
Mendelian Randomization Study of Lipid Metabolites Reveals Causal Associations with Heel Bone Mineral Density

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Data Source
2.2. Instrument Variables Selection
2.3. Metabolomic Pathway Analysis
2.4. Mendelian Randomization Analysis
2.5. Analysis Software
3. Results
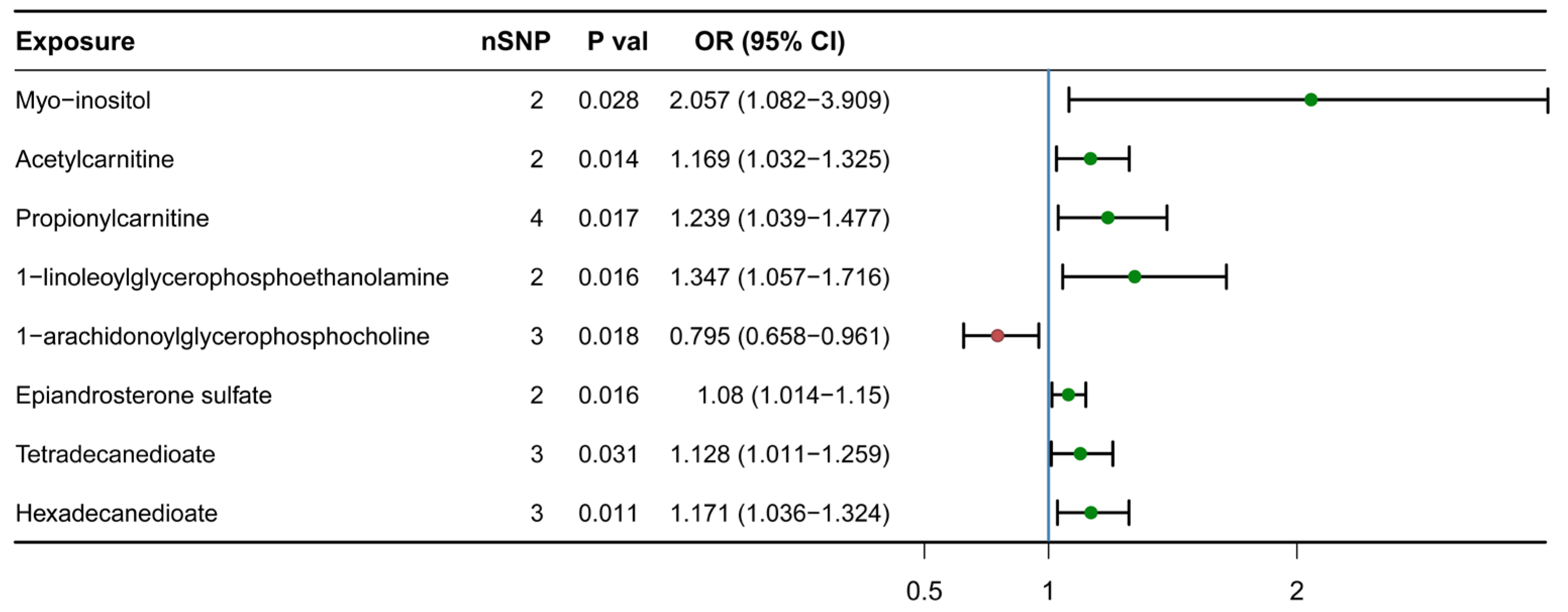
3.1. Causal Association Identified between Eight Lipid Metabolites and H-BMD
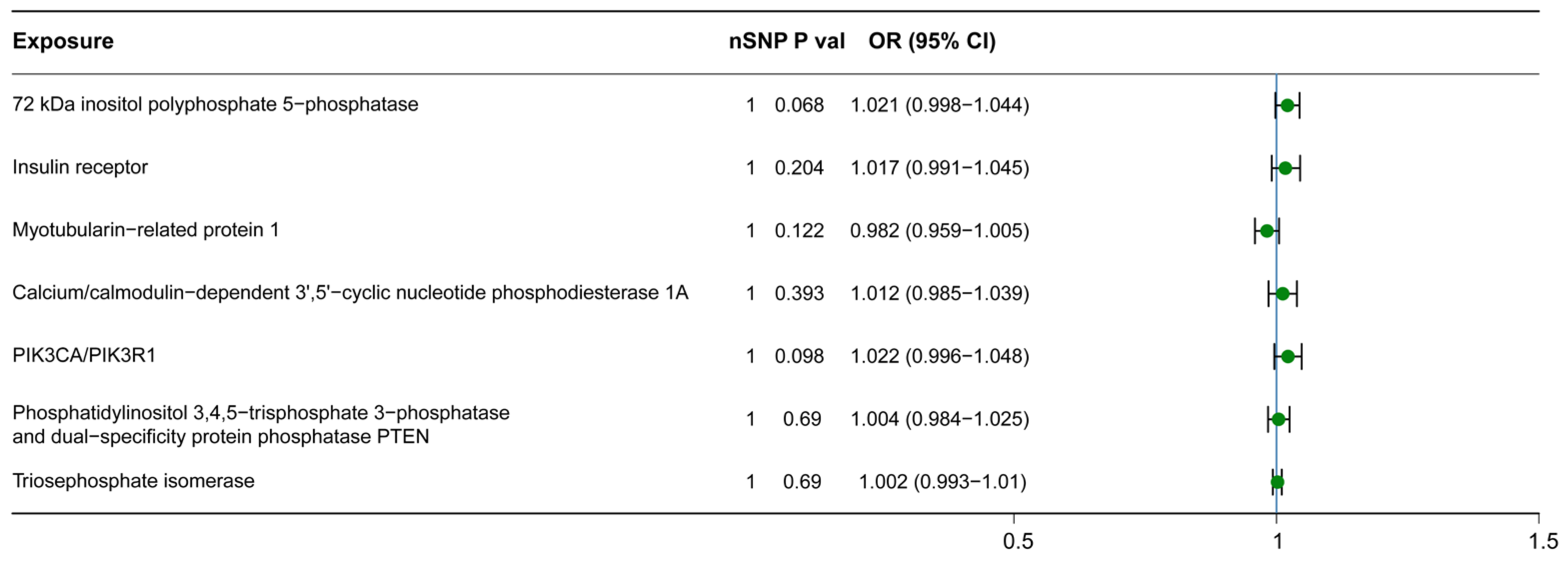
3.2. Causal Relationship between Downstream Metabolites and H-BMD
3.3. The Robustness of MR Results: Sensitivity Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Black, D.M.; Rosen, C.J. Clinical Practice. Postmenopausal Osteoporosis. N. Engl. J. Med. 2016, 374, 254–262. [Google Scholar] [CrossRef]
- Nazrun, A.S.; Tzar, M.N.; Mokhtar, S.A.; Mohamed, I.N. A systematic review of the outcomes of osteoporotic fracture patients after hospital discharge: Morbidity, subsequent fractures, and mortality. Ther. Clin. Risk Manag. 2014, 10, 937–948. [Google Scholar]
- Small, R.E. Uses and limitations of bone mineral density measurements in the management of osteoporosis. MedGenMed Medscape Gen. Med. 2005, 7, 3. [Google Scholar]
- Kanis, J.A.; Melton, L.J., 3rd; Christiansen, C.; Johnston, C.C.; Khaltaev, N. The diagnosis of osteoporosis. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1994, 9, 1137–1141. [Google Scholar] [CrossRef]
- Grampp, S.; Genant, H.K.; Mathur, A.; Lang, P.; Jergas, M.; Takada, M.; Glüer, C.C.; Lu, Y.; Chavez, M. Comparisons of noninvasive bone mineral measurements in assessing age-related loss, fracture discrimination, and diagnostic classification. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 1997, 12, 697–711. [Google Scholar] [CrossRef]
- Jiang, C.; Giger, M.L.; Kwak, S.M.; Chinander, M.R.; Martell, J.M.; Favus, M.J. Normalized BMD as a predictor of bone strength. Acad. Radiol. 2000, 7, 33–39. [Google Scholar] [CrossRef]
- Bainbridge, N.J.; Davie, M.W.; Haddaway, M.J. Bone loss after stroke over 52 weeks at os calcis: Influence of sex, mobility and relation to bone density at other sites. Age Ageing 2006, 35, 127–132. [Google Scholar] [CrossRef]
- Khaw, K.T.; Reeve, J.; Luben, R.; Bingham, S.; Welch, A.; Wareham, N.; Oakes, S.; Day, N. Prediction of total and hip fracture risk in men and women by quantitative ultrasound of the calcaneus: EPIC-Norfolk prospective population study. Lancet 2004, 363, 197–202. [Google Scholar] [CrossRef]
- Yamada, M.; Ito, M.; Hayashi, K.; Nakamura, T. Calcaneus as a site for assessment of bone mineral density: Evaluation in cadavers and healthy volunteers. AJR. Am. J. Roentgenol. 1993, 161, 621–627. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, C.; Liang, H.; Zhou, N.; Huang, T.; Zhao, Z.; Luo, X. Polyunsaturated Fatty Acids Level and Bone Mineral Density: A Two-Sample Mendelian Randomization Study. Front. Endocrinol. 2022, 13, 858851. [Google Scholar] [CrossRef]
- Kan, B.; Zhao, Q.; Wang, L.; Xue, S.; Cai, H.; Yang, S. Association between lipid biomarkers and osteoporosis: A cross-sectional study. BMC Musculoskelet. Disord. 2021, 22, 759. [Google Scholar] [CrossRef] [PubMed]
- Orozco, P. Atherogenic lipid profile and elevated lipoprotein (a) are associated with lower bone mineral density in early postmenopausal overweight women. Eur. J. Epidemiol. 2004, 19, 1105–1112. [Google Scholar] [CrossRef] [PubMed]
- Di Pietro, G.; Capuani, S.; Manenti, G.; Vinicola, V.; Fusco, A.; Baldi, J.; Scimeca, M.; Hagberg, G.; Bozzali, M.; Simonetti, G.; et al. Bone Marrow Lipid Profiles from Peripheral Skeleton as Potential Biomarkers for Osteoporosis: A 1H-MR Spectroscopy Study. Acad. Radiol. 2016, 23, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Mattioli, D.; Vinicola, V.; Aragona, M.; Montuori, M.; Tarantino, U.; Capuani, S. Behavior during aging of bone-marrow fatty-acids profile in women’s calcaneus to search for early potential osteoporotic biomarkers: A 1H-MR Spectroscopy study. Bone 2022, 164, 116514. [Google Scholar] [CrossRef]
- Evans, D.M.; Davey Smith, G. Mendelian Randomization: New Applications in the Coming Age of Hypothesis-Free Causality. Annu. Rev. Genom. Hum. Genet. 2015, 16, 327–350. [Google Scholar] [CrossRef]
- Emdin, C.A.; Khera, A.V.; Kathiresan, S. Mendelian Randomization. JAMA 2017, 318, 1925–1926. [Google Scholar] [CrossRef]
- Minelli, C.; Del Greco, M.F.; van der Plaat, D.A.; Bowden, J.; Sheehan, N.A.; Thompson, J. The use of two-sample methods for Mendelian randomization analyses on single large datasets. Int. J. Epidemiol. 2021, 50, 1651–1659. [Google Scholar] [CrossRef]
- Shin, S.Y.; Fauman, E.B.; Petersen, A.K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.P.; et al. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef]
- Morris, J.A.; Kemp, J.P.; Youlten, S.E.; Laurent, L.; Logan, J.G.; Chai, R.C.; Vulpescu, N.A.; Forgetta, V.; Kleinman, A.; Mohanty, S.T.; et al. An atlas of genetic influences on osteoporosis in humans and mice. Nat. Genet. 2019, 51, 258–266. [Google Scholar] [CrossRef]
- von Elm, E.; Altman, D.G.; Egger, M.; Pocock, S.J.; Gøtzsche, P.C.; Vandenbroucke, J.P. The Strengthening the Reporting of Observational Studies in Epidemiology (STROBE) statement: Guidelines for reporting observational studies. Lancet 2007, 370, 1453–1457. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 2011, 40, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Scott, R.A.; Timpson, N.J.; Davey Smith, G.; Thompson, S.G. Using published data in Mendelian randomization: A blueprint for efficient identification of causal risk factors. Eur. J. Epidemiol. 2015, 30, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Burgess, S.; Thompson, S.G. Interpreting findings from Mendelian randomization using the MR-Egger method. Eur. J. Epidemiol. 2017, 32, 377–389. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef] [PubMed]
- Milne, R.L.; Kuchenbaecker, K.B.; Michailidou, K.; Beesley, J.; Kar, S.; Lindström, S.; Hui, S.; Lemaçon, A.; Soucy, P.; Dennis, J.; et al. Identification of ten variants associated with risk of estrogen-receptor-negative breast cancer. Nat. Genet. 2017, 49, 1767–1778. [Google Scholar] [CrossRef]
- Hartwig, F.P.; Davey Smith, G.; Bowden, J. Robust inference in summary data Mendelian randomization via the zero modal pleiotropy assumption. Int. J. Epidemiol. 2017, 46, 1985–1998. [Google Scholar] [CrossRef]
- Burgess, S.; Dudbridge, F.; Thompson, S.G. Combining information on multiple instrumental variables in Mendelian randomization: Comparison of allele score and summarized data methods. Stat. Med. 2016, 35, 1880–1906. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef]
- Nowak, C.; Ärnlöv, J. A Mendelian randomization study of the effects of blood lipids on breast cancer risk. Nat. Commun. 2018, 9, 3957. [Google Scholar] [CrossRef]
- Bowden, J.; Del Greco, M.F.; Minelli, C.; Davey Smith, G.; Sheehan, N.A.; Thompson, J.R. Assessing the suitability of summary data for two-sample Mendelian randomization analyses using MR-Egger regression: The role of the I2 statistic. Int. J. Epidemiol. 2016, 45, 1961–1974. [Google Scholar] [CrossRef]
- Hemani, G.; Zheng, J.; Elsworth, B.; Wade, K.H.; Haberland, V.; Baird, D.; Laurin, C.; Burgess, S.; Bowden, J.; Langdon, R.; et al. The MR-Base platform supports systematic causal inference across the human phenome. eLife 2018, 7, e34408. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; He, W. Metabolome-Wide Mendelian Randomization Assessing the Causal Relationship between Blood Metabolites and Bone Mineral Density. Calcif. Tissue Int. 2023, 112, 543–562. [Google Scholar] [CrossRef] [PubMed]
- Tao, S.S.; Wang, P.; Wang, X.Y.; Yin, K.J.; Yang, X.K.; Wang, Z.X.; Wang, D.G.; Pan, H.F. Causal effect of polyunsaturated fatty acids on bone mineral density and fracture. Front. Nutr. 2022, 9, 1014847. [Google Scholar] [CrossRef] [PubMed]
- Izzo, L.T.; Trefely, S.; Demetriadou, C.; Drummond, J.M.; Mizukami, T.; Kuprasertkul, N.; Farria, A.T.; Nguyen, P.T.T.; Murali, N.; Reich, L.; et al. Acetylcarnitine shuttling links mitochondrial metabolism to histone acetylation and lipogenesis. Sci. Adv. 2023, 9, eadf0115. [Google Scholar] [CrossRef]
- Bohmer, T.; Bremer, J. Propionylcarnitine. Physiological variations in vivo. Biochim. Biophys. Acta 1968, 152, 559–567. [Google Scholar] [CrossRef]
- Rosca, M.G.; Lemieux, H.; Hoppel, C.L. Mitochondria in the elderly: Is acetylcarnitine a rejuvenator? Adv. Drug Deliv. Rev. 2009, 61, 1332–1342. [Google Scholar] [CrossRef]
- Müller, F.; Sharma, A.; König, J.; Fromm, M.F. Biomarkers for In Vivo Assessment of Transporter Function. Pharmacol. Rev. 2018, 70, 246–277. [Google Scholar] [CrossRef]
- Sun, D.; Tiedt, S.; Yu, B.; Jian, X.; Gottesman, R.F.; Mosley, T.H.; Boerwinkle, E.; Dichgans, M.; Fornage, M. A prospective study of serum metabolites and risk of ischemic stroke. Neurology 2019, 92, e1890–e1898. [Google Scholar] [CrossRef]
- Sigal, S.H.; Yandrasitz, J.R.; Berry, G.T. Kinetic evidence for compartmentalization of myo-inositol in hepatocytes. Metabolism 1993, 42, 395–401. [Google Scholar] [CrossRef]
- Chhetri, D.R. Myo-Inositol and Its Derivatives: Their Emerging Role in the Treatment of Human Diseases. Front. Pharmacol. 2019, 10, 1172. [Google Scholar] [CrossRef]
- Chau, J.F.; Lee, M.K.; Law, J.W.; Chung, S.K.; Chung, S.S. Sodium/myo-inositol cotransporter-1 is essential for the development and function of the peripheral nerves. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2005, 19, 1887–1889. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Chung, S.K.; Miao, D.; Lau, K.S.; Chan, A.W.; Kung, A.W. Sodium/myo-inositol cotransporter 1 and myo-inositol are essential for osteogenesis and bone formation. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2011, 26, 582–590. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Guijo, A.; Neunzig, J.; Gerber, A.; Oji, V.; Hartmann, M.F.; Schuppe, H.C.; Traupe, H.; Bernhardt, R.; Wudy, S.A. Role of steroid sulfatase in steroid homeostasis and characterization of the sulfated steroid pathway: Evidence from steroid sulfatase deficiency. Mol. Cell. Endocrinol. 2016, 437, 142–153. [Google Scholar] [CrossRef]
- Piper, T.; Putz, M.; Schänzer, W.; Pop, V.; McLeod, M.D.; Uduwela, D.R.; Stevenson, B.J.; Thevis, M. Epiandrosterone sulfate prolongs the detectability of testosterone, 4-androstenedione, and dihydrotestosterone misuse by means of carbon isotope ratio mass spectrometry. Drug Test. Anal. 2017, 9, 1695–1703. [Google Scholar] [CrossRef]
- Esquivel, A.; Alechaga, É.; Monfort, N.; Ventura, R. Sulfate metabolites improve retrospectivity after oral testosterone administration. Drug Test. Anal. 2019, 11, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Moayyeri, A.; Cheung, C.L.; Tan, K.C.; Morris, J.A.; Cerani, A.; Mohney, R.P.; Richards, J.B.; Hammond, C.; Spector, T.D.; Menni, C. Metabolomic Pathways to Osteoporosis in Middle-Aged Women: A Genome-Metabolome-Wide Mendelian Randomization Study. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2018, 33, 643–650. [Google Scholar] [CrossRef]
- Song, F.; Xie, T.; Liu, X.; Chin, B.; Luo, X.; Liao, S.; Feng, W.; He, M.; Huang, N.; Su, Z.; et al. UPLC/Q-TOF-MS-based Metabolomics Study of the Antiosteoporosis Effects of Vaccarin in Ovariectomized Mice. Planta Medica 2023, 89, 218–230. [Google Scholar] [CrossRef]
- Calzada, E.; Onguka, O.; Claypool, S.M. Phosphatidylethanolamine Metabolism in Health and Disease. Int. Rev. Cell Mol. Biol. 2016, 321, 29–88. [Google Scholar] [CrossRef]
- Bazzi, M.D.; Youakim, M.A.; Nelsestuen, G.L. Importance of phosphatidylethanolamine for association of protein kinase C and other cytoplasmic proteins with membranes. Biochemistry 1992, 31, 1125–1134. [Google Scholar] [CrossRef]
- Luthringer, B.J.; Katha, U.M.; Willumeit, R. Phosphatidylethanolamine biomimetic coating increases mesenchymal stem cell osteoblastogenesis. J. Mater. Sci. Mater. Med. 2014, 25, 2561–2571. [Google Scholar] [CrossRef]
- Bispo, D.S.C.; Michálková, L.; Correia, M.; Jesus, C.S.H.; Duarte, I.F.; Goodfellow, B.J.; Oliveira, M.B.; Mano, J.F.; Gil, A.M. Endo- and Exometabolome Crosstalk in Mesenchymal Stem Cells Undergoing Osteogenic Differentiation. Cells 2022, 11, 1257. [Google Scholar] [CrossRef] [PubMed]
- Aleidi, S.M.; Al-Ansari, M.M.; Alnehmi, E.A.; Malkawi, A.K.; Alodaib, A.; Alshaker, M.; Benabdelkamel, H.; Abdel Rahman, A.M. Lipidomics Profiling of Patients with Low Bone Mineral Density (LBMD). Int. J. Mol. Sci. 2022, 23, 2017. [Google Scholar] [CrossRef] [PubMed]
- Irie, A.; Yamamoto, K.; Miki, Y.; Murakami, M. Phosphatidylethanolamine dynamics are required for osteoclast fusion. Sci. Rep. 2017, 7, 46715. [Google Scholar] [CrossRef] [PubMed]
- Altmaier, E.; Fobo, G.; Heier, M.; Thorand, B.; Meisinger, C.; Römisch-Margl, W.; Waldenberger, M.; Gieger, C.; Illig, T.; Adamski, J.; et al. Metabolomics approach reveals effects of antihypertensives and lipid-lowering drugs on the human metabolism. Eur. J. Epidemiol. 2014, 29, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Hui, L.; Sun, L.; Guo, D.; Shi, M.; Zhang, K.; Yang, P.; Wang, Y.; Liu, F.; Shen, O.; et al. Association Between Human Blood Metabolome and the Risk of Psychiatric Disorders. Schizophr. Bull. 2023, 49, 428–443. [Google Scholar] [CrossRef]
- Christiansen, C. Osteoporosis: Diagnosis and management today and tomorrow. Bone 1995, 17, 513s–516s. [Google Scholar] [CrossRef]
- Ensrud, K.E.; Crandall, C.J. Osteoporosis. Ann. Intern. Med. 2017, 167, ITC17–ITC32. [Google Scholar] [CrossRef]
- Taylor, A.; Staruchowicz, L. The Experience and Effectiveness of Nurse Practitioners in Orthopaedic Settings: A Comprehensive Systematic Review. JBI Libr. Syst. Rev. 2012, 10, 1–22. [Google Scholar] [CrossRef]
- Garvey, W.T.; Mechanick, J.I.; Brett, E.M.; Garber, A.J.; Hurley, D.L.; Jastreboff, A.M.; Nadolsky, K.; Pessah-Pollack, R.; Plodkowski, R. American association of clinical endocrinologists and american college of endocrinology comprehensive clinical practice guidelines for medical care of patients with obesity. Endocr. Pract. Off. J. Am. Coll. Endocrinol. Am. Assoc. Clin. Endocrinol. 2016, 22 (Suppl. S3), 1–203. [Google Scholar] [CrossRef]
- Khosla, S.; Hofbauer, L.C. Osteoporosis treatment: Recent developments and ongoing challenges. Lancet Diabetes Endocrinol. 2017, 5, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Tao, Z.; Zhu, Y.; Tao, L. Mapping theme trends and recognizing hot spots in postmenopausal osteoporosis research: A bibliometric analysis. PeerJ 2019, 7, e8145. [Google Scholar] [CrossRef] [PubMed]
- Mundy, G.R. Nutritional modulators of bone remodeling during aging. Am. J. Clin. Nutr. 2006, 83, 427s–430s. [Google Scholar] [CrossRef] [PubMed]
- Tosi, L.L.; Kyle, R.F. Fragility fractures: The fall and decline of bone health. Commentary on “Interventions to improve osteoporosis treatment following hip fracture” by Gardner et al. J. Bone Jt. Surg. 2005, 87, 1–2. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, M.; Du, Y.; Zhang, C.; Li, Z.; Li, Q.; Qi, E.; Ruan, W.; Feng, S.; Zhou, H. Mendelian Randomization Study of Lipid Metabolites Reveals Causal Associations with Heel Bone Mineral Density. Nutrients 2023, 15, 4160. https://doi.org/10.3390/nu15194160
Wu M, Du Y, Zhang C, Li Z, Li Q, Qi E, Ruan W, Feng S, Zhou H. Mendelian Randomization Study of Lipid Metabolites Reveals Causal Associations with Heel Bone Mineral Density. Nutrients. 2023; 15(19):4160. https://doi.org/10.3390/nu15194160
Chicago/Turabian StyleWu, Mingxin, Yufei Du, Chi Zhang, Zhen Li, Qingyang Li, Enlin Qi, Wendong Ruan, Shiqing Feng, and Hengxing Zhou. 2023. "Mendelian Randomization Study of Lipid Metabolites Reveals Causal Associations with Heel Bone Mineral Density" Nutrients 15, no. 19: 4160. https://doi.org/10.3390/nu15194160
APA StyleWu, M., Du, Y., Zhang, C., Li, Z., Li, Q., Qi, E., Ruan, W., Feng, S., & Zhou, H. (2023). Mendelian Randomization Study of Lipid Metabolites Reveals Causal Associations with Heel Bone Mineral Density. Nutrients, 15(19), 4160. https://doi.org/10.3390/nu15194160