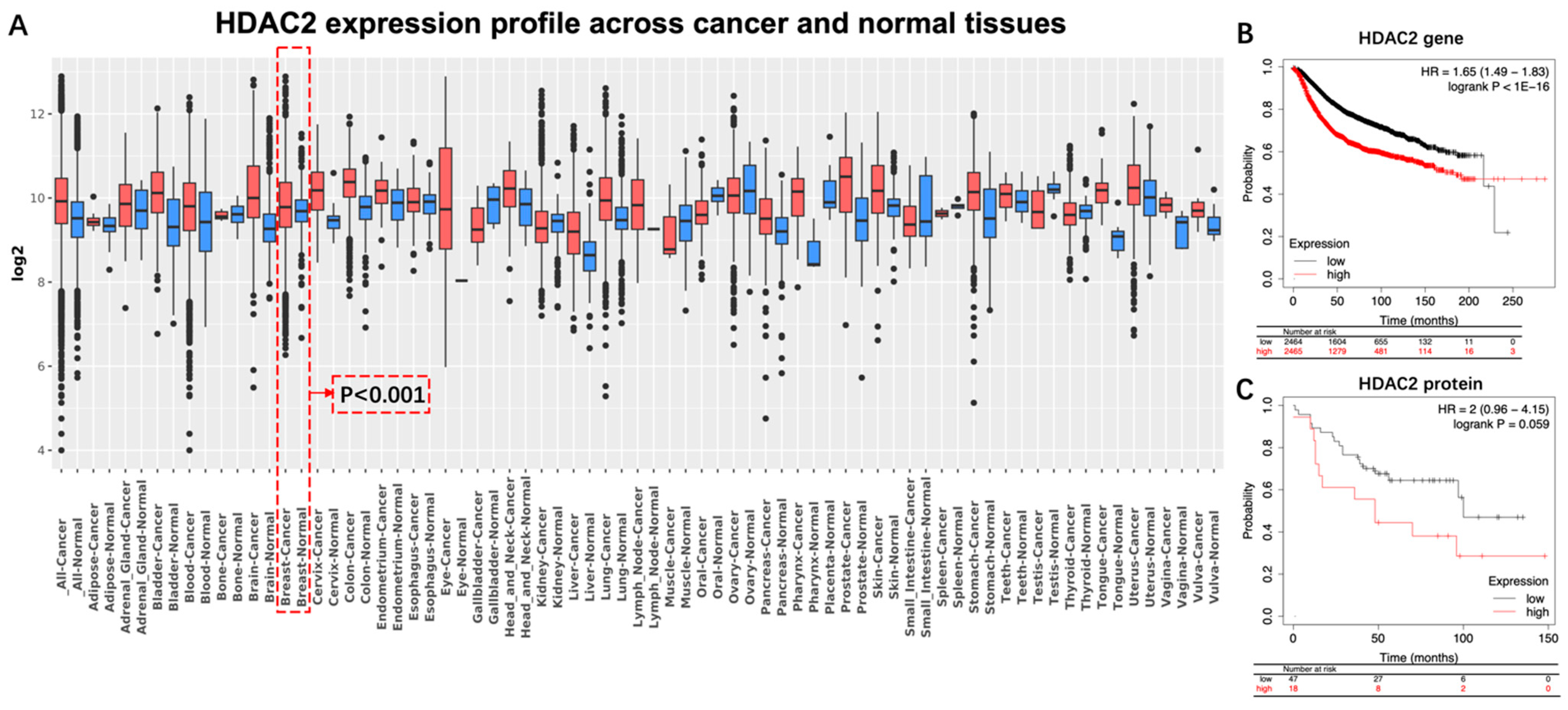
4.1. HDAC2: A Potential Index of Aggressiveness and a Therapeutic Target against BC
4.2. HDAC2 Inhibition for Treating Breast Cancer
4.3. HDAC2 Inhibition Enhances the Therapeutic Effect of ICIs in BC Treatment
4.3.1. HDAC2 Regulates PD-L1 Nuclear Translocation
4.3.2. HDAC2 Regulates IFN-γ- Induced PD-L1 Expression
- a.
- IFN-γ upregulates the expression of PD-L1
- b.
- HDAC2i affects IFN-γ induced PD-L1
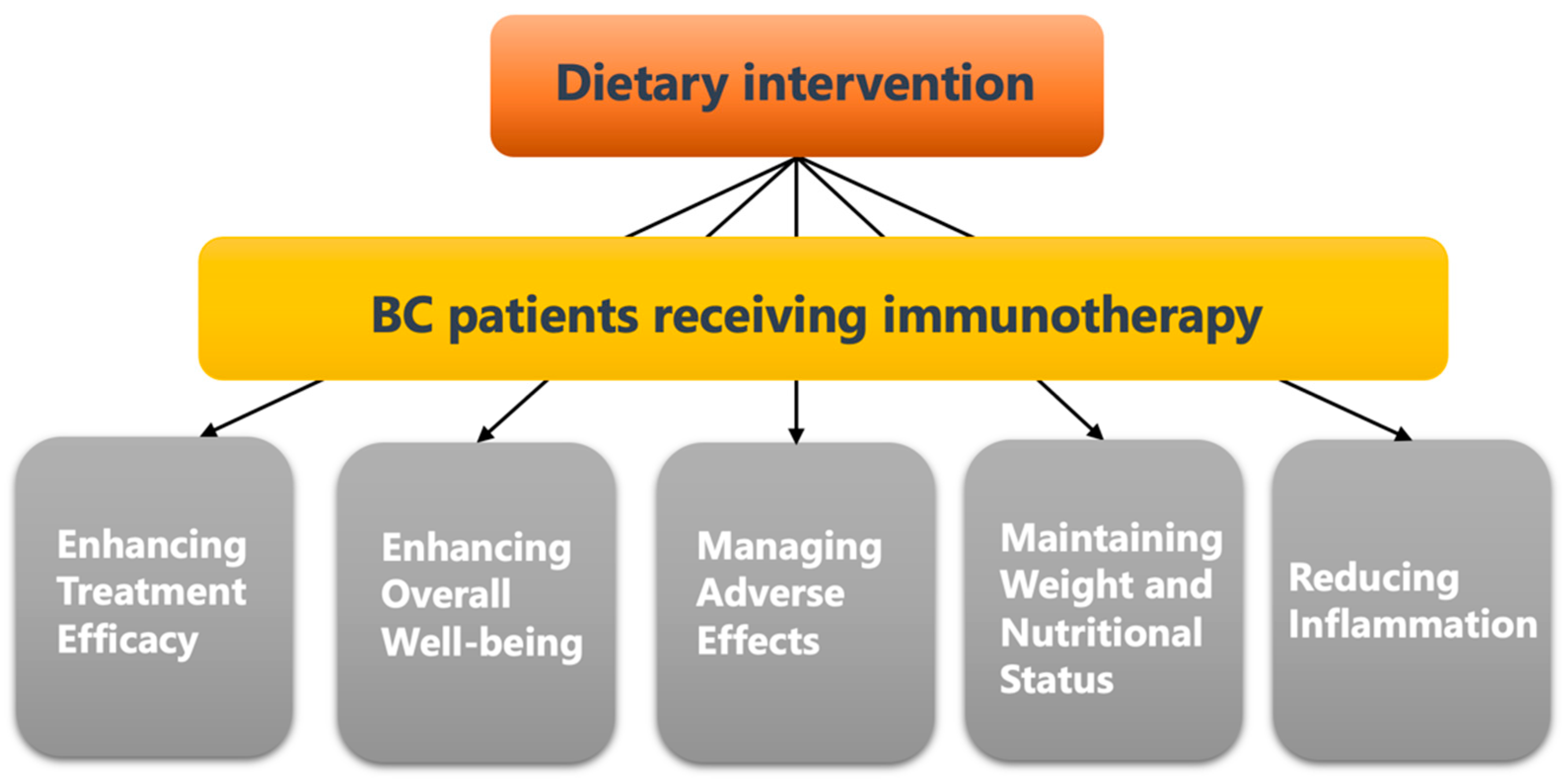
4.4. Dietary Intervention Is Important for Breast Cancer Patients Receiving Anti-Cancer Immunotherapy
4.5. Dietary HDAC2i
4.6. Selected Candidates of HDAC2i
4.6.1. Genistein (GE)
4.6.2. Sulforaphane (SFN)
4.6.3. Chrysin and Its Analogues
4.6.4. Resveratrol (RSV)
4.6.5. Oleuropein (OLE)
4.6.6. Curcumin
4.6.7. Valeric Acid
4.6.8. Rh4
4.6.9. Butyrate (NaB)
4.6.10. Other Potential Candidates
4.7. Potential Approaches of Taking Bioactive Compound
- a.
- Nutraceuticals and Dietary Supplements
- b.
- Nanotechnology and Drug Delivery
- c.
- Pharmaceuticals and Medicinal Products
- d.
- Phytotherapy and Traditional Medicine
4.8. Nutrients That May Impair the Therapeutic Effect of ICIs
- a.
- Omega-3 Fatty Acids
- b.
- Vitamins
- c.
- Probiotics
- d.
- High-Fiber Diets
- e.
- Ketogenic diet
- f.
- Protein-restricted diet
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tao, X.; Li, T.; Gandomkar, Z.; Brennan, P.C.; Reed, W.M. Incidence, mortality, survival, and disease burden of breast cancer in China compared to other developed countries. Asia-Pac. J. Clin. Oncol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, T.; Al-Husseini, M.; Saravolatz, L.; Kafri, Z. Trends in Breast Cancer Incidence and Mortality in the United States From 2004-2018: A Surveillance, Epidemiology, and End Results (SEER)-Based Study. Cureus 2023, 15, e37982. [Google Scholar] [CrossRef] [PubMed]
- Pedersini, R.; di Mauro, P.; Bosio, S.; Zanini, B.; Zanini, A.; Amoroso, V.; Turla, A.; Vassalli, L.; Ardine, M.; Monteverdi, S.; et al. Changes in eating habits and food preferences in breast cancer patients undergoing adjuvant chemotherapy. Sci. Rep. 2021, 11, 12975. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; McGale, P.; Probert, J.; Broggio, J.; Charman, J.; Darby, S.C.; Kerr, A.J.; Whelan, T.; Cutter, D.J.; Mannu, G.; et al. Breast cancer mortality in 500 000 women with early invasive breast cancer in England, 1993–2015: Population based observational cohort study. BMJ 2023, 381, e074684. [Google Scholar] [CrossRef]
- Bertucci, F.; Gonçalves, A. Immunotherapy in Breast Cancer: The Emerging Role of PD-1 and PD-L1. Curr. Oncol. Rep. 2017, 19, 64. [Google Scholar] [CrossRef]
- Lu, L.; Bai, Y.; Wang, Z. Elevated T cell activation score is associated with improved survival of breast cancer. Breast Cancer Res. Treat. 2017, 164, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.L.; Huppert, L.A.; Rugo, H.S. Role of Immunotherapy in Breast Cancer. JCO Oncol. Pract. 2023, 19, 167–179. [Google Scholar] [CrossRef]
- Hattori, M.; Masuda, N.; Takano, T.; Tsugawa, K.; Inoue, K.; Matsumoto, K.; Ishikawa, T.; Itoh, M.; Yasojima, H.; Tanabe, Y.; et al. Pembrolizumab plus chemotherapy in Japanese patients with triple-negative breast cancer: Results from KEYNOTE-355. Cancer Med. 2023, 12, 10280–10293. [Google Scholar] [CrossRef]
- Downs-Canner, S.; Mittendorf, E.A. Correction: Preoperative Immunotherapy Combined with Chemotherapy for Triple-Negative Breast Cancer: Perspective on the KEYNOTE-522 Study. Ann. Surg. Oncol. 2023, 30, 3286. [Google Scholar] [CrossRef]
- Khalid, A.B.; Calderon, G.; Jalal, S.I.; Durm, G.A. Physician Awareness of Immune-Related Adverse Events of Immune Checkpoint Inhibitors. Breast Cancer Res. Treat 2022, 20, 1316–1320. [Google Scholar] [CrossRef]
- Gumusay, O.; Callan, J.; Rugo, H.S. Immunotherapy toxicity: Identification and management. Breast Cancer Res. Treat. 2022, 192, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zhao, S.; Dong, Y.; Wang, T.; Niu, X.; Zhao, L.; Wang, G. Antitumor activity and mechanism of resistance of the novel HDAC and PI3K dual inhibitor CUDC-907 in pancreatic cancer. Cancer Chemother. Pharmacol. 2021, 87, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Garmpis, N.; Damaskos, C.; Dimitroulis, D.; Kouraklis, G.; Garmpi, A.; Sarantis, P.; Koustas, E.; Patsouras, A.; Psilopatis, I.; Antoniou, E.A.; et al. Clinical Significance of the Histone Deacetylase 2 (HDAC-2) Expression in Human Breast Cancer. J. Pers. Med. 2022, 12, 1672. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Jiang, Y.; Yu, H.; Huang, Q.; Liu, L.; Guo, X.; Li, L.; Mi, Q.; Zhang, K.; Yang, Z. HDAC2 overexpression correlates with aggressive clinicopathological features and DNA-damage response pathway of breast cancer. Am. J. Cancer Res. 2017, 7, 1213–1226. [Google Scholar]
- Zhao, H.; Yu, Z.; Zhao, L.; He, M.; Ren, J.; Wu, H.; Chen, Q.; Yao, W.; Wei, M. HDAC2 overexpression is a poor prognostic factor of breast cancer patients with increased multidrug resistance-associated protein expression who received anthracyclines therapy. Jpn. J. Clin. Oncol. 2016, 46, 893–902. [Google Scholar] [CrossRef]
- Maccallini, C.; Ammazzalorso, A.; De Filippis, B.; Fantacuzzi, M.; Giampietro, L.; Amoroso, R. HDAC Inhibitors for the Therapy of Triple Negative Breast Cancer. Pharmaceuticals 2022, 15, 667. [Google Scholar] [CrossRef]
- Li, Y.; Seto, E. HDACs and HDAC Inhibitors in Cancer Development and Therapy. Cold Spring Harb. Perspect. Med. 2016, 6, a026831. [Google Scholar] [CrossRef]
- Xu, P.; Xiong, W.; Lin, Y.; Fan, L.; Pan, H.; Li, Y. Histone deacetylase 2 knockout suppresses immune escape of triple-negative breast cancer cells via downregulating PD-L1 expression. Cell Death Dis. 2021, 12, 779. [Google Scholar] [CrossRef]
- Gao, Y.; Nihira, N.T.; Bu, X.; Chu, C.; Zhang, J.; Kolodziejczyk, A.; Fan, Y.; Chan, N.T.; Ma, L.; Liu, J.; et al. Acetylation-dependent regulation of PD-L1 nuclear translocation dictates the efficacy of anti-PD-1 immunotherapy. Nat. Cell Biol. 2020, 22, 1064–1075. [Google Scholar] [CrossRef]
- Bassett, S.A.; Barnett, M.P.G. The Role of Dietary Histone Deacetylases (HDACs) Inhibitors in Health and Disease. Nutrients 2014, 6, 4273–4301. [Google Scholar] [CrossRef]
- Zhu, G.; Su, H.; Johnson, C.H.; Khan, S.A.; Kluger, H.; Lu, L. Intratumour microbiome associated with the infiltration of cytotoxic CD8+ T cells and patient survival in cutaneous melanoma. Eur. J. Cancer 2021, 151, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Limon-Miro, A.T.; Lopez-Teros, V.; Astiazaran-Garcia, H. Dietary Guidelines for Breast Cancer Patients: A Critical Review. Adv. Nutr. Int. Rev. J. 2017, 8, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.; Min, S.K.; Park, H.-R.; Kim, D.H.; Kwon, M.J.; Kim, L.S.; Ju, Y.-S. Expression of Histone Deacetylases HDAC1, HDAC2, HDAC3, and HDAC6 in Invasive Ductal Carcinomas of the Breast. J. Breast Cancer 2014, 17, 323–331. [Google Scholar] [CrossRef]
- Muller, B.M.; Jana, L.; Kasajima, A.; Lehmann, A.; Prinzler, J.; Budczies, J.; Winzer, K.-J.; Dietel, M.; Weichert, W.; Denkert, C. Differential expression of histone deacetylases HDAC1, 2 and 3 in human breast cancer—Overexpression of HDAC2 and HDAC3 is associated with clinicopathological indicators of disease progression. BMC Cancer 2013, 13, 215. [Google Scholar] [CrossRef]
- Long, M.; Hou, W.; Liu, Y.; Hu, T. A Histone Acetylation Modulator Gene Signature for Classification and Prognosis of Breast Cancer. Curr. Oncol. 2021, 28, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.R.; Hwang, C.Y.; Lee, J.; Cho, K.-H. Network Analysis Identifies Regulators of Basal-Like Breast Cancer Reprogramming and Endocrine Therapy Vulnerability. Cancer Res. 2022, 82, 320–333. [Google Scholar] [CrossRef]
- Harms, K.L.; Chen, X. Histone Deacetylase 2 Modulates p53 Transcriptional Activities through Regulation of p53-DNA Binding Activity. Cancer Res. 2007, 67, 3145–3152. [Google Scholar] [CrossRef]
- He, X.-H.; Zhu, W.; Yuan, P.; Jiang, S.; Li, D.; Zhang, H.-W.; Liu, M.-F. miR-155 downregulates ErbB2 and suppresses ErbB2-induced malignant transformation of breast epithelial cells. Oncogene 2016, 35, 6015–6025. [Google Scholar] [CrossRef]
- Jo, H.; Shim, K.; Kim, H.-U.; Jung, H.S.; Jeoung, D. HDAC2 as a target for developing anti-cancer drugs. Comput. Struct. Biotechnol. J. 2023, 21, 2048–2057. [Google Scholar] [CrossRef]
- Biçaku, E.; Marchion, D.C.; Schmitt, M.L.; Münster, P.N. Selective Inhibition of Histone Deacetylase 2 Silences Progesterone Receptor–Mediated Signaling. Cancer Res. 2008, 68, 1513–1519. [Google Scholar] [CrossRef]
- Marchion, D.C.; Bicaku, E.; Turner, J.G.; Schmitt, M.L.; Morelli, D.R.; Munster, P.N. HDAC2 regulates chromatin plasticity and enhances DNA vulnerability. Mol. Cancer Ther. 2009, 8, 794–801. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qiu, N.; Yin, J.; Zhang, J.; Liu, H.; Guo, W.; Liu, M.; Liu, T.; Chen, D.; Luo, K.; et al. SRGN crosstalks with YAP to maintain chemoresistance and stemness in breast cancer cells by modulating HDAC2 expression. Theranostics 2020, 10, 4290–4307. [Google Scholar] [CrossRef] [PubMed]
- Masoumi, E.; Tahaghoghi-Hajghorbani, S.; Jafarzadeh, L.; Sanaei, M.-J.; Pourbagheri-Sigaroodi, A.; Bashash, D. The application of immune checkpoint blockade in breast cancer and the emerging role of nanoparticle. J. Control. Release 2021, 340, 168–187. [Google Scholar] [CrossRef] [PubMed]
- Isaacs, J.; Anders, C.; McArthur, H.; Force, J. Biomarkers of Immune Checkpoint Blockade Response in Triple-Negative Breast Cancer. Curr. Treat. Options Oncol. 2021, 22, 38. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Risch, E.; Halaban, R.; Zhen, P.; Bacchiocchi, A.; Risch, H.A. Dynamic changes of circulating soluble PD-1/PD-L1 and its association with patient survival in immune checkpoint blockade-treated melanoma. Int. Immunopharmacol. 2023, 118, 110092. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.; Wu, C.; Chen, M.; Jiang, Y.; Wu, Y.; Mao, R.; Fan, Y. The generation of PD-L1 and PD-L2 in cancer cells: From nuclear chromatin reorganization to extracellular presentation. Acta Pharm. Sin. B 2022, 12, 1041–1053. [Google Scholar] [CrossRef]
- Xiong, W.; Gao, Y.; Wei, W.; Zhang, J. Extracellular and nuclear PD-L1 in modulating cancer immunotherapy. Trends Cancer 2021, 7, 837–846. [Google Scholar] [CrossRef]
- Koh, Y.W.; Han, J.-H.; Haam, S.; Lee, H.W. HIP1R Expression and Its Association with PD-1 Pathway Blockade Response in Refractory Advanced NonSmall Cell Lung Cancer: A Gene Set Enrichment Analysis. J. Clin. Med. 2020, 9, 1425. [Google Scholar] [CrossRef]
- Muenst, S.; Schaerli, A.R.; Gao, F.; Däster, S.; Trella, E.; Droeser, R.A.; Muraro, M.G.; Zajac, P.; Zanetti, R.; Gillanders, W.E.; et al. Expression of programmed death ligand 1 (PD-L1) is associated with poor prognosis in human breast cancer. Breast Cancer Res. Treat. 2014, 146, 15–24. [Google Scholar] [CrossRef]
- Lotfinejad, P.; Kazemi, T.; Safaei, S.; Amini, M.; Baghbani, E.; Shotorbani, S.S.; Niaragh, F.J.; Derakhshani, A.; Shadbad, M.A.; Silvestris, N. PD-L1 silencing inhibits triple-negative breast cancer development and upregulates T-cell-induced pro-inflammatory cytokines. Biomed. Pharmacother. 2021, 138, 111436. [Google Scholar] [CrossRef]
- Setordzi, P.; Chang, X.; Liu, Z.; Wu, Y.; Zuo, D. The recent advances of PD-1 and PD-L1 checkpoint signaling inhibition for breast cancer immunotherapy. Eur. J. Pharmacol. 2021, 895, 173867. [Google Scholar] [CrossRef] [PubMed]
- Schmid, P.; Adams, S.; Rugo, H.S.; Schneeweiss, A.; Barrios, C.H.; Iwata, H.; Diéras, V.; Hegg, R.; Im, S.-A.; Shaw Wright, G.; et al. Atezolizumab and Nab-Paclitaxel in Advanced Triple-Negative Breast Cancer. N. Engl. J. Med. 2018, 379, 2108–2121. [Google Scholar] [CrossRef] [PubMed]
- Minn, A.J.; Wherry, E.J. Combination Cancer Therapies with Immune Checkpoint Blockade: Convergence on Interferon Signaling. Cell 2016, 165, 272–275. [Google Scholar] [CrossRef] [PubMed]
- Abiko, K.; Matsumura, N.; Hamanishi, J.; Horikawa, N.; Murakami, R.; Yamaguchi, K.; Yoshioka, Y.; Baba, T.; Konishi, I.; Mandai, M. IFN-γ from lymphocytes induces PD-L1 expression and promotes progression of ovarian cancer. Br. J. Cancer 2015, 112, 1501–1509. [Google Scholar] [CrossRef] [PubMed]
- Bouhet, S.; Lafont, V.; Billard, E.; Gross, A.; Dornand, J. The IFNgamma-induced STAT1-CBP/P300 association, required for a normal response to the cytokine, is disrupted in Brucella-infected macrophages. Microb. Pathog. 2009, 46, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, Y.; Mimura, K.; Tamaki, T.; Shiraishi, K.; Kua, L.-F.; Koh, V.; Ohmori, M.; Kimura, A.; Inoue, S.; Okayama, H.; et al. Phospho-STAT1 expression as a potential biomarker for anti-PD-1/anti-PD-L1 immunotherapy for breast cancer. Int. J. Oncol. 2019, 54, 2030–2038. [Google Scholar] [CrossRef]
- Calder, P.C. Foods to deliver immune-supporting nutrients. Curr. Opin. Food Sci. 2022, 43, 136–145. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.G.; Gombart, A.F.; Eggersdorfer, M. Reply to Comment on: Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef]
- Chen, O.; Mah, E.; Dioum, E.; Marwaha, A.; Shanmugam, S.; Malleshi, N.; Sudha, V.; Gayathri, R.; Unnikrishnan, R.; Anjana, R.M.; et al. The Role of Oat Nutrients in the Immune System: A Narrative Review. Nutrients 2021, 13, 1048. [Google Scholar] [CrossRef]
- Noor, S.; Piscopo, S.; Gasmi, A. Nutrients Interaction with the Immune System. Arch. Razi. Inst. 2021, 76, 1579–1588. [Google Scholar] [CrossRef]
- Kichloo, A.; Albosta, M.; Dahiya, D.; Guidi, J.C.; Aljadah, M.; Singh, J.; Shaka, H.; Wani, F.; Kumar, A.; Lekkala, M. Systemic adverse effects and toxicities associated with immunotherapy: A review. World J. Clin. Oncol. 2021, 12, 150–163. [Google Scholar] [CrossRef]
- Ticinesi, A.; Nouvenne, A.; Chiussi, G.; Castaldo, G.; Guerra, A.; Meschi, T. Calcium Oxalate Nephrolithiasis and Gut Microbiota: Not just a Gut-Kidney Axis. A Nutritional Perspective. Nutrients 2020, 12, 548. [Google Scholar] [CrossRef]
- Taraszewska, A. Risk factors for gastroesophageal reflux disease symptoms related to lifestyle and diet. Rocz Panstw Zakl Hig 2021, 72, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Jadhav, A.; Bajaj, A.; Xiao, Y.; Markandey, M.; Ahuja, V.; Kashyap, P.C. Role of Diet–Microbiome Interaction in Gastrointestinal Disorders and Strategies to Modulate Them with Microbiome-Targeted Therapies. Annu. Rev. Nutr. 2023, 43, 355–383. [Google Scholar] [CrossRef]
- Campmans-Kuijpers, M.J.E.; Dijkstra, G. Food and Food Groups in Inflammatory Bowel Disease (IBD): The Design of the Groningen Anti-Inflammatory Diet (GrAID). Nutrients 2021, 13, 1067. [Google Scholar] [CrossRef] [PubMed]
- Tolkien, K.; Bradburn, S.; Murgatroyd, C. An anti-inflammatory diet as a potential intervention for depressive disorders: A systematic review and meta-analysis. Clin. Nutr. 2019, 38, 2045–2052. [Google Scholar] [CrossRef] [PubMed]
- De Groot, S.; Lugtenberg, R.T.; Cohen, D.; Welters, M.J.P.; Ehsan, I.; Vreeswijk, M.P.G.; Smit, V.T.H.B.M.; de Graaf, H.; Heijns, J.B.; Portielje, J.E.A.; et al. Fasting mimicking diet as an adjunct to neoadjuvant chemotherapy for breast cancer in the multicentre randomized phase 2 DIRECT trial. Nat. Commun. 2020, 11, 3083. [Google Scholar] [CrossRef] [PubMed]
- Arends, J.; Baracos, V.; Bertz, H.; Bozzetti, F.; Calder, P.C.; Deutz, N.E.P.; Erickson, N.; Laviano, A.; Lisanti, M.P.; Lobo, D.N.; et al. ESPEN expert group recommendations for action against cancer-related malnutrition. Clin. Nutr. 2017, 36, 1187–1196. [Google Scholar] [CrossRef]
- Cheng, Y.; Zhang, J.; Zhang, L.; Wu, J.; Zhan, Z. Enteral immunonutrition versus enteral nutrition for gastric cancer patients undergoing a total gastrectomy: A systematic review and meta-analysis. BMC Gastroenterol. 2018, 18, 11. [Google Scholar] [CrossRef]
- Greathouse, K.L.; Wyatt, M.; Johnson, A.J.; Toy, E.P.; Khan, J.M.; Dunn, K.; Clegg, D.J.; Reddy, S. Diet-microbiome interactions in cancer treatment: Opportunities and challenges for precision nutrition in cancer. Neoplasia 2022, 29, 100800. [Google Scholar] [CrossRef]
- Owen, L.; Corfe, B. The role of diet and nutrition on mental health and wellbeing. Proc. Nutr. Soc. 2017, 76, 425–426. [Google Scholar] [CrossRef] [PubMed]
- Berding, K.; Vlckova, K.; Marx, W.; Schellekens, H.; Stanton, C.; Clarke, G.; Jacka, F.; Dinan, T.G.; Cryan, J.F. Diet and the Microbiota–Gut–Brain Axis: Sowing the Seeds of Good Mental Health. Adv. Nutr. Int. Rev. J. 2021, 12, 1239–1285. [Google Scholar] [CrossRef] [PubMed]
- Carreira, H.; Williams, R.; Müller, M.; Harewood, R.; Stanway, S.; Bhaskaran, K. Associations Between Breast Cancer Survivorship and Adverse Mental Health Outcomes: A Systematic Review. J. Natl. Cancer Inst. 2018, 110, 1311–1327. [Google Scholar] [CrossRef] [PubMed]
- Fanakidou, I.; Zyga, S.; Alikari, V.; Tsironi, M.; Stathoulis, J.; Theofilou, P. Mental health, loneliness, and illness perception outcomes in quality of life among young breast cancer patients after mastectomy: The role of breast reconstruction. Qual. Life Res. 2018, 27, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Garay, C.; Djouder, N. Dietary interventions and precision nutrition in cancer therapy. Trends Mol. Med. 2023, 29, 489–511. [Google Scholar] [CrossRef]
- Vernieri, C.; Casola, S.; Foiani, M.; Pietrantonio, F.; De Braud, F.; Longo, V. Targeting Cancer Metabolism: Dietary and Pharmacologic Interventions. Cancer Discov. 2016, 6, 1315–1333. [Google Scholar] [CrossRef]
- Bose, S.; Allen, A.E.; Locasale, J.W. The Molecular Link from Diet to Cancer Cell Metabolism. Mol. Cell 2020, 78, 1034–1044. [Google Scholar] [CrossRef]
- Locasale, J.W. Diet and Exercise in Cancer Metabolism. Cancer Discov. 2022, 12, 2249–2257. [Google Scholar] [CrossRef]
- Morita, M.; Kudo, K.; Shima, H.; Tanuma, N. Dietary intervention as a therapeutic for cancer. Cancer Sci. 2021, 112, 498–504. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Das, S.; Patra, S.K.; Efferth, T.; Jena, M.; Bhutia, S.K. Dietary polyphenols in chemoprevention and synergistic effect in cancer: Clinical evidences and molecular mechanisms of action. Phytomedicine 2021, 90, 153554. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, H. Neurological adverse events associated with PD-1/PD-L1 immune checkpoint inhibitors. Front. Neurosci. 2023, 17, 1227049. [Google Scholar] [CrossRef] [PubMed]
- Alturki, N.A. Review of the Immune Checkpoint Inhibitors in the Context of Cancer Treatment. J. Clin. Med. 2023, 12, 4301. [Google Scholar] [CrossRef] [PubMed]
- Zraik, I.M.; Heß-Busch, Y. Management of chemotherapy side effects and their long-term sequelae. Urologe A 2021, 60, 862–871. [Google Scholar] [CrossRef]
- Buchholz, T.A. Radiation Therapy for Early-Stage Breast Cancer after Breast-Conserving Surgery. New Engl. J. Med. 2009, 360, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Xu, C.; Gao, X.; Yao, Q. Platinum-based drugs for cancer therapy and anti-tumor strategies. Theranostics 2022, 12, 2115–2132. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Gao, Z.-J.; Yu, X.; Wang, P. Dietary regulation in health and disease. Signal Transduct. Target. Ther. 2022, 7, 252. [Google Scholar] [CrossRef]
- Prado, C.M.; Antoun, S.; Sawyer, M.B.; Baracos, V.E. Two faces of drug therapy in cancer: Drug-related lean tissue loss and its adverse consequences to survival and toxicity. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 250–254. [Google Scholar] [CrossRef]
- Soldati, L.; Di Renzo, L.; Jirillo, E.; Ascierto, P.A.; Marincola, F.M.; De Lorenzo, A. The influence of diet on anti-cancer immune responsiveness. J. Transl. Med. 2018, 16, 75. [Google Scholar] [CrossRef]
- Tao, J.; Li, S.; Gan, R.-Y.; Zhao, C.-N.; Meng, X.; Li, H.-B. Targeting gut microbiota with dietary components on cancer: Effects and potential mechanisms of action. Crit. Rev. Food Sci. Nutr. 2020, 60, 1025–1037. [Google Scholar] [CrossRef]
- Dieli-Conwright, C.M.; Harrigan, M.; Cartmel, B.; Chagpar, A.; Bai, Y.; Li, F.-Y.; Rimm, D.L.; Pusztai, L.; Lu, L.; Sanft, T.; et al. Impact of a randomized weight loss trial on breast tissue markers in breast cancer survivors. npj Breast Cancer 2022, 8, 29. [Google Scholar] [CrossRef]
- Puklin, L.; Cartmel, B.; Harrigan, M.; Lu, L.; Li, F.-Y.; Sanft, T.; Irwin, M.L. Randomized trial of weight loss on circulating ghrelin levels among breast cancer survivors. npj Breast Cancer 2021, 7, 49. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.A.; Alvarez-Reeves, M.; Lu, L.; Yu, H.; Irwin, M.L. Effect of Exercise on Metabolic Syndrome Variables in Breast Cancer Survivors. Int. J. Endocrinol. 2013, 2013, 168797. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Irwin, M.L.; Lu, L.; Risch, H.; Mayne, S.; Mu, L.; Deng, Q.; Scarampi, L.; Mitidieri, M.; Katsaros, D.; et al. Physical activity and breast cancer survival: An epigenetic link through reduced methylation of a tumor suppressor gene L3MBTL1. Breast Cancer Res. Treat. 2012, 133, 127–135. [Google Scholar] [CrossRef]
- Garbiec, E.; Cielecka-Piontek, J.; Kowalówka, M.; Hołubiec, M.; Zalewski, P. Genistein—Opportunities Related to an Interesting Molecule of Natural Origin. Molecules 2022, 27, 815. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Arora, I.; Chen, M.; Wu, H.; Crowley, M.R.; Tollefsbol, T.O.; Li, Y. Therapeutic Effects of Dietary Soybean Genistein on Triple-Negative Breast Cancer via Regulation of Epigenetic Mechanisms. Nutrients 2021, 13, 3944. [Google Scholar] [CrossRef]
- Paul, B.; Li, Y.; Tollefsbol, T.O. The Effects of Combinatorial Genistein and Sulforaphane in Breast Tumor Inhibition: Role in Epigenetic Regulation. Int. J. Mol. Sci. 2018, 19, 1754. [Google Scholar] [CrossRef]
- Chang, H.; Mi, M.; Ling, W.; Zhu, J.; Zhang, Q.; Wei, N.; Zhou, Y.; Tang, Y.; Yuan, J. Structurally related cytotoxic effects of flavonoids on human cancer cells in vitro. Arch. Pharmacal Res. 2008, 31, 1137–1144. [Google Scholar] [CrossRef]
- Androutsopoulos, V.; Papakyriakou, A.; Vourloumis, D.; Spandidos, D.A. Comparative CYP1A1 and CYP1B1 substrate and inhibitor profile of dietary flavonoids. Bioorg. Med. Chem. 2011, 19, 2842–2849. [Google Scholar] [CrossRef]
- Pal-Bhadra, M.; Ramaiah, M.J.; Reddy, T.L.; Krishnan, A.; Pushpavalli, S.; Babu, K.S.; Tiwari, A.K.; Rao, J.M.; Yadav, J.S.; Bhadra, U. Plant HDAC inhibitor chrysin arrest cell growth and induce p21 WAF1 by altering chromatin of STAT response element in A375 cells. BMC Cancer 2012, 12, 180. [Google Scholar] [CrossRef]
- Sedlak, L.; Wojnar, W.; Zych, M.; Wyględowska-Promieńska, D.; Mrukwa-Kominek, E.; Kaczmarczyk-Sedlak, I. Effect of Resveratrol, a Dietary-Derived Polyphenol, on the Oxidative Stress and Polyol Pathway in the Lens of Rats with Streptozotocin-Induced Diabetes. Nutrients 2018, 10, 1423. [Google Scholar] [CrossRef]
- Galiniak, S.; Aebisher, D.; Bartusik-Aebisher, D. Health benefits of resveratrol administration. Acta Biochim. Pol. 2019, 66, 13–21. [Google Scholar] [CrossRef]
- Venturelli, S.; Berger, A.; Böcker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Schleicher, S.; Mayer, M.; Weiss, T.S.; et al. Resveratrol as a pan-HDAC inhibitor alters the acetylation status of histone [corrected] proteins in human-derived hepatoblastoma cells. PLoS ONE 2013, 8, e73097. [Google Scholar] [CrossRef]
- Izquierdo-Torres, E.; Hernández-Oliveras, A.; Meneses-Morales, I.; Rodríguez, G.; Fuentes-García, G.; Zarain-Herzberg, Á. Resveratrol up-regulates ATP2A3 gene expression in breast cancer cell lines through epigenetic mechanisms. Int. J. Biochem. Cell Biol. 2019, 113, 37–47. [Google Scholar] [CrossRef]
- Zheng, Y.; Liu, Z.; Yang, X.; Liu, L.; Ahn, K.S. An updated review on the potential antineoplastic actions of oleuropein. Phytother. Res. 2022, 36, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Bayat, S.; Derakhshan, S.M.; Derakhshan, N.M.; Khaniani, M.S.; Alivand, M.R. Downregulation of HDAC2 and HDAC3 via oleuropein as a potent prevention and therapeutic agent in MCF-7 breast cancer cells. J. Cell. Biochem. 2019, 120, 9172–9180. [Google Scholar] [CrossRef] [PubMed]
- Jabczyk, M.; Nowak, J.; Hudzik, B.; Zubelewicz-Szkodzińska, B. Curcumin in Metabolic Health and Disease. Nutrients 2021, 13, 4440. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-Q.; Yu, K.; Yan, Q.-X.; Xing, C.-Y.; Chen, Y.; Yan, Z.; Shi, Y.-F.; Zhao, K.-W.; Gao, S.-M. Pure curcumin increases the expression of SOCS1 and SOCS3 in myeloproliferative neoplasms through suppressing class Ι histone deacetylases. Carcinogenesis 2013, 34, 1442–1449. [Google Scholar] [CrossRef]
- Han, R.; Yang, H.; Li, Y.; Ling, C.; Lu, L. Valeric acid acts as a novel HDAC3 inhibitor against prostate cancer. Med. Oncol. 2022, 39, 213. [Google Scholar] [CrossRef]
- Shi, F.; Li, Y.; Han, R.; Fu, A.; Wang, R.; Nusbaum, O.; Qin, Q.; Chen, X.; Hou, L.; Zhu, Y. Valerian and valeric acid inhibit growth of breast cancer cells possibly by mediating epigenetic modifications. Sci. Rep. 2021, 11, 2519. [Google Scholar] [CrossRef]
- Wu, Y.; Duan, Z.; Qu, L.; Zhang, Y.; Zhu, C.; Fan, D. Gastroprotective effects of ginsenoside Rh4 against ethanol-induced gastric mucosal injury by inhibiting the MAPK/NF-κB signaling pathway. Food Funct. 2023, 14, 5167–5181. [Google Scholar] [CrossRef]
- Dong, F.; Qu, L.; Duan, Z.; He, Y.; Ma, X.; Fan, D. Ginsenoside Rh4 inhibits breast cancer growth through targeting histone deacetylase 2 to regulate immune microenvironment and apoptosis. Bioorganic Chem. 2023, 135, 106537. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; He, T.; Becker, S.; Zhang, G.; Li, D.; Ma, X. Butyrate: A Double-Edged Sword for Health? Adv. Nutr. 2018, 9, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr. Protein Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Canani, R.B.; Di Costanzo, M.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef] [PubMed]
- Andrade, F.; Nagamine, M.; De Conti, A.; Chaible, L.; Fontelles, C.; Junior, A.J.; Vannucchi, H.; Dagli, M.; Bassoli, B.; Moreno, F.; et al. Efficacy of the dietary histone deacetylase inhibitor butyrate alone or in combination with vitamin A against proliferation of MCF-7 human breast cancer cells. Braz. J. Med. Biol. Res. 2012, 45, 841–850. [Google Scholar] [CrossRef]
- Chopin, V.; Slomianny, C.; Hondermarck, H.; Le Bourhis, X. Synergistic induction of apoptosis in breast cancer cells by cotreatment with butyrate and TNF-alpha, TRAIL, or anti-Fas agonist antibody involves enhancement of death receptors’ signaling and requires P21(waf1). Exp. Cell Res. 2004, 298, 560–573. [Google Scholar] [CrossRef]
- Thakur, V.S.; Gupta, K.; Gupta, S. Green tea polyphenols causes cell cycle arrest and apoptosis in prostate cancer cells by suppressing class I histone deacetylases. Carcinogenesis 2012, 33, 377–384. [Google Scholar] [CrossRef]
- Wu, K.; Wei, Y.; Yu, Y.; Shan, M.; Tang, Y.; Sun, Y. Green tea polyphenols inhibit malignant melanoma progression via regulating circ_MITF/miR-30e-3p/HDAC2 axis. Biotechnol. Appl. Biochem. 2022, 69, 808–821. [Google Scholar] [CrossRef]
- Jang, Y.-G.; Hwang, K.-A.; Choi, K.-C. Rosmarinic Acid, a Component of Rosemary Tea, Induced the Cell Cycle Arrest and Apoptosis through Modulation of HDAC2 Expression in Prostate Cancer Cell Lines. Nutrients 2018, 10, 1784. [Google Scholar] [CrossRef]
- Kim, H.; Ramirez, C.N.; Su, Z.-Y.; Kong, A.-N.T. Epigenetic modifications of triterpenoid ursolic acid in activating Nrf2 and blocking cellular transformation of mouse epidermal cells. J. Nutr. Biochem. 2016, 33, 54–62. [Google Scholar] [CrossRef]
- Çakır, I.; Pan, P.L.; Hadley, C.K.; El-Gamal, A.; Fadel, A.; Elsayegh, D.; Mohamed, O.; Rizk, N.M. Is a corresponding author Masoud Ghamari-Langroudi. Sulforaphane reduces obesity by reversing leptin resistance. Elife 2022, 11, e67368. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Bai, Q.; Zou, L.-Y.; Zhang, Q.-Y.; Zhou, Y.; Chang, H.; Yi, L.; Zhu, J.-D.; Mi, M.-T. Genistein inhibits DNA methylation and increases expression of tumor suppressor genes in human breast cancer cells. Genes Chromosom. Cancer 2014, 53, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Tortorella, S.M.; Royce, S.G.; Licciardi, P.V.; Karagiannis, T.C. Dietary Sulforaphane in Cancer Chemoprevention: The Role of Epigenetic Regulation and HDAC Inhibition. Antioxid. Redox Signal. 2015, 22, 1382–1424. [Google Scholar] [CrossRef]
- Middleton, E., Jr.; Kandaswami, C.; Theoharides, T.C. The effects of plant flavonoids on mammalian cells: Implications for inflammation, heart disease, and cancer. Pharmacol. Rev. 2000, 52, 673–751. [Google Scholar] [PubMed]
- Davie, J.R. Inhibition of Histone Deacetylase Activity by Butyrate. J. Nutr. 2003, 133 (Suppl. 7), 2485S–2493S. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, Z.; Ma, Y.; Liu, Y.; Lin, C.-C.; Li, S.; Zhan, J.; Ho, C.-T. Immunomodulatory Effects of Green Tea Polyphenols. Molecules 2021, 26, 3755. [Google Scholar] [CrossRef]
- Han, R.; Yang, H.; Ling, C.; Lu, L. Neurospora crassa is a potential source of anti-cancer agents against breast cancer. Breast Cancer 2022, 29, 1032–1041. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Dima, C.; Assadpour, E.; Nechifor, A.; Dima, S.; Li, Y.; Jafari, S.M. Oral bioavailability of bioactive compounds; modulating factors, in vitro analysis methods, and enhancing strategies. Crit. Rev. Food Sci. Nutr. 2023, 1–39. [Google Scholar] [CrossRef]
- Rosenfeld, R.M.; Juszczak, H.M.; Wong, M.A. Scoping review of the association of plant-based diet quality with health out-comes. Front. Nutr. 2023, 10, 1211535. [Google Scholar] [CrossRef]
- Weaver, C.M.; Alekel, D.L.; Ward, W.E.; Ronis, M.J. Flavonoid Intake and Bone Health. J. Nutr. Gerontol. Geriatr. 2012, 31, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Shao, H.; Gao, L.; Li, H.; Sheng, H.; Zhu, L. Nano-drug co-delivery system of natural active ingredients and chemotherapy drugs for cancer treatment: A review. Drug Deliv. 2022, 29, 2130–2161. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, Y.; Mukai, S.; Yamada, S.; Matsuoka, M.; Tarumi, E.; Hashimoto, T.; Tamura, C.; Imaizumi, A.; Nishihira, J.; Nakamura, T. Short-term effects of highly-bioavailable curcumin for treating knee osteoarthritis: A randomized, double-blind, placebo-controlled prospective study. J. Orthop. Sci. 2014, 19, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Kanai, M.; Imaizumi, A.; Otsuka, Y.; Sasaki, H.; Hashiguchi, M.; Tsujiko, K.; Matsumoto, S.; Ishiguro, H.; Chiba, T. Dose-escalation and pharmacokinetic study of nanoparticle curcumin, a potential anticancer agent with improved bioavailability, in healthy human volunteers. Cancer Chemother. Pharmacol. 2012, 69, 65–70. [Google Scholar] [CrossRef]
- Sasaki, H.; Sunagawa, Y.; Takahashi, K.; Imaizumi, A.; Fukuda, H.; Hashimoto, T.; Wada, H.; Katanasaka, Y.; Kakeya, H.; Fujita, M.; et al. Innovative Preparation of Curcumin for Improved Oral Bioavailability. Biol. Pharm. Bull. 2011, 34, 660–665. [Google Scholar] [CrossRef]
- Gao, C.; Tang, F.; Gong, G.; Zhang, J.; Hoi, M.P.M.; Lee, S.M.Y.; Wang, R. pH-Responsive prodrug nanoparticles based on a sodium alginate derivative for selective co-release of doxorubicin and curcumin into tumor cells. Nanoscale 2017, 9, 12533–12542. [Google Scholar] [CrossRef]
- Zhang, H.; Xue, Q.; Zhou, Z.; He, N.; Li, S.; Zhao, C. Co-delivery of doxorubicin and hydroxychloroquine via chitosan/alginate nanoparticles for blocking autophagy and enhancing chemotherapy in breast cancer therapy. Front. Pharmacol. 2023, 14, 1176232. [Google Scholar] [CrossRef]
- Dong, X.; Sun, Y.; Li, Y.; Ma, X.; Zhang, S.; Yuan, Y.; Kohn, J.; Liu, C.; Qian, J. Synergistic Combination of Bioactive Hydroxyapatite Nanoparticles and the Chemotherapeutic Doxorubicin to Over-come Tumor Multidrug Resistance. Small 2021, 17, e2007672. [Google Scholar] [CrossRef]
- Li, Z.; Wang, L.; Lin, X.; Shen, L.; Feng, Y. Drug delivery for bioactive polysaccharides to improve their drug-like properties and curative efficacy. Drug Deliv. 2017, 24 (Suppl. 1), 70–80. [Google Scholar] [CrossRef]
- Prasad, S.; Gupta, S.C.; Tyagi, A.K.; Aggarwal, B.B. Curcumin, a component of golden spice: From bedside to bench and back. Biotechnol. Adv. 2014, 32, 1053–1064. [Google Scholar] [CrossRef]
- Passos, C.L.A.; Polinati, R.M.; Ferreira, C.; dos Santos, N.A.N.; Lima, D.G.V.; da Silva, J.L.; Fialho, E. Curcumin and melphalan cotreatment induces cell cycle arrest and apoptosis in MDA-MB-231 breast cancer cells. Sci. Rep. 2023, 13, 13446. [Google Scholar] [CrossRef]
- Zhu, L.; Chen, L. Progress in research on paclitaxel and tumor immunotherapy. Cell. Mol. Biol. Lett. 2019, 24, 40. [Google Scholar] [CrossRef] [PubMed]
- Nootim, P.; Kapol, N.; Bunchuailua, W.; Poompruek, P.; Tungsukruthai, P. Current state of cancer patient care incorporating Thai traditional medicine in Thailand: A qualitative study. J. Integr. Med. 2020, 18, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Yazdi, N.; Salehi, A.; Vojoud, M.; Sharifi, M.H.; Hoseinkhani, A. Use of complementary and alternative medicine in pregnant women: A cross-sectional survey in the south of Iran. J. Integr. Med. 2019, 17, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, W.; Meng, Y.; Guo, G.; Zhao, C.; Zhou, X. Efficacy and safety of traditional Chinese medicine in the treatment of osteonecrosis of the femoral head. J. Orthop. Surg. Res. 2023, 18, 600. [Google Scholar] [CrossRef]
- Bu, Z.-J.; Liu, Y.-N.; Shahjalal, M.; Zheng, Y.-Y.; Liu, C.-J.; Ye, M.-M.; Xu, J.-Y.; Peng, X.-Y.; Wang, X.-H.; Chen, X.; et al. Comparative effectiveness and safety of Chinese medicine belly button application for childhood diarrhea: A Bayesian network meta-analysis of randomized controlled trials. Front. Pediatr. 2023, 11, 1180694. [Google Scholar] [CrossRef]
- Bushehri, R.H.; Navabi, P.; Saeedifar, A.M.; Keshavarzian, N.; Rouzbahani, N.H.; Mosayebi, G.; Ghazavi, A.; Ghorban, K.; Ganji, A. Integration of phytotherapy and chemotherapy: Recent advances in anticancer molecular pathways. Iran J. Basic Med. Sci. 2023, 26, 987–1000. [Google Scholar] [CrossRef]
- Zhang, X.; Li, H.; Lv, X.; Hu, L.; Li, W.; Zi, M.; He, Y. Impact of Diets on Response to Immune Checkpoint Inhibitors (ICIs) Therapy against Tumors. Life 2022, 12, 409. [Google Scholar] [CrossRef]
- Kromhout, D.; Yasuda, S.; Geleijnse, J.M.; Shimokawa, H. Fish oil and omega-3 fatty acids in cardiovascular disease: Do they really work? Eur. Heart J. 2012, 33, 436–443. [Google Scholar] [CrossRef]
- Björkström, N.K.; Strunz, B.; Ljunggren, H.-G. Natural killer cells in antiviral immunity. Nat. Rev. Immunol. 2022, 22, 112–123. [Google Scholar] [CrossRef]
- Thies, F.; Nebe-von-Caron, G.; Powell, J.R.; Yaqoob, P.; Newsholme, E.A.; Calder, P.C. Dietary supplementation with eicosapentaenoic acid, but not with other long-chain n-3 or n-6 polyunsaturated fatty acids, decreases natural killer cell activity in healthy subjects aged >55 y. Am. J. Clin. Nutr. 2001, 73, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, S.P.; Burda, B.U.; Senger, C.A.; Lin, J.S.; Whitlock, E.P. Vitamin and Mineral Supplements in the Primary Prevention of Cardiovascular Disease and Cancer: An Updated Systematic Evidence Review for the U.S. Preventive Services Task Force. Ann. Intern. Med. 2013, 159, 824–834. [Google Scholar] [CrossRef]
- Beck, K.L.; von Hurst, P.R.; O’Brien, W.J.; Badenhorst, C.E. Micronutrients and athletic performance: A review. Food Chem. Toxicol. 2021, 158, 112618. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.; Sartory, N.A.; Zahn, N.; Radeke, H.H.; Stein, J.M. Immune modulatory treatment of trinitrobenzene sulfonic acid colitis with calcitriol is associated with a change of a T helper (Th) 1/Th17 to a Th2 and regulatory T cell profile. J. Pharmacol. Exp. Ther. 2008, 324, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Larkin, J.; Chiarion-Sileni, V.; Gonzalez, R.; Grob, J.-J.; Cowey, C.L.; Lao, C.D.; Schadendorf, D.; Dummer, R.; Smylie, M.; Rutkowski, P.; et al. Combined Nivolumab and Ipilimumab or Monotherapy in Untreated Melanoma. N. Engl. J. Med. 2015, 373, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Tobin, R.P.; Jordan, K.R.; Robinson, W.A.; Davis, D.; Borges, V.F.; Gonzalez, R.; Lewis, K.D.; McCarter, M.D. Targeting myeloid-derived suppressor cells using all-trans retinoic acid in melanoma patients treated with Ipili-mumab. Int. Immunopharmacol. 2018, 63, 282–291. [Google Scholar] [CrossRef]
- Chen, L.; Diao, L.; Yang, Y.; Yi, X.; Rodriguez, B.L.; Li, Y.; Villalobos, P.A.; Cascone, T.; Liu, X.; Tan, L.; et al. CD38-Mediated Immunosuppression as a Mechanism of Tumor Cell Escape from PD-1/PD-L1 Blockade. Cancer Discov. 2018, 8, 1156–1175. [Google Scholar] [CrossRef]
- Yuan, J.; Li, J.; Shang, M.; Fu, Y.; Wang, T. Identification of vitamin B6 as a PD-L1 suppressor and an adjuvant for cancer immunotherapy. Biochem. Biophys. Res. Commun. 2021, 561, 187–194. [Google Scholar] [CrossRef]
- Kim, S.-K.; Guevarra, R.B.; Kim, Y.-T.; Kwon, J.; Kim, H.; Cho, J.H.; Kim, H.B.; Lee, J.-H. Role of Probiotics in Human Gut Microbiome-Associated Diseases. J. Microbiol. Biotechnol. 2019, 29, 1335–1340. [Google Scholar] [CrossRef]
- Bedada, T.L.; Feto, T.K.; Awoke, K.S.; Garedew, A.D.; Yifat, F.T.; Birri, D.J. Probiotics for cancer alternative prevention and treatment. BioMedicine 2020, 129, 110409. [Google Scholar] [CrossRef]
- Suez, J.; Zmora, N.; Zilberman-Schapira, G.; Mor, U.; Dori-Bachash, M.; Bashiardes, S.; Zur, M.; Regev-Lehavi, D.; Brik, R.B.-Z.; Federici, S.; et al. Post-Antibiotic Gut Mucosal Microbiome Reconstitution Is Impaired by Probiotics and Improved by Autologous FMT. Cell 2018, 174, 1406–1423.e16. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.N.; Gopalakrishnan, V.; McQuade, J.; Andrews, M.C.; Helmink, B.; Khan, M.W.; Sirmans, E.; Haydu, L.; Cogdill, A.; Burton, E.; et al. Abstract 2838: The gut microbiome (GM) and immunotherapy response are influenced by host lifestyle factors. Cancer Res 2019, 79 (Suppl. 13), 2838. [Google Scholar] [CrossRef]
- Kuang, R.; Binion, D.G. Should high-fiber diets be recommended for patients with inflammatory bowel disease? Curr. Opin. Gas-Troenterol. 2022, 38, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef] [PubMed]
- Kocher, F.; Amann, A.; Zimmer, K.; Geisler, S.; Fuchs, D.; Pichler, R.; Wolf, D.; Kurz, K.; Seeber, A.; Pircher, A. High indoleamine-2,3-dioxygenase 1 (IDO) activity is linked to primary resistance to immunotherapy in non-small cell lung cancer (NSCLC). Transl. Lung Cancer Res. 2021, 10, 304–313. [Google Scholar] [CrossRef] [PubMed]
- Weber, D.D.; Aminzadeh-Gohari, S.; Tulipan, J.; Catalano, L.; Feichtinger, R.G.; Kofler, B. Ketogenic diet in the treatment of cancer—Where do we stand? Mol. Metab. 2020, 33, 102–121. [Google Scholar] [CrossRef]
- Lussier, D.M.; Woolf, E.C.; Johnson, J.L.; Brooks, K.S.; Blattman, J.N.; Scheck, A.C. Enhanced immunity in a mouse model of malignant glioma is mediated by a therapeutic ketogenic diet. BMC Cancer 2016, 16, 310. [Google Scholar] [CrossRef]
- Rom-Jurek, E.-M.; Kirchhammer, N.; Ugocsai, P.; Ortmann, O.; Wege, A.K.; Brockhoff, G. Regulation of Programmed Death Ligand 1 (PD-L1) Expression in Breast Cancer Cell Lines In Vitro and in Immunodeficient and Humanized Tumor Mice. Int. J. Mol. Sci. 2018, 19, 563. [Google Scholar] [CrossRef]
- Orillion, A.R.; Damayanti, N.P.; Shen, L.; Adelaiye-Ogala, R.; Affronti, H.C.; Elbanna, M.; Chintala, S.; Ciesielski, M.J.; Fontana, L.; Kao, C.; et al. Dietary Protein Restriction Reprograms Tumor-Associated Macrophages and Enhances Immunotherapy. Clin. Cancer Res. 2018, 24, 6383–6395. [Google Scholar] [CrossRef]
- Nabe, S.; Yamada, T.; Suzuki, J.; Toriyama, K.; Yasuoka, T.; Kuwahara, M.; Shiraishi, A.; Takenaka, K.; Yasukawa, M.; Yamashita, M. Reinforce the antitumor activity of CD8+ T cells via glutamine restriction. Cancer Sci. 2018, 109, 3737–3750. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Measurement | Sample Size | Patient Selection Criteria | Outcomes | Year | Refs |
|---|---|---|---|---|---|
| IHC | 226 | sporadic breast cancer patients who underwent surgery; All patients did not undergo radiation therapy and chemotherapy before surgery | High expression of HDAC2 was associated with: 1. advanced clinical stages (p = 0.016); 2. lymphatic metastasis (p = 0.02); 3. high histological grade (p = 0.001); 4. shorter OS of BC patients (p = 0.035); 5. shorter OS in multidrug resistance protein-positive patients (p = 0.034); 6. shorter survival in patients who received chemotherapy containing anthracyclines (OS, p = 0.041; disease-free survival, p = 0.084). | 2016 | [15] |
| IHC | 118 | Tumor size < 20 mm; Stage I, II and III; No preoperative anticancer therapy; Deceased from BC | High expression of HDAC2 was associated with: 1. BC in stage III (p < 0.001); 2. BC with histological grade 3 (p = 0.013). | 2022 | [13] |
| IHC | 300 | Invasive ductal carcinoma patients who underwent curative surgery | High expression of HDAC2 was correlated with improved OS in ER-negative BC patients (p = 0.048). | 2014 | [23] |
| IHC | 212 | patients with primary invasive breast cancer | High expression of HDAC2 was associated with: 1.overexpression of HER2 (p = 0.005); 2. lymphatic metastasis (p = 0.04). | 2013 | [24] |
| Dietary Component | Food Source | Potential Benefits in Breast Cancer | Diseases | Refs |
|---|---|---|---|---|
| Genistein (GE) | Soybean products | Anti-cancer effects on breast cancer, modulation of Dnmt3b, Tet3 and HDAC | Breast cancer, cervical cancer, | [85] |
| Sulforaphane (SFN) | Broccoli sprouts, kale | Cytotoxic effects in breast, colon, and prostate cancer cells, inhibition of HDAC-2 and HDAC-8 | Breast cancer, colon cancer, prostate cancer, neurodegenerative diseases, bladder carcinoma, | [86] |
| Chrysin | Fruits, vegetables, olive oil and red wine | Cytotoxic effects in breast, colon, and prostate cancer cells, inhibition of HDAC-2 and HDAC-8 | Breast cancer, melanoma, fibrosarcoma, leukemia | [87,88,89] |
| Resveratrol (RSV) | Grapes, apples, blueberries, mulberries, peanuts, pistachios, plums, and red wine | Induced ATP2A3 upregulation correlates with reduced HDAC activity and reduced nuclear HDAC2 expression and occupancy on ATP2A3 promoter | Breast cancer, glioma | [90,91,92,93] |
| Oleuropein (OLE) | Virgin olive oil | Antineoplastic properties, modulation of HDAC2 and HDAC3 in breast cancer cells | Breast cancer, alzheimer, | [94,95] |
| Curcumin | Turmeric | Inhibited both HDAC activity and the expression of HDACs 1 and 2 in a concentration-dependent manner in cancer cells | Breast Cancer, lung cancer, hematological cancers, inflammation, arthritis, metabolic syndrome, | [96,97] |
| Valeric acid | Valerian herb | Anti-cancer effects on liver and breast cancer, modulation of HDAC2 and HDAC3 | Breast cancer, prostate cancer, liver cancer | [98,99] |
| Ginsenoside Rh4 | Ginseng herb | Inhibition of PD-L1 expression by regulating HDAC2-mediated JAK/STAT pathway in breast cancer cells | Breast cancer, lung adenocarcinoma, colorectal cancer, gastric cancer | [100,101] |
| Butyrate | Dietary fiber, resistant starch, undigested carbohydrates | Anticancer activity, inhibition of breast cancer cell proliferation, induction of apoptosis | Multiple cancer, cardiovascular disease, and type 2 diabetes | [102,103,104,105,106] |
| Green tea polyphenols | Green tea | Suppressed cancer progression by regulating circ_MITF/miR-30e-3p/HDAC2 axis | breast cancer, malignant melanoma, prostate cancer, | [107,108] |
| Rosmarinic acid (RA) | Rosemary tea | Potential pan-HDAC inhibitor, inhibition of nuclear HDAC2 protein levels in breast cancer cells | Breast cancer, prostate cancer | [109] |
| Ursolic acid (UA) | Blueberries, cranberries and apple peels | Reduced the expression of epigenetic modifying enzymes, including the DNMT1 and DNMT3a and the histone deacetylases (HDACs) HDAC1, HDAC2, HDAC3, HDAC6 and HDAC7 activity | Skin cancer, breast cancer, colorectal cancer | [110] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Lu, L.; Ling, C.; Zhang, P.; Han, R. Potential of Dietary HDAC2i in Breast Cancer Patients Receiving PD-1/PD-L1 Inhibitors. Nutrients 2023, 15, 3984. https://doi.org/10.3390/nu15183984
Wang Y, Lu L, Ling C, Zhang P, Han R. Potential of Dietary HDAC2i in Breast Cancer Patients Receiving PD-1/PD-L1 Inhibitors. Nutrients. 2023; 15(18):3984. https://doi.org/10.3390/nu15183984
Chicago/Turabian StyleWang, Yuqian, Lingeng Lu, Changquan Ling, Ping Zhang, and Rui Han. 2023. "Potential of Dietary HDAC2i in Breast Cancer Patients Receiving PD-1/PD-L1 Inhibitors" Nutrients 15, no. 18: 3984. https://doi.org/10.3390/nu15183984