
Effects of Low Vitamin C Intake on Fertility Parameters and Pregnancy Outcomes in Guinea Pigs
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Animal Model
2.3. Breeding
2.4. Delivery and Postnatal Care of Offspring
2.5. Euthanasia and Tissue Collection
2.6. High-Performance Liquid Chromatography (HPLC)
2.7. Cortisol Enzyme Linked Immunosorbent Assay (ELISA)
2.8. Statistical Analyses
3. Results
3.1. Adult Physical Characteristics
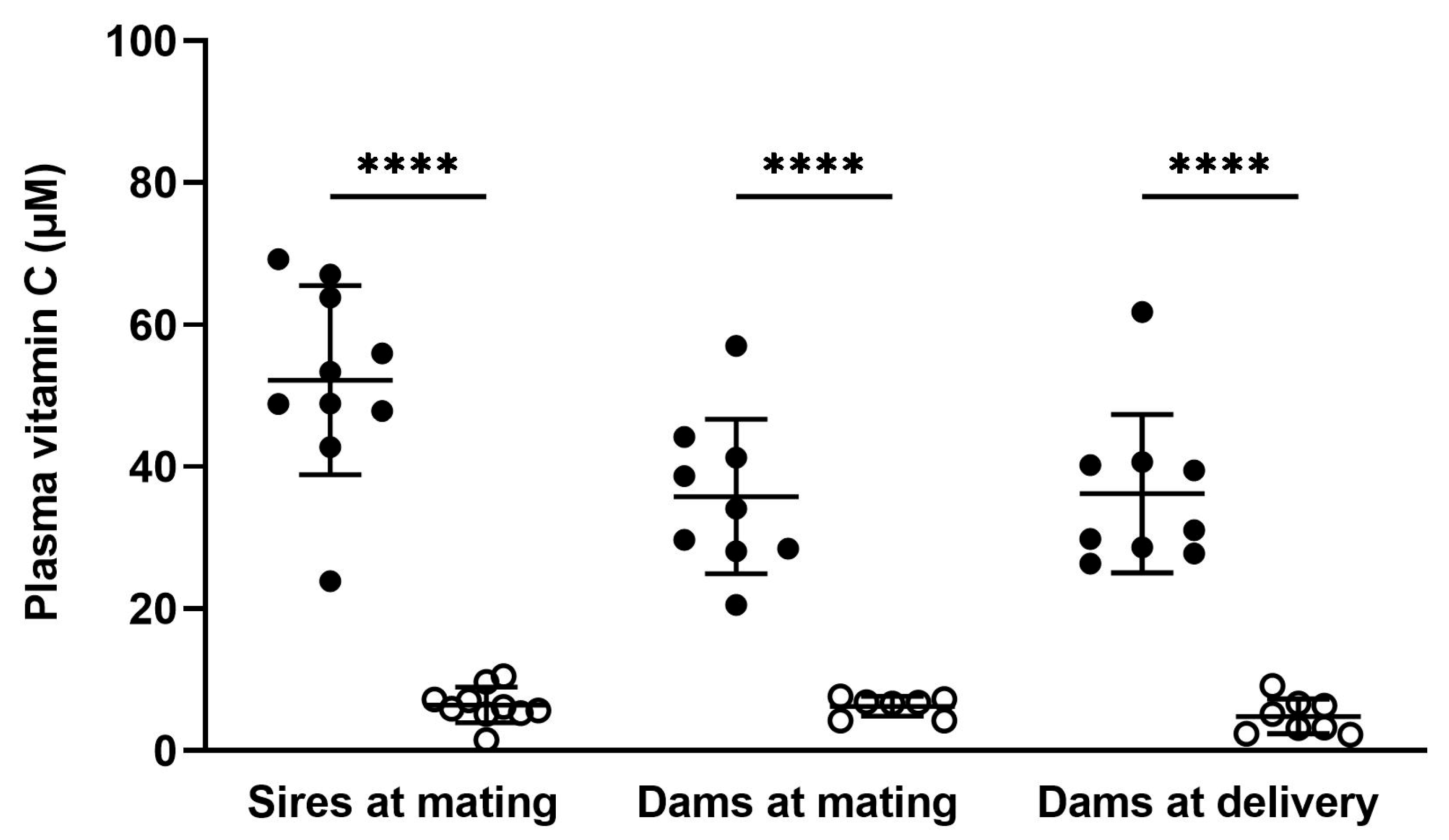
3.2. Vitamin C Concentrations in Adults
3.2.1. Plasma
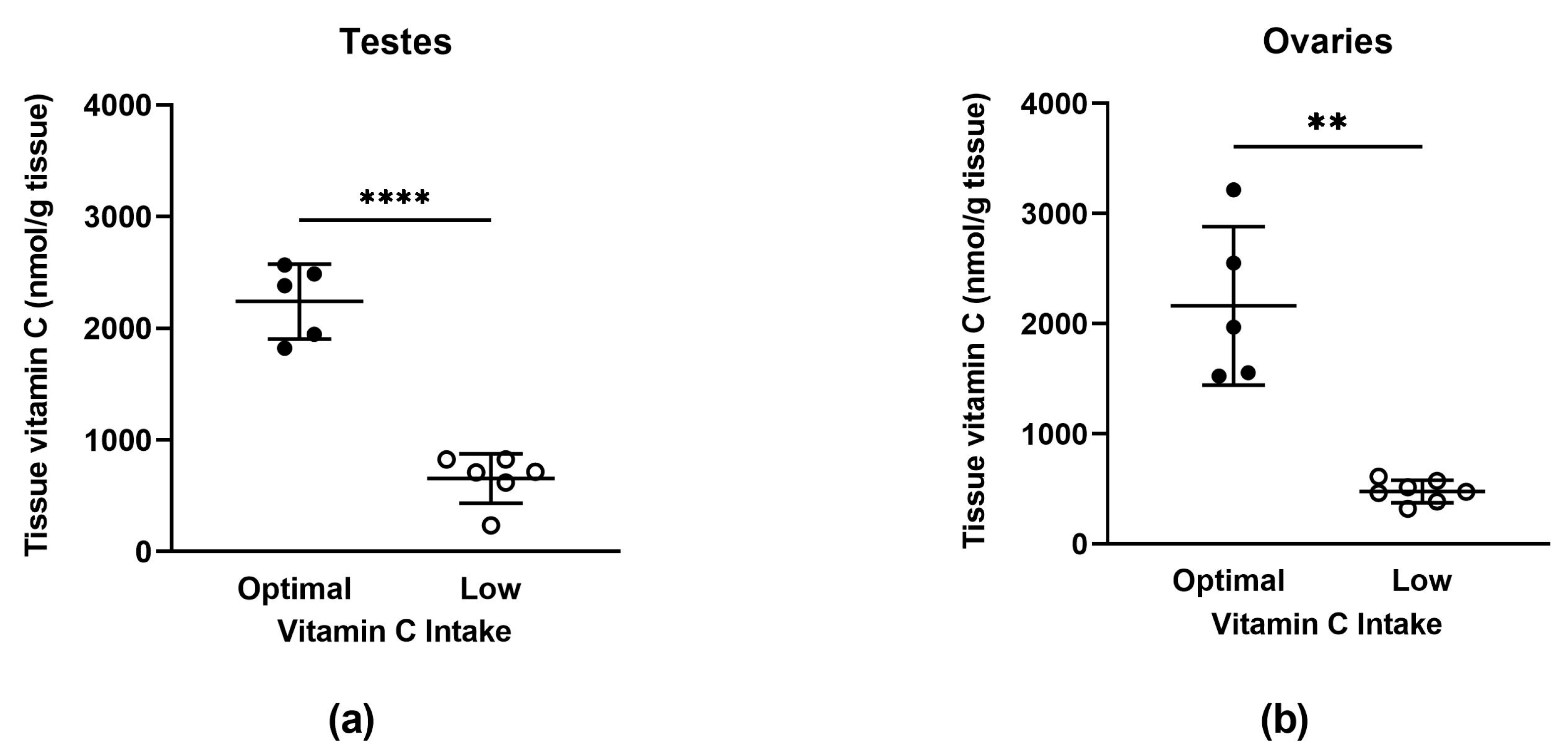
3.2.2. Tissue
3.3. Fertility Parameters
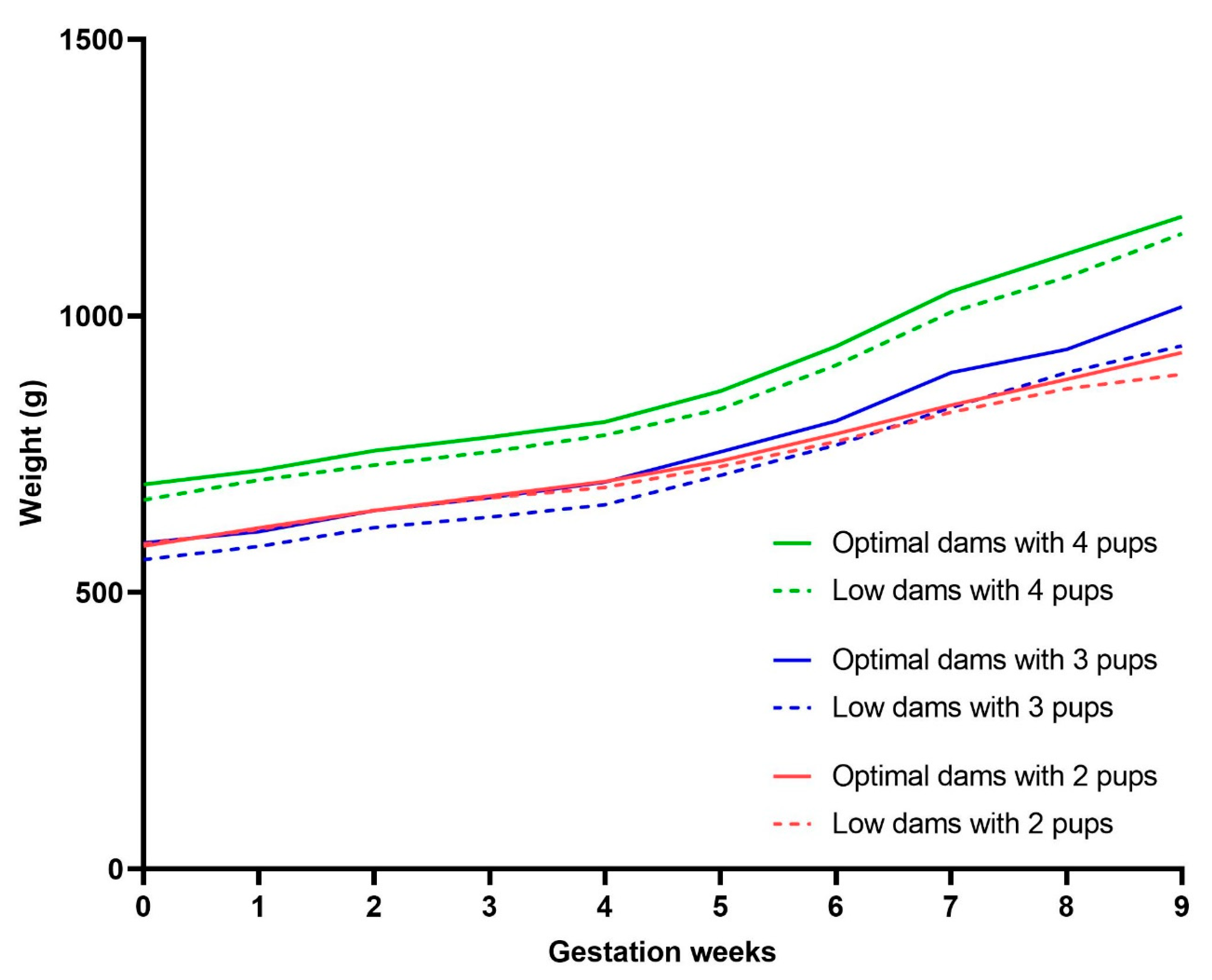
3.4. Pregnancy Weight Gain
3.5. Pregnancy Outcomes
3.6. Offspring Physical Characteristics
3.7. Vitamin C Concentrations in Offspring
Plasma
3.8. Salivary Cortisol Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drouin, G.; Godin, J.-R.; Page, B. The genetics of vitamin C loss in vertebrates. Curr. Genom. 2011, 12, 371–378. [Google Scholar] [CrossRef]
- Paeschke, K.D. Ovulation preliminaries and ovulation. I. Generative function of the ovary and ascorbic acid metabolism during the ovarian cycle. Fortschr. Geburtshilfe Gynakol. 1970, 43, 1–58. [Google Scholar] [PubMed]
- Das, P.C.; Das, K.P.; Bagchi, K.; Dey, C.D. Evaluation of tissue ascorbic acid status in different hormonal states of female rat. Life Sci. 1993, 52, 1493–1498. [Google Scholar] [CrossRef]
- Ranjan, R.; Ranjan, A.; Dhaliwal, G.; Patra, R. l-Ascorbic acid (vitamin C) supplementation to optimize health and reproduction in cattle. Vet. Q. 2012, 32, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Michos, C.; Kiortsis, D.N.; Evangelou, A.; Karkabounas, S. Antioxidant protection during the menstrual cycle: The effects of estradiol on ascorbic-dehydroascorbic acid plasma levels and total antioxidant plasma status in eumenorrhoic women during the menstrual cycle. Acta Obstet. Gynecol. Scand. 2006, 85, 960–965. [Google Scholar] [CrossRef] [PubMed]
- Loh, H.; Wilson, C. Relationship of human ascorbic-acid metabolism to ovulation. Lancet 1971, 1, 110–112. [Google Scholar] [CrossRef] [PubMed]
- Pillay, A.P. Vitamin C and Ovulation. Ind. Med. Gaz. 1940, 75, 91–93. [Google Scholar] [PubMed]
- Olaniyan, O.T.; Femi, A.; Iliya, G.; Ayobami, D.; Godam, E.; Olugbenga, E.; Bamidele, O.; Mali, P.C. Vitamin C suppresses ovarian pathophysiology in experimental polycystic ovarian syndrome. Pathophysiology 2019, 26, 331–341. [Google Scholar] [CrossRef]
- Luck, M.R.; Jeyaseelan, I.; Scholes, R.A. Ascorbic acid and fertility. Biol. Reprod. 1995, 52, 262–266. [Google Scholar] [CrossRef]
- Iervolino, M.; Lepore, E.; Forte, G.; Laganà, A.; Buzzaccarini, G.; Unfer, V. Natural Molecules in the Management of Polycystic Ovary Syndrome (PCOS): An Analytical Review. Nutrients 2021, 13, 1677. [Google Scholar] [CrossRef]
- Mesen, T.B.; Young, S.L. Progesterone and the luteal phase: A requisite to reproduction. Obstet. Gynecol. Clin. N. Am. 2015, 42, 135–151. [Google Scholar] [CrossRef] [PubMed]
- Henmi, H.; Endo, T.; Kitajima, Y.; Manase, K.; Hata, H.; Kudo, R. Effects of ascorbic acid supplementation on serum progesterone levels in patients with a luteal phase defect. Fertil. Steril. 2003, 80, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Mumford, S.L.; Browne, R.W.; Schliep, K.C.; Schmelzer, J.; Plowden, T.C.; Michels, K.A.; Sjaarda, L.A.; Zarek, S.M.; Perkins, N.J.; Messer, L.C.; et al. Serum Antioxidants Are Associated with Serum Reproductive Hormones and Ovulation among Healthy Women. J. Nutr. 2016, 146, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Kramer, M.M.; Harman, M.T.; Brill, A.K.; Takami, M.; Preston, S.L.; Toyloy, V.A.; Behrman, H.R. Disturbances of reproduction and ovarian changes in the guinea-pig in relation to vitamin C deficiency. Am. J. Physiol. Leg. Content 1933, 106, 611–622. [Google Scholar] [CrossRef]
- Morris, G.E. Hypermenorrhea due to scurvy. Postgrad. Med. 1953, 14, 443–445. [Google Scholar] [CrossRef]
- Jacob, R.A.; Planalto, F.S.; Agee, R.E. Cellular ascorbate depletion in healthy men. J. Nutr. 1992, 122, 1111–1118. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, H.; Zhu, S.; Wang, H.; Sun, S. Effects of vitamin E and vitamin C on male infertility: A meta-analysis. Int. Urol. Nephrol. 2022, 54, 1793–1805. [Google Scholar] [CrossRef]
- Asadi, N.; Bahmani, M.; Kheradmand, A.; Rafieian-Kopaei, M. The Impact of Oxidative Stress on Testicular Function and the Role of Antioxidants in Improving it: A Review. J. Clin. Diagn. Res. 2017, 11, IE01–IE05. [Google Scholar] [CrossRef]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell. Mol. Life Sci. 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Akmal, M.; Qadri, J.; Al-Waili, N.S.; Thangal, S.; Haq, A.; Saloom, K.Y.; Deruelle, F.; Baron, B.; Vani, K.; Kurakula, M.; et al. Improvement in human semen quality after oral supplementation of vitamin C. J. Med. Food 2006, 9, 440–442. [Google Scholar] [CrossRef]
- De Oliveira, A.M.; Rondo, P.H.; Barros, S.B. Concentrations of ascorbic acid in the plasma of pregnant smokers and nonsmokers and their newborns. Int. J. Vitam. Nutr. Res. 2004, 74, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Scaife, A.R.; McNeill, G.; Campbell, D.M.; Martindale, S.; Devereux, G.; Seaton, A. Maternal intake of antioxidant vitamins in pregnancy in relation to maternal and fetal plasma levels at delivery. Br. J. Nutr. 2006, 95, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Schjoldager, J.G.; Tveden-Nyborg, P.; Lykkesfeldt, J. Prolonged maternal vitamin C deficiency overrides preferential fetal ascorbate transport but does not influence perinatal survival in guinea pigs. Br. J. Nutr. 2013, 110, 1573–1579. [Google Scholar] [CrossRef]
- Juhl, B.; Lauszus, F.F.; Lykkesfeldt, J. Poor Vitamin C Status Late in Pregnancy Is Associated with Increased Risk of Complications in Type 1 Diabetic Women: A Cross-Sectional Study. Nutrients 2017, 9, 186. [Google Scholar] [CrossRef] [PubMed]
- Mikhail, M.S.; Anyaegbunam, A.; Garfinkel, D.; Palan, P.R.; Basu, J.; Romney, S.L. Preeclampsia and antioxidant nutrients: Decreased plasma levels of reduced ascorbic acid, alpha-tocopherol, and beta-carotene in women with preeclampsia. Am. J. Obstet. Gynecol. 1994, 171, 150–157. [Google Scholar] [CrossRef]
- Siega-Riz, A.M.; Promislow, J.H.; Savitz, D.A.; Thorp, J.M., Jr.; McDonald, T. Vitamin C intake and the risk of preterm delivery. Am. J. Obstet. Gynecol. 2003, 189, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.K.; Wise, R.; Yanamandra, K.; Dhanireddy, R.; Bocchini, J.A., Jr. The effect of maternal and cord-blood vitamin C, vitamin E and lipid peroxide levels on newborn birth weight. Mol. Cell. Biochem. 2008, 309, 217–221. [Google Scholar] [CrossRef]
- Lee, B.E.; Hong, Y.C.; Lee, K.H.; Kim, Y.J.; Kim, W.K.; Chang, N.S.; Park, E.A.; Park, H.S.; Hann, H.J. Influence of maternal serum levels of vitamins C and E during the second trimester on birth weight and length. Eur. J. Clin. Nutr. 2004, 58, 1365–1371. [Google Scholar] [CrossRef]
- Jang, W.; Kim, H.; Lee, B.-E.; Chang, N. Maternal fruit and vegetable or vitamin C consumption during pregnancy is associated with fetal growth and infant growth up to 6 months: Results from the Korean Mothers and Children’s Environmental Health (MOCEH) cohort study. Nutr. J. 2018, 17, 105. [Google Scholar] [CrossRef]
- Rumbold, A.R.; Crowther, C.A.; Haslam, R.R.; Dekker, G.A.; Robinson, J.S.; ACTS Study Group. Vitamins C and E and the risks of preeclampsia and perinatal complications. N. Engl. J. Med. 2006, 354, 1796–1806. [Google Scholar] [CrossRef]
- Roberts, J.M.; Myatt, L.; Spong, C.Y.; Thom, E.A.; Hauth, J.C.; Leveno, K.J.; Pearson, G.D.; Wapner, R.J.; Varner, M.; Thorp, J.M., Jr.; et al. Vitamins C and E to prevent complications of pregnancy-associated hypertension. N. Engl. J. Med. 2010, 362, 1282–1291. [Google Scholar] [CrossRef] [PubMed]
- Chappell, L.C.; Seed, P.T.; Briley, A.L.; Kelly, F.J.; Lee, R.; Hunt, B.J.; Parmar, K.; Bewley, S.J.; Shennan, A.H.; Steer, P.J.; et al. Effect of antioxidants on the occurrence of pre-eclampsia in women at increased risk: A randomised trial. Lancet 1999, 354, 810–816. [Google Scholar] [CrossRef] [PubMed]
- Chappell, L.C.; Seed, P.T.; Cstat; Kelly, F.J.; Briley, A.; Hunt, B.J.; Charnock-Jones, D.; Mallet, A.; Poston, L. Vitamin C and E supplementation in women at risk of preeclampsia is associated with changes in indices of oxidative stress and placental function. Am. J. Obstet. Gynecol. 2002, 187, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Kilkenny, C.; Browne, W.J.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Improving bioscience research reporting: The ARRIVE guidelines for reporting animal research. Osteoarthr. Cartil. 2012, 20, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Tveden-Nyborg, P.; Johansen, L.K.; Raida, Z.; Villumsen, C.K.; Larsen, J.O.; Lykkesfeldt, J. Vitamin C deficiency in early postnatal life impairs spatial memory and reduces the number of hippocampal neurons in guinea pigs. Am. J. Clin. Nutr. 2009, 90, 540–546. [Google Scholar] [CrossRef]
- Hasselholt, S.; Tveden-Nyborg, P.; Lykkesfeldt, J. Distribution of vitamin C is tissue specific with early saturation of the brain and adrenal glands following differential oral dose regimens in guinea pigs. Br. J. Nutr. 2015, 113, 1539–1549. [Google Scholar] [CrossRef]
- Tveden-Nyborg, P.; Hasselholt, S.; Miyashita, N.; Moos, T.; Poulsen, H.E.; Lykkesfeldt, J. Chronic vitamin C deficiency does not accelerate oxidative stress in ageing brains of guinea pigs. Basic Clin. Pharmacol. Toxicol. 2012, 110, 524–529. [Google Scholar] [CrossRef]
- Lykkesfeldt, J.; Trueba, G.P.; Poulsen, H.E.; Christen, S. Vitamin C deficiency in weanling guinea pigs: Differential expression of oxidative stress and DNA repair in liver and brain. Br. J. Nutr. 2007, 98, 1116–1119. [Google Scholar] [CrossRef]
- Schjoldager, J.G.; Paidi, M.D.; Lindblad, M.M.; Birck, M.M.; Kjærgaard, A.B.; Dantzer, V.; Lykkesfeldt, J.; Tveden-Nyborg, P. Maternal vitamin C deficiency during pregnancy results in transient fetal and placental growth retardation in guinea pigs. Eur. J. Nutr. 2015, 54, 667–676. [Google Scholar] [CrossRef]
- Tveden-Nyborg, P.; Vogt, L.; Schjoldager, J.G.; Jeannet, N.; Hasselholt, S.; Paidi, M.D.; Christen, S.; Lykkesfeldt, J. Maternal vitamin C deficiency during pregnancy persistently impairs hippocampal neurogenesis in offspring of guinea pigs. PLoS ONE 2012, 7, e48488. [Google Scholar] [CrossRef]
- Clarke, G.L.; Allen, A.M.; Small, J.D.; Lock, A. Subclinical scurvy in the guinea pig. Vet. Pathol. 1980, 17, 40–44. [Google Scholar] [CrossRef]
- Wilson, R.L.; Lampe, K.; Matushewski, B.J.; Regnault, T.R.H.; Jones, H.N. Time Mating Guinea Pigs by Monitoring Changes to the Vaginal Membrane throughout the Estrus Cycle and with Ultrasound Confirmation. Methods Protoc. 2021, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Pullar, J.M.; Bayer, S.; Carr, A.C. Appropriate Handling, Processing and Analysis of Blood Samples is Essential to Avoid Oxidation of Vitamin C to Dehydroascorbic Acid. Antioxidants 2018, 7, 29. [Google Scholar] [CrossRef]
- Carr, A.C.; Pullar, J.M.; Moran, S.; Vissers, M.C.M. Bioavailability of vitamin C from kiwifruit in non-smoking males: Determination of ‘healthy’ and ‘optimal’ intakes. J. Nutr. Sci. 2012, 1, e14. [Google Scholar] [CrossRef] [PubMed]
- Smith-Díaz, C.C.; Magon, N.J.; McKenzie, J.L.; Hampton, M.B.; Vissers, M.C.M.; Das, A.B. Ascorbate Inhibits Proliferation and Promotes Myeloid Differentiation in TP53-Mutant Leukemia. Front. Oncol. 2021, 11, 709543. [Google Scholar] [CrossRef] [PubMed]
- Festing, M.F.W.; Altman, D.G. Guidelines for the design and statistical analysis of experiments using laboratory animals. ILAR J. 2002, 43, 244–258. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.C.; Crombie, G.K.; Palliser, H.K.; Hirst, J.J. Impaired Oligodendrocyte Development Following Preterm Birth: Promoting GABAergic Action to Improve Outcomes. Front. Pediatr. 2021, 9, 618052. [Google Scholar] [CrossRef]
- Morrison, J.L.; Botting, K.J.; Darby, J.R.T.; David, A.L.; Dyson, R.M.; Gatford, K.L.; Gray, C.; Herrera, E.A.; Hirst, J.J.; Kim, B.; et al. Guinea pig models for translation of the developmental origins of health and disease hypothesis into the clinic. J. Physiol. 2018, 596, 5535–5569. [Google Scholar] [CrossRef]
- Chinoy, N.J.; Mehta, R.R.; Seethalakshmi, L.; Sharma, J.D.; Chinoy, M.R. Effects of vitamin C deficiency on physiology of male reproductive organs of guinea pigs. Int. J. Fertil. 1986, 31, 232–239. [Google Scholar]
- Bessesen, D. Changes in organ weights of the guinea pig during experimental scurvy. Am. J. Physiol. Leg. Content 1923, 63, 245–256. [Google Scholar] [CrossRef]
- Hamidian, S.; Talebi, A.R.; Fesahat, F.; Bayat, M.; Mirjalili, A.M.; Ashrafzadeh, H.R.; Rajabi, M.; Montazeri, F.; Babaei, S. The effect of vitamin C on the gene expression profile of sperm protamines in the male partners of couples with recurrent pregnancy loss: A randomized clinical trial. Clin. Exp. Reprod. Med. 2020, 47, 68–76. [Google Scholar] [CrossRef] [PubMed]
- Rafiee, B.; Morowvat, M.H.; Rahimi-Ghalati, N. Comparing the Effectiveness of Dietary Vitamin C and Exercise Interventions on Fertility Parameters in Normal Obese Men. Urol. J. 2016, 13, 2635–2639. [Google Scholar] [PubMed]
- Coker, S.J.; Smith-Díaz, C.C.; Dyson, R.M.; Vissers, M.C.M.; Berry, M.J. The Epigenetic Role of Vitamin C in Neurodevelopment. Int. J. Mol. Sci. 2022, 23, 1208. [Google Scholar] [CrossRef] [PubMed]
- Blaschke, K.; Ebata, K.; Karimi, M.M.; Zepeda-Martínez, J.A.; Goyal, P.; Mahapatra, S.; Tam, A.; Laird, D.J.; Hirst, M.; Rao, A.; et al. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature 2013, 500, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Yin, R.; Mao, S.-Q.; Zhao, B.; Chong, Z.; Yang, Y.; Zhao, C.; Zhang, D.; Huang, H.; Gao, J.; Li, Z.; et al. Ascorbic acid enhances Tet-mediated 5-methylcytosine oxidation and promotes DNA demethylation in mammals. J. Am. Chem. Soc. 2013, 135, 10396–10403. [Google Scholar] [CrossRef]
- Ni, K.; Dansranjavin, T.; Rogenhofer, N.; Oeztuerk, N.; Deuker, J.; Bergmann, M.; Schuppe, H.-C.; Wagenlehner, F.; Weidner, W.; Steger, K.; et al. TET enzymes are successively expressed during human spermatogenesis and their expression level is pivotal for male fertility. Hum. Reprod. 2016, 31, 1411–1424. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Sadeghi, H.M.; Adeli, I.; Calina, D.; Docea, A.O.; Mousavi, T.; Daniali, M.; Nikfar, S.; Tsatsakis, A.; Abdollahi, M. Polycystic Ovary Syndrome: A Comprehensive Review of Pathogenesis, Management, and Drug Repurposing. Int. J. Mol. Sci. 2022, 23, 583. [Google Scholar] [CrossRef]
- Igarashi, M. Augmentative effect of ascorbic acid upon induction of human ovulation in clomiphene-ineffective anovulatory women. Int. J. Fertil. 1977, 22, 168–173. [Google Scholar]
- Habibzadeh, N.; Schorah, C.J.; Smithells, R.W. The effects of maternal folic acid and vitamin C nutrition in early pregnancy on reproductive performance in the guinea-pig. Br. J. Nutr. 1986, 55, 23–35. [Google Scholar] [CrossRef]
- Alves, C.; Rapp, A. Spontaneous Abortion; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- McCollin, A.; Swann, R.L.; Summers, M.C.; Handyside, A.H.; Ottolini, C.S. Abnormal cleavage and developmental arrest of human preimplantation embryos in vitro. Eur. J. Med. Genet. 2020, 63, 103651. [Google Scholar] [CrossRef]
- Gu, T.-P.; Guo, F.; Yang, H.; Wu, H.-P.; Xu, G.-F.; Liu, W.; Xie, Z.-G.; Shi, L.; He, X.; Jin, S.-G.; et al. The role of Tet3 DNA dioxygenase in epigenetic reprogramming by oocytes. Nature 2011, 477, 606–610. [Google Scholar] [CrossRef]
- Khoueiry, R.; Sohni, A.; Thienpont, B.; Luo, X.; Velde, J.V.; Bartoccetti, M.; Boeckx, B.; Zwijsen, A.; Rao, A.; Lambrechts, D.; et al. Lineage-specific functions of TET1 in the postimplantation mouse embryo. Nat. Genet. 2017, 49, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Chu, M.; Yao, F.; Xi, G.; Yang, J.; Zhang, Z.; Yang, Q.; Tian, J.; An, L. Vitamin C Rescues in vitro Embryonic Development by Correcting Impaired Active DNA Demethylation. Front. Cell Dev. Biol. 2021, 9, 784244. [Google Scholar] [CrossRef] [PubMed]
- Chronopoulou, E.; Harper, J.C. IVF culture media: Past, present and future. Hum. Reprod. Update 2015, 21, 39–55. [Google Scholar] [CrossRef] [PubMed]
- DiTroia, S.P.; Percharde, M.; Guerquin, M.-J.; Wall, E.; Collignon, E.; Ebata, K.T.; Mesh, K.; Mahesula, S.; Agathocleous, M.; Laird, D.J.; et al. Maternal vitamin C regulates reprogramming of DNA methylation and germline development. Nature 2019, 573, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Kawahori, K.; Kondo, Y.; Yuan, X.; Kawasaki, Y.; Hanzawa, N.; Tsujimoto, K.; Wada, F.; Kohda, T.; Ishigami, A.; Yamada, T.; et al. Ascorbic acid during the suckling period is required for proper DNA demethylation in the liver. Sci. Rep. 2020, 10, 21228. [Google Scholar] [CrossRef]
- Monsen, E.R. Dietary reference intakes for the antioxidant nutrients: Vitamin C, vitamin E, selenium, and carotenoids. J. Am. Diet. Assoc. 2000, 100, 637–640. [Google Scholar] [CrossRef]
- Dror, D.K.; Allen, L.H. Overview of Nutrients in Human Milk. Adv. Nutr. 2018, 9 (Suppl. S1), 278S–294S. [Google Scholar] [CrossRef]
- Ahmed, L.; Islam, S.; Khan, N.; Nahid, S. Vitamin C content in human milk (colostrum, transitional and mature) and serum of a sample of bangladeshi mothers. Malays. J. Nutr. 2004, 10, 1–4. [Google Scholar]
- Bakshi, S.; Paswan, V.K.; Yadav, S.P.; Bhinchhar, B.K.; Kharkwal, S.; Rose, H.; Kanetkar, P.; Kumar, V.; Al-Zamani, Z.A.S.; Bunkar, D.S. A comprehensive review on infant formula: Nutritional and functional constituents, recent trends in processing and its impact on infants’ gut microbiota. Front. Nutr. 2023, 10, 1194679. [Google Scholar] [CrossRef] [PubMed]
- Francis, J.; Rogers, K.; Brewer, P.; Dickton, D.; Pardini, R. Comparative analysis of ascorbic acid in human milk and infant formula using varied milk delivery systems. Int. Breastfeed. J. 2008, 3, 19. [Google Scholar] [CrossRef]
- Karra, M.V.; Udipi, S.; Kirksey, A.; Roepke, J.L.B. Changes in specific nutrients in breast milk during extended lactation. Am. J. Clin. Nutr. 1986, 43, 495–503. [Google Scholar] [CrossRef] [PubMed]
- Bates, C.; Prentice, A.; Paul, A.; Whitehead, R. Seasonal variations in ascorbic acid status and breast milk ascorbic acid levels in rural Gambian women in relation to dietary intake. Trans. R. Soc. Trop. Med. Hyg. 1982, 76, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Tawfeek, H.I.; Muhyaddin, O.M.; Al-Sanwi, H.I.; Al-Baety, N. Effect of maternal dietary vitamin C intake on the level of vitamin C in breastmilk among nursing mothers in Baghdad, Iraq. Food Nutr. Bull. 2002, 23, 244–247. [Google Scholar] [CrossRef]
- Schleicher, R.L.; Carroll, M.D.; Ford, E.S.; Lacher, D.A. Serum vitamin C and the prevalence of vitamin C deficiency in the United States: 2003–2004 National Health and Nutrition Examination Survey (NHANES). Am. J. Clin. Nutr. 2009, 90, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.C.; Frei, B. Toward a new recommended dietary allowance for vitamin C based on antioxidant and health effects in humans. Am. J. Clin. Nutr. 1999, 69, 1086–1107. [Google Scholar] [CrossRef]
- Carr, A.C.; Lykkesfeldt, J. Factors Affecting the Vitamin C Dose-Concentration Relationship: Implications for Global Vitamin C Dietary Recommendations. Nutrients 2023, 15, 1657. [Google Scholar] [CrossRef]
- Carr, A.C.; Lykkesfeldt, J. Discrepancies in global vitamin C recommendations: A review of RDA criteria and underlying health perspectives. Crit. Rev. Food Sci. Nutr. 2021, 61, 742–755. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Optimal Vitamin C | (n) | Low Vitamin C | (n) |
|---|---|---|---|---|
| Male weight (g) at enrolment | 463.3 ± 86.3 | 13 | 489.5 ± 57.9 | 17 |
| Sire weight (g) at mating | 744.6 ± 132.4 | 34 | 734.9 ± 104.4 | 36 |
| Sire age (weeks) at mating | 16.5 ± 3.1 | 34 | 17.8 ± 3.6 | 36 |
| Female weight (g) at enrolment | 451.9 ± 49.0 | 35 | 432.7 ± 48.1 | 38 |
| Dam weight (g) at mating | 632.7 ± 111.8 | 34 | 588.5 ± 85.9 | 36 |
| Dam age (weeks) at mating | 14.9 ± 4.0 | 34 | 16.1 ± 5.2 | 36 |
| Testis-to-body weight | 0.228 ± 0.030 | 5 | 0.217 ± 0.029 | 6 |
| Ovary-to-body weight | 0.011 ± 0.003 | 5 | 0.009 ± 0.002 | 7 |
| Characteristics | Optimal Vitamin C | (n) | Low Vitamin C | (n) |
|---|---|---|---|---|
| Oestrous cycle length (days) | 15.9 ± 0.7 | 35 | 15.8 ± 0.7 | 38 |
| Unsuccessful matings (n = total matings) | 30 (34.9%) | 86 | 62 (51.7%) * | 120 |
| Failure to establish pregnancy (n = total females) | 1 (2.9%) | 35 | 2 (5.3%) | 38 |
| Failure to sire progeny (n = total males) | 1 (7.7%) | 13 | 4 (23.5%) | 17 |
| Reproductive success rate (n = total animals) | 95.8% | 48 | 89.1% | 55 |
| Outcomes | Optimal Vitamin C | (n) | Low Vitamin C | (n) |
|---|---|---|---|---|
| Miscarriage (delivery before GA62) (n = total pregnancies) | 4 (11.8%) | 34 | 2 (5.6%) | 36 |
| Foetal reabsorption (n = total pregnancies) | 2 (5.9%) | 34 | 9 (25%) * | 36 |
| Preterm delivery (born GA62–66) (n = litters born GA62+) | 0 | 30 | 2 (5.9%) | 34 |
| Stillbirth # of pregnancies (n = litters born GA62+) | 5 (16.7%) | 30 | 9 (26.5%) | 34 |
| Cumulative adverse outcome (n = total pregnancies) | 11 (32.4%) | 34 | 22 (61.1%) * | 36 |
| Stillbirth # of pups (n = total pups) | 6 (5.9%) | 102 | 11 (13.1%) | 85 |
| Litter size (n pups) (n = term litters (born GA66+)) | 3.4 ± 1.0 (2–6) | 30 | 2.6 ± 0.9 (1–4) * | 32 |
| GA of pups at delivery (n = term litters of 2–4 pups) † | 68.6 ± 1.0 | 27 | 68.9 ± 1.2 | 27 |
| Litter birth weight (g) (n = term litters of 2–4 pups) † | 95.3 ± 10.4 | 27 | 97.2 ± 10.0 | 27 |
| Pup sexes (n = total liveborn pups) | 49♂ 47♀ | 96 | 37♂ 37♀ | 74 |
| Characteristics | Males | Females | ||
|---|---|---|---|---|
| Optimal (n = 44) | Low (n = 36) | Optimal (n = 42) | Low (n = 35) | |
| Body weight (g) | 93.3 ± 14.8 | 96.3 ± 13.2 | 91.4 ± 12.9 | 93.5 ± 14.8 |
| Crown-rump (mm) | 128.8 ± 8.8 | 129.0 ± 9.0 | 127.7 ± 8.7 | 126.0 ± 8.7 |
| Hind limb (mm) | 38.5 ± 5.0 | 35.1 ± 3.7 ** | 36.7 ± 4.3 | 34.7 ± 3.6 |
| Hock-toe (mm) | 38.2 ± 5.1 | 36.8 ± 3.1 | 37.3 ± 4.6 | 35.0 ± 3.6 |
| Ponderal Index (PI) | 20.2 ± 3.4 | 22.3 ± 2.9 * | 20.6 ± 3.3 | 23.2 ± 2.7 ** |
| Characteristics | Males | Females | ||
|---|---|---|---|---|
| Optimal (n = 23) | Low (n = 19) | Optimal (n = 22) | Low (n = 18) | |
| Brain-to-body wgt | 2.67 ± 0.43 | 2.58 ± 0.29 | 2.71 ± 0.46 | 2.52 ± 0.30 |
| Liver-to-body wgt | 3.94 ± 0.74 | 3.79 ± 0.36 | 4.26 ± 0.56 | 3.88 ± 0.52 |
| Brain-to-liver ratio | 0.71 ± 0.19 | 0.68 ± 0.11 | 0.66 ± 0.16 | 0.67 ± 0.11 |
| Heart-to-body wgt | 0.41 ± 0.04 | 0.40 ± 0.05 | 0.43 ± 0.04 | 0.40 ± 0.03 |
| Kidney-to-body wgt | 0.43 ± 0.04 | 0.44 ± 0.04 | 0.43 ± 0.04 | 0.44 ± 0.04 |
| Adrenal-to-body wgt | 0.03 ± 0.02 | 0.02 ± 0.01 | 0.02 ± 0.01 | 0.05 ± 0.07 |
| Testis-to-body wgt | 0.07 ± 0.05 | 0.07 ± 0.09 | N/A | N/A |
| Subcut. Fat-to-body wgt | 1.50 ± 0.5 | 1.40 ± 0.3 | 1.34 ± 0.4 | 1.30 ± 0.3 |
| Visc. Fat-to-body wgt | 1.06 ± 0.3 | 1.04 ± 0.4 | 0.73 ± 0.3 | 0.81 ± 0.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coker, S.J.; Dyson, R.M.; Smith-Díaz, C.C.; Vissers, M.C.M.; Berry, M.J. Effects of Low Vitamin C Intake on Fertility Parameters and Pregnancy Outcomes in Guinea Pigs. Nutrients 2023, 15, 4107. https://doi.org/10.3390/nu15194107
Coker SJ, Dyson RM, Smith-Díaz CC, Vissers MCM, Berry MJ. Effects of Low Vitamin C Intake on Fertility Parameters and Pregnancy Outcomes in Guinea Pigs. Nutrients. 2023; 15(19):4107. https://doi.org/10.3390/nu15194107
Chicago/Turabian StyleCoker, Sharna J., Rebecca M. Dyson, Carlos C. Smith-Díaz, Margreet C. M. Vissers, and Mary J. Berry. 2023. "Effects of Low Vitamin C Intake on Fertility Parameters and Pregnancy Outcomes in Guinea Pigs" Nutrients 15, no. 19: 4107. https://doi.org/10.3390/nu15194107