
Elucidating the Pharmacological Properties of Zingiber officinale Roscoe (Ginger) on Muscle Ageing by Untargeted Metabolomic Profiling of Human Myoblasts

 ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Replicative Senescence
2.2. Ginger (Zingiber officinale Roscoe) Extract Preparation and Treatment Protocol
2.3. Sample Preparation
2.4. Metabolomic Analysis by UHPLC-MS/MS
2.5. Statistical Analysis
3. Results
3.1. Differential Metabolomic Analysis between Control and Treated Myoblast Group
3.2. Comparison of Metabolites Profile in Control and Ginger-Treated Myoblast Cell
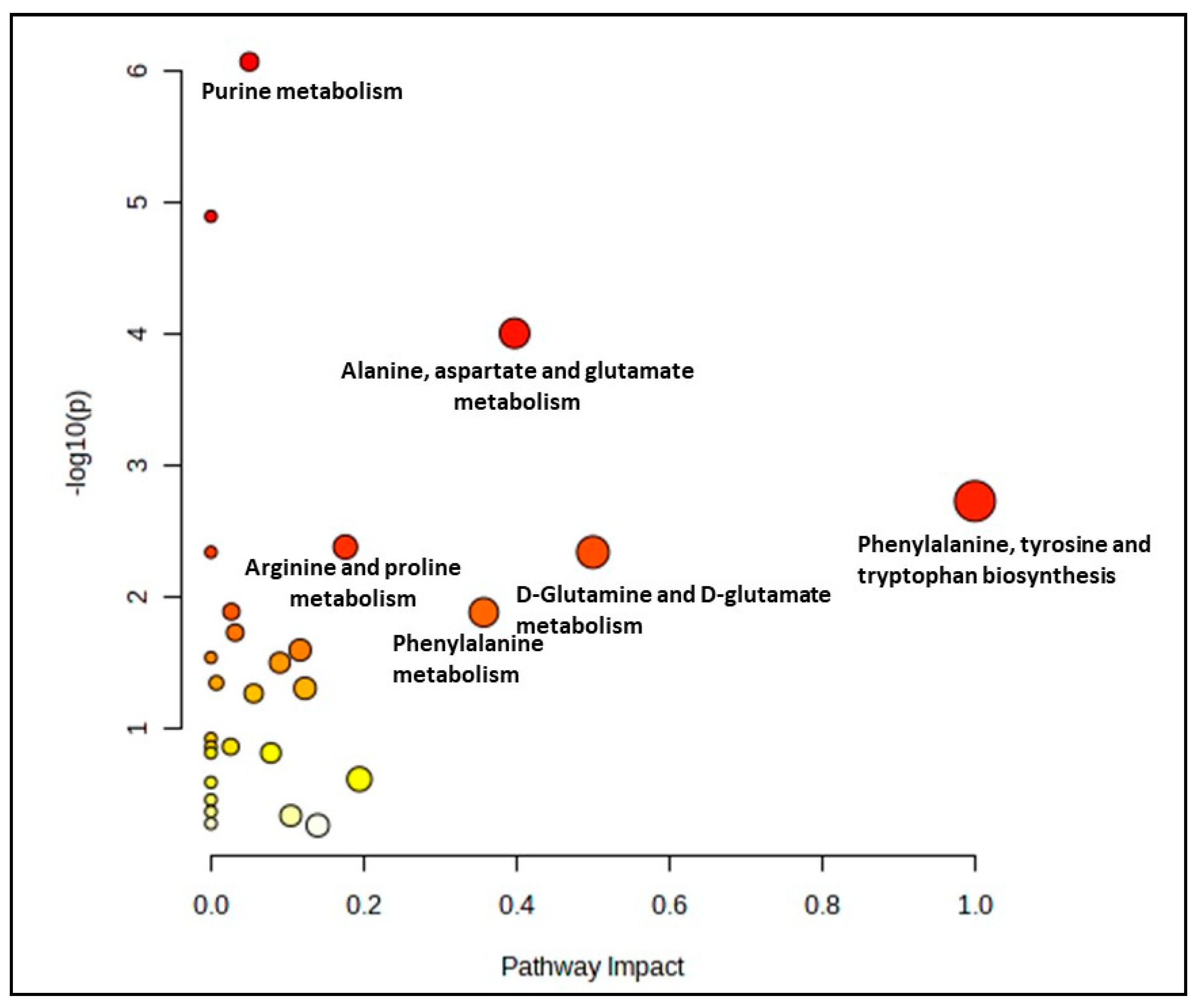
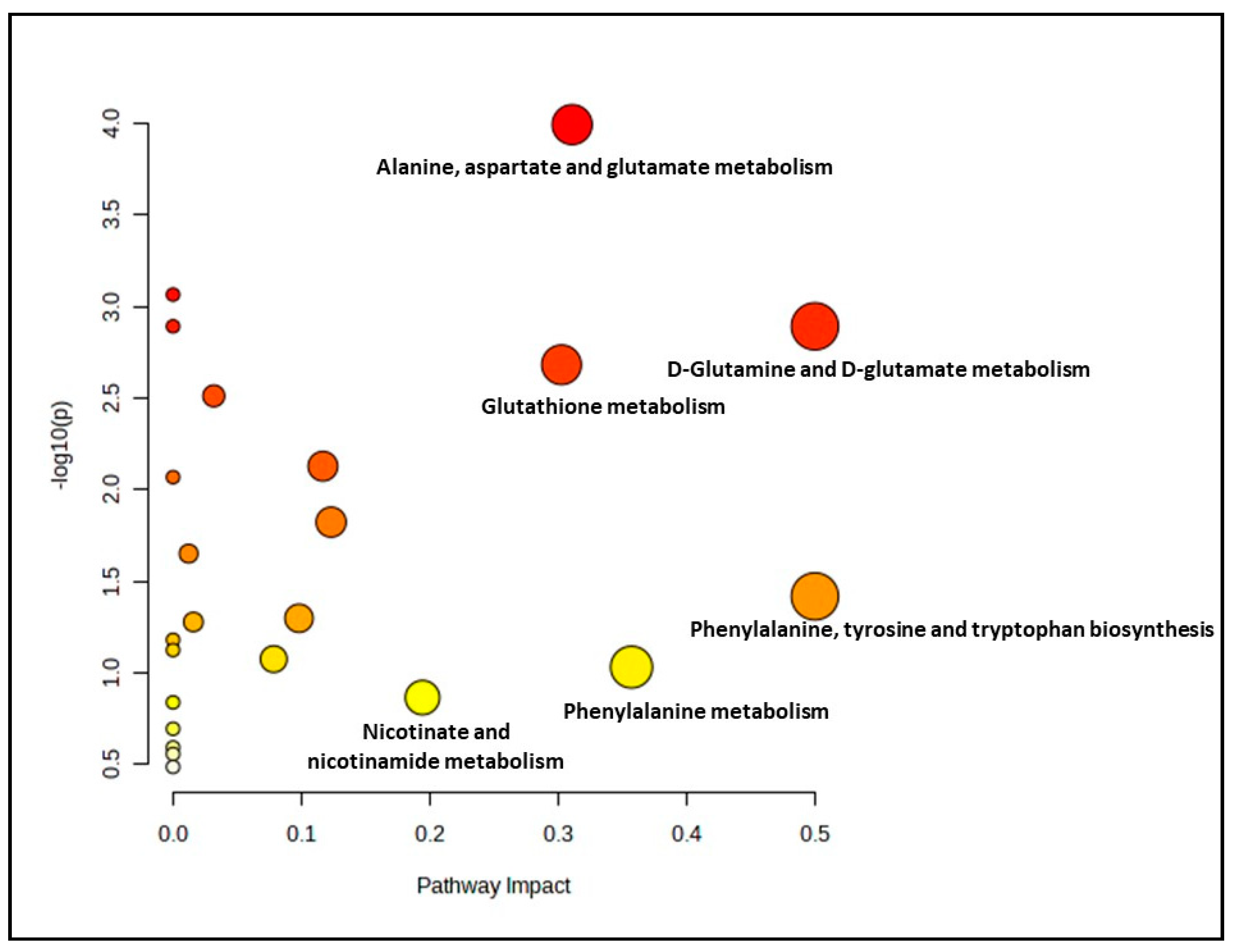
3.3. Biochemical Pathways Analysis of Muscle Metabolomes in Control and Myoblast-Treated Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, A.W.; Ogunwole, S.U.; Blakeslee, L.; Rabe, M.A. The Population 65 Years and Older in the United States: 2016; US Department of Commerce, Economics and Statistics Administration; US Census Bureau: Washington, DC, USA, 2018.
- Hou, Y.; Dan, X.; Babbar, M.; Wei, Y.; Hasselbalch, S.G.; Croteau, D.L.; Bohr, V.A. Ageing as a risk factor for neurodegenerative disease. Nat. Rev. Neurol. 2019, 15, 565–581. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Jentoft, A.J.; Bahat, G.; Bauer, J.; Boirie, Y.; Bruyère, O.; Cederholm, T.; Cooper, C.; Landi, F.; Rolland, Y.; Sayer, A.A. Sarcopenia: Revised European consensus on definition and diagnosis. Age Ageing 2019, 48, 16–31. [Google Scholar] [CrossRef]
- Larsson, L.; Degens, H.; Li, M.; Salviati, L.; Lee, Y.I.; Thompson, W.; Kirkland, J.L.; Sandri, M. Sarcopenia: Aging-Related Loss of Muscle Mass and Function. Physiol. Rev. 2019, 99, 427–511. [Google Scholar] [CrossRef]
- Serra, A.J.; Prokić, M.D.; Vasconsuelo, A.; Pinto, J.R. Oxidative Stress in Muscle Diseases: Current and Future Therapy. Oxid. Med. Cell Longev. 2018, 2018, 6439138. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, B.M.; Banik, B.K.; Borah, P.; Jain, A. Reactive Oxygen Species (ROS): Key Components in Cancer Therapies. Anticancer. Agents Med. Chem. 2022, 22, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, X.; Liu, Y.; Zhang, Z. Effects of Exercise-Induced ROS on the Pathophysiological Functions of Skeletal Muscle. Oxid. Med. Cell Longev. 2021, 2021, 3846122. [Google Scholar] [CrossRef] [PubMed]
- Murata, M. Inflammation and cancer. Environ. Health Prev. Med. 2018, 23, 50. [Google Scholar] [CrossRef]
- Watanabe, H.; Enoki, Y.; Maruyama, T. Sarcopenia in Chronic Kidney Disease: Factors, Mechanisms, and Therapeutic Interventions. Biol. Pharm. Bull. 2019, 42, 1437–1445. [Google Scholar] [CrossRef]
- Cartee, G.D.; Hepple, R.T.; Bamman, M.M.; Zierath, J.R. Exercise Promotes Healthy Aging of Skeletal Muscle. Cell Metab. 2016, 23, 1034–1047. [Google Scholar] [CrossRef]
- Papaioannou, K.G.; Nilsson, A.; Nilsson, L.M.; Kadi, F. Healthy Eating Is Associated with Sarcopenia Risk in Physically Active Older Adults. Nutrients 2021, 13, 2813. [Google Scholar] [CrossRef] [PubMed]
- Tohma, H.; Gülçin, İ.; Bursal, E.; Gören, A.C.; Alwasel, S.H.; Köksal, E. Antioxidant activity and phenolic compounds of ginger (Zingiber officinale Rosc.) determined by HPLC-MS/MS. J. Food Meas. Charact. 2017, 11, 556–566. [Google Scholar] [CrossRef]
- Si, W.; Chen, Y.P.; Zhang, J.; Chen, Z.-Y.; Chung, H.Y. Antioxidant activities of ginger extract and its constituents toward lipids. Food Chem. 2018, 239, 1117–1125. [Google Scholar] [CrossRef] [PubMed]
- Kiyama, R. Nutritional implications of ginger: Chemistry, biological activities and signaling pathways. J. Nutr. Biochem. 2020, 86, 108486. [Google Scholar] [CrossRef]
- Yasodai, R.; Kavimani, M.; Prabhu, K. Phytochemical analysis and quantitative nutritional evaluvation of Zingiber Officinale Roscae (Ginger). Int. J. Pharm. Sci. Res. 2020, 11, 2090–2094. [Google Scholar] [CrossRef]
- Kirdat, P.; Dandge, P.; Hagwane, R.; Nikam, A.; Mahadik, S.; Jirange, S. Synthesis and characterization of ginger (Z. officinale) extract mediated iron oxide nanoparticles and its antibacterial activity. Mater. Today Proc. 2021, 43, 2826–2831. [Google Scholar] [CrossRef]
- Pashaei-Asl, R.; Pashaei-Asl, F.; Mostafa Gharabaghi, P.; Khodadadi, K.; Ebrahimi, M.; Ebrahimie, E.; Pashaiasl, M. The Inhibitory Effect of Ginger Extract on Ovarian Cancer Cell Line; Application of Systems Biology. Adv. Pharm. Bull. 2017, 7, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Ezzat, S.M.; Ezzat, M.I.; Okba, M.M.; Menze, E.T.; Abdel-Naim, A.B. The hidden mechanism beyond ginger (Zingiber officinale Rosc.) potent in vivo and in vitro anti-inflammatory activity. J. Ethnopharmacol. 2018, 214, 113–123. [Google Scholar] [CrossRef]
- Uchitomi, R.; Hatazawa, Y.; Senoo, N.; Yoshioka, K.; Fujita, M.; Shimizu, T.; Miura, S.; Ono, Y.; Kamei, Y. Metabolomic Analysis of Skeletal Muscle in Aged Mice. Sci. Rep. 2019, 9, 10425. [Google Scholar] [CrossRef]
- Dabaj, I.; Ferey, J.; Marguet, F.; Gilard, V.; Basset, C.; Bahri, Y.; Brehin, A.-C.; Vanhulle, C.; Leturcq, F.; Marret, S.; et al. Muscle metabolic remodelling patterns in Duchenne muscular dystrophy revealed by ultra-high-resolution mass spectrometry imaging. Sci. Rep. 2021, 11, 1906. [Google Scholar] [CrossRef]
- Elhassan, Y.S.; Kluckova, K.; Fletcher, R.S.; Schmidt, M.S.; Garten, A.; Doig, C.L.; Cartwright, D.M.; Oakey, L.; Burley, C.V.; Jenkinson, N.; et al. Nicotinamide Riboside Augments the Aged Human Skeletal Muscle NAD(+) Metabolome and Induces Transcriptomic and Anti-inflammatory Signatures. Cell Rep. 2019, 28, 1717–1728.e1716. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, H.; Mukai, K.; Takahashi, Y.; Takahashi, T. Metabolomic analysis of skeletal muscle before and after strenuous exercise to fatigue. Sci. Rep. 2021, 11, 11261. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sahardi, N.F.N.; Jaafar, F.; Mad Nordin, M.F.; Makpol, S. Zingiber Officinale Roscoe Prevents Cellular Senescence of Myoblasts in Culture and Promotes Muscle Regeneration. Evid. Based Complement. Alternat Med. 2020, 2020, 1787342. [Google Scholar] [CrossRef] [PubMed]
- Sarip, M. Subcritical Water Extraction of 6-Gingerol and 6-Shogaol from Zingiber officinale. Master’s Thesis, Universiti Teknologi Malaysia, Skudai, Malaysia, 2012. [Google Scholar]
- Sahardi, N.; Jaafar, F.; Zakaria, S.; Tan, J.; Nordin, M.; Makpol, S. Comparison of the Antioxidant Activity of Malaysian Ginger (Zingiber officinale Roscoe) Extracts with that of Selected Natural Products and its Effect on the Viability of Myoblast Cells in Culture. Sains Malays. 2021, 50, 1445–1456. [Google Scholar] [CrossRef]
- Osman, E.; Mohammad Zahariluddin, A.S.; Sharip, S.; Md Idris, Z.; Tan, J.K. Metabolomic Profiling Reveals Common Metabolic Alterations in Plasma of Patients with Toxoplasma Infection and Schizophrenia. Genes 2022, 13, 1482. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.K.; Zakaria, S.N.A.; Gunasekaran, G.; Abdul Sani, N.F.; Nasaruddin, M.L.; Jaafar, F.; Abu Bakar, Z.H.; Amir Hamzah, A.I.Z.; Nor Aripin, K.N.; Mohd Rani, M.D.; et al. Metabolomics Profiling of Age-Associated Metabolites in Malay Population. Oxid. Med. Cell Longev. 2023, 2023, 4416410. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Using MetaboAnalyst 3.0 for Comprehensive Metabolomics Data Analysis. Curr. Protoc. Bioinform. 2016, 55, 14.10.11–14.10.91. [Google Scholar] [CrossRef]
- Ju, J.-H.; Oh, B.-R.; Heo, S.-Y.; Lee, Y.-U.; Shon, J.-h.; Kim, C.-H.; Kim, Y.-M.; Seo, J.-W.; Hong, W.-K. Production of adipic acid by short- and long-chain fatty acid acyl-CoA oxidase engineered in yeast Candida tropicalis. Bioprocess. Biosyst. Eng. 2020, 43, 33–43. [Google Scholar] [CrossRef]
- Matthews, D.E. Review of Lysine Metabolism with a Focus on Humans. J. Nutr. 2020, 150, S2548–S2555. [Google Scholar] [CrossRef]
- Munasinghe, M.; Afshari, R.; Heydarian, D.; Almotayri, A.; Dias, D.A.; Thomas, J.; Jois, M. Effects of cocoa on altered metabolite levels in purine metabolism pathways and urea cycle in Alzheimer’s disease in C. elegans. Transl. Med. Aging 2022, 6, 14–24. [Google Scholar] [CrossRef]
- Wanders, R.J.A.; Visser, G.; Ferdinandusse, S.; Vaz, F.M.; Houtkooper, R.H. Mitochondrial Fatty Acid Oxidation Disorders: Laboratory Diagnosis, Pathogenesis, and the Complicated Route to Treatment. J. Lipid Atheroscler. 2020, 9, 313–333. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Koves, T.R.; Hubal, M.J.; Abouassi, H.; Beri, N.; Bateman, L.A.; Stevens, R.D.; Ilkayeva, O.R.; Hoffman, E.P.; Muoio, D.M.; et al. Metabolite signatures of exercise training in human skeletal muscle relate to mitochondrial remodelling and cardiometabolic fitness. Diabetologia 2014, 57, 2282–2295. [Google Scholar] [CrossRef] [PubMed]
- Chouchani, E.T.; Pell, V.R.; Gaude, E.; Aksentijević, D.; Sundier, S.Y.; Robb, E.L.; Logan, A.; Nadtochiy, S.M.; Ord, E.N.J.; Smith, A.C.; et al. Ischaemic accumulation of succinate controls reperfusion injury through mitochondrial ROS. Nature 2014, 515, 431–435. [Google Scholar] [CrossRef]
- Wang, T.; Xu, Y.Q.; Yuan, Y.X.; Xu, P.W.; Zhang, C.; Li, F.; Wang, L.N.; Yin, C.; Zhang, L.; Cai, X.C.; et al. Succinate induces skeletal muscle fiber remodeling via SUNCR1 signaling. EMBO Rep. 2019, 20, e47892. [Google Scholar] [CrossRef] [PubMed]
- Picca, A.; Guerra, F.; Calvani, R.; Bucci, C.; Lo Monaco, M.R.; Bentivoglio, A.R.; Coelho-Júnior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial Dysfunction and Aging: Insights from the Analysis of Extracellular Vesicles. Int. J. Mol. Sci. 2019, 20, 805. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhang, S.; Wu, J.; Sun, X.; Shen, Z.; Dong, J.; Huang, J. Promotion of mitochondrial biogenesis via activation of AMPK-PGC1ɑ signaling pathway by Ginger (Zingiber officinale Roscoe) extract, and its major active component 6-Gingerol. J. Food Sci. 2019, 84, 2101–2111. [Google Scholar] [CrossRef] [PubMed]
- Ballester, P.; Cerdá, B.; Arcusa, R.; Marhuenda, J.; Yamedjeu, K.; Zafrilla, P. Effect of ginger on inflammatory diseases. Molecules 2022, 27, 7223. [Google Scholar] [CrossRef]
- Maiuolo, J.; Oppedisano, F.; Gratteri, S.; Muscoli, C.; Mollace, V. Regulation of uric acid metabolism and excretion. Int. J. Cardiol. 2016, 213, 8–14. [Google Scholar] [CrossRef]
- Nahas, P.C.; Rossato, L.T.; de Branco, F.M.S.; Azeredo, C.M.; Rinaldi, A.E.M.; de Oliveira, E.P. Serum uric acid is positively associated with muscle strength in older men and women: Findings from NHANES 1999-2002. Clin. Nutr. 2021, 40, 4386–4393. [Google Scholar] [CrossRef]
- Nahas, P.C.; de Branco, F.M.S.; Azeredo, C.M.; Rinaldi, A.E.M.; de Oliveira, E.P. Serum uric acid is not associated with appendicular muscle mass index in young and middle-aged adults: Results from NHANES 2011-2012. Clin. Nutr. ESPEN 2022, 52, 262–269. [Google Scholar] [CrossRef]
- Drey, M. Sarcopenia—Pathophysiology and clinical relevance. Wien. Med. Wochenschr. 2011, 161, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Drory, V.E. Influence of serum uric acid levels on prognosis and survival in amyotrophic lateral sclerosis: A meta-analysis. J. Neurol. 2014, 261, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, Y.S.; Park, S.H.; Kang, K.Y. High serum uric acid level is associated with greater handgrip strength in the aged population. Arthritis Res. Ther. 2019, 21, 73. [Google Scholar] [CrossRef]
- Iwama, M.; Kondo, Y.; Shimokado, K.; Maruyama, N.; Ishigami, A. Uric acid levels in tissues and plasma of mice during aging. Biol. Pharm. Bull. 2012, 35, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Batta, A. Uric Acid is a Significant Antioxidant. Eur. J. Pharm. Med. Res. 2016, 2016, 242–248. [Google Scholar]
- Nile, S.H.; Park, S.W. Chromatographic analysis, antioxidant, anti-inflammatory, and xanthine oxidase inhibitory activities of ginger extracts and its reference compounds. Ind. Crops Prod. 2015, 70, 238–244. [Google Scholar] [CrossRef]
- Packer, M. Uric acid is a biomarker of oxidative stress in the failing heart: Lessons learned from trials with allopurinol and SGLT2 inhibitors. J. Card. Fail. 2020, 26, 977–984. [Google Scholar] [CrossRef]
- Ostojic, S.M.; Premusz, V.; Nagy, D.; Acs, P. Guanidinoacetic acid as a novel food for skeletal muscle health. J. Funct. Foods 2020, 73, 104129. [Google Scholar] [CrossRef]
- Wu, H.; Tang, D.; Yun, M.; Liu, H.; Huang, S.; Yun, C.; Hocher, B.; Zhang, X.; Liu, F.; Yin, L. Metabolic dysfunctions of intestinal fatty acids and tryptophan reveal immuno-inflammatory response activation in IgA nephropathy. Front. Med. 2022, 9, 22. [Google Scholar] [CrossRef]
- Arnold, R.; Issar, T.; Krishnan, A.V.; Pussell, B.A. Neurological complications in chronic kidney disease. JRSM Cardiovasc. Dis. 2016, 5, 2048004016677687. [Google Scholar] [CrossRef]
- Wishart, D.S. Metabolomics for investigating physiological and pathophysiological processes. Physiol. Rev. 2019, 99, 1819–1875. [Google Scholar] [CrossRef] [PubMed]
- Tessari, P. Leucine Transamination Is Lower in Middle-Aged Compared with Younger Adults. J. Nutr. 2017, 147, 2025–2030. [Google Scholar] [CrossRef] [PubMed]
- Opazo, R.; Angel, B.; Márquez, C.; Lera, L.; Cardoso Dos Santos, G.R.; Monnerat, G.; Albala, C. Sarcopenic metabolomic profile reflected a sarcopenic phenotype associated with amino acid and essential fatty acid changes. Metabolomics 2021, 17, 83. [Google Scholar] [CrossRef]
- Lin, C.-C.; Shih, M.-H.; Chen, C.-D.; Yeh, S.-L. Effects of adequate dietary protein with whey protein, leucine, and vitamin D supplementation on sarcopenia in older adults: An open-label, parallel-group study. Clin. Nutr. 2021, 40, 1323–1329. [Google Scholar] [CrossRef] [PubMed]
- Chae, M.; Park, H.S.; Park, K. Association between dietary branched-chain amino acid intake and skeletal muscle mass index among Korean adults: Interaction with obesity. Nutr. Res. Pract. 2020, 15, 203–212. [Google Scholar] [CrossRef] [PubMed]
- Yoshizawa, F.; Mochizuki, S.; Sugahara, K. Differential dose response of mTOR signaling to oral administration of leucine in skeletal muscle and liver of rats. Biosci. Biotechnol. Biochem. 2013, 77, 839–842. [Google Scholar] [CrossRef]
- Duan, Y.; Zeng, L.; Li, F.; Wang, W.; Li, Y.; Guo, Q.; Ji, Y.; Tan, B.e.; Yin, Y. Effect of branched-chain amino acid ratio on the proliferation, differentiation, and expression levels of key regulators involved in protein metabolism of myocytes. Nutrition 2017, 36, 8–16. [Google Scholar] [CrossRef]
- Petrocelli, J.J.; Mahmassani, Z.S.; Fix, D.K.; Montgomery, J.A.; Reidy, P.T.; McKenzie, A.I.; de Hart, N.M.; Ferrara, P.J.; Kelley, J.J.; Eshima, H. Metformin and leucine increase satellite cells and collagen remodeling during disuse and recovery in aged muscle. FASEB J. 2021, 35, e21862. [Google Scholar] [CrossRef]
- Parkhitko, A.A.; Jouandin, P.; Mohr, S.E.; Perrimon, N. Methionine metabolism and methyltransferases in the regulation of aging and lifespan extension across species. Aging Cell 2019, 18, e13034. [Google Scholar] [CrossRef]
- Tripodi, F.; Castoldi, A.; Nicastro, R.; Reghellin, V.; Lombardi, L.; Airoldi, C.; Falletta, E.; Maffioli, E.; Scarcia, P.; Palmieri, L.; et al. Methionine supplementation stimulates mitochondrial respiration. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 1901–1913. [Google Scholar] [CrossRef]
- Perrone, C.E.; Mattocks, D.A.L.; Jarvis-Morar, M.; Plummer, J.D.; Orentreich, N. Methionine restriction effects on mitochondrial biogenesis and aerobic capacity in white adipose tissue, liver, and skeletal muscle of F344 rats. Metabolism 2010, 59, 1000–1011. [Google Scholar] [CrossRef]
- Kozieł, R.; Ruckenstuhl, C.; Albertini, E.; Neuhaus, M.; Netzberger, C.; Bust, M.; Madeo, F.; Wiesner, R.J.; Jansen-Dürr, P. Methionine restriction slows down senescence in human diploid fibroblasts. Aging Cell 2014, 13, 1038–1048. [Google Scholar] [CrossRef]
- Sanchez-Roman, I.; Barja, G. Regulation of longevity and oxidative stress by nutritional interventions: Role of methionine restriction. Exp. Gerontol. 2013, 48, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Guan, K.-L. mTOR as a central hub of nutrient signalling and cell growth. Nat. Cell Biol. 2019, 21, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Ano, Y.; Ozawa, M.; Kutsukake, T.; Sugiyama, S.; Uchida, K.; Yoshida, A.; Nakayama, H. Preventive effects of a fermented dairy product against Alzheimer’s disease and identification of a novel oleamide with enhanced microglial phagocytosis and anti-inflammatory activity. PLoS ONE 2015, 10, e0118512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Bhattacharyya, S.; Hickner, R.C.; Light, A.R.; Lambert, C.J.; Gale, B.K.; Fiehn, O.; Adams, S.H. Skeletal muscle interstitial fluid metabolomics at rest and associated with an exercise bout: Application in rats and humans. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E43–E53. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Watanabe, N.; Kitakaze, T.; Sugimoto, K.; Izawa, T.; Kai, K.; Harada, N.; Yamaji, R. Oleamide rescues tibialis anterior muscle atrophy of mice housed in small cages. Br. J. Nutr. 2021, 126, 481–491. [Google Scholar] [CrossRef]
- Ueland, P.M.; McCann, A.; Midttun, Ø.; Ulvik, A. Inflammation, vitamin B6 and related pathways. Mol. Aspects Med. 2017, 53, 10–27. [Google Scholar] [CrossRef]
- Kumar, A.; Kumar, Y.; Sevak, J.K.; Kumar, S.; Kumar, N.; Gopinath, S.D. Metabolomic analysis of primary human skeletal muscle cells during myogenic progression. Sci. Rep. 2020, 10, 11824. [Google Scholar] [CrossRef]
- Suidasari, S.; Uragami, S.; Yanaka, N.; Kato, N. Dietary vitamin B6 modulates the gene expression of myokines, Nrf2-related factors, myogenin and HSP60 in the skeletal muscle of rats. Exp. Ther. Med. 2017, 14, 3239–3246. [Google Scholar] [CrossRef]
- Kumrungsee, T.; Onishi, K.; Komaru, T.; Yanaka, N.; Kato, N. Vitamin B6 Regulates Muscle Satellite Cell Function: A Novel Possible Role of Vitamin B6 in Muscle Regeneration. Curr. Dev. Nutr. 2020, 4, 4141818. [Google Scholar] [CrossRef]
- Calderón-Ospina, C.A.; Nava-Mesa, M.O. B Vitamins in the nervous system: Current knowledge of the biochemical modes of action and synergies of thiamine, pyridoxine, and cobalamin. CNS Neurosci. Ther. 2020, 26, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Effiong, U. Isolated Pyridoxine Deficiency Presenting as Muscle Spasms in a Patient With Type 2 Diabetes: A Case Report and Literature Review. Am. J. Med. Sci. 2021, 361, 791–794. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, J.; Baur, J.A.; Imai, S.-i. NAD+ intermediates: The biology and therapeutic potential of NMN and NR. Cell Metab. 2018, 27, 513–528. [Google Scholar] [CrossRef]
- Wohlrab, J.; Kreft, D. Niacinamide—Mechanisms of action and its topical use in dermatology. Skin. Pharmacol. Physiol. 2014, 27, 311–315. [Google Scholar] [CrossRef]
- Villeda-González, J.D.; Gómez-Olivares, J.L.; Baiza-Gutman, L.A.; Manuel-Apolinar, L.; Damasio-Santana, L.; Millán-Pacheco, C.; Ángeles-Mejía, S.; Cortés-Ginez, M.C.; Cruz-López, M.; Vidal-Moreno, C.J.; et al. Nicotinamide reduces inflammation and oxidative stress via the cholinergic system in fructose-induced metabolic syndrome in rats. Life Sci. 2020, 250, 117585. [Google Scholar] [CrossRef]
- Bierman, J.C.; Laughlin, T.; Tamura, M.; Hulette, B.C.; Mack, C.E.; Sherrill, J.D.; Tan, C.Y.R.; Morenc, M.; Bellanger, S.; Oblong, J.E. Niacinamide mitigates SASP-related inflammation induced by environmental stressors in human epidermal keratinocytes and skin. Int. J. Cosmet. Sci. 2020, 42, 501–511. [Google Scholar] [CrossRef]
- Sahin, K.; Kucuk, O.; Orhan, C.; Tuzcu, M.; Durmus, A.S.; Ozercan, I.H.; Sahin, N.; Juturu, V. Niacinamide and undenatured type II collagen modulates the inflammatory response in rats with monoiodoacetate-induced osteoarthritis. Sci. Rep. 2021, 11, 14724. [Google Scholar] [CrossRef]
- Unuofin, J.O.; Masuku, N.P.; Paimo, O.K.; Lebelo, S.L. Ginger from farmyard to town: Nutritional and pharmacological applications. Front. Pharmacol. 2021, 12, 779352. [Google Scholar] [CrossRef]
- Shieh, P.-C.; Chen, Y.-O.; Kuo, D.-H.; Chen, F.-A.; Tsai, M.-L.; Chang, I.-S.; Wu, H.; Sang, S.; Ho, C.-T.; Pan, M.-H. Induction of Apoptosis by (8)-Shogaol via Reactive Oxygen Species Generation, Glutathione Depletion, and Caspase Activation in Human Leukemia Cells. J. Agric. Food Chem. 2010, 58, 3847–3854. [Google Scholar] [CrossRef]
- Jo, S.; Samarpita, S.; Lee, J.S.; Lee, Y.J.; Son, J.E.; Jeong, M.; Kim, J.H.; Hong, S.; Yoo, S.-A.; Kim, W.-U.; et al. 8-Shogaol inhibits rheumatoid arthritis through targeting TAK1. Pharmacol. Res. 2022, 178, 106176. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-R.; Noh, E.-M.; Kim, S.-Y. Anti-inflammatory effect and signaling mechanism of 8-shogaol and 10-shogaol in a dextran sodium sulfate-induced colitis mouse model. Heliyon 2023, 9, e12778. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Zhang, Y.; Li, W.; Chen, K.; Xiang, M.; Ma, H. Glutamine metabolism: From proliferating cells to cardiomyocytes. Metabolism 2021, 121, 154778. [Google Scholar] [CrossRef]
- Yoo, H.C.; Yu, Y.C.; Sung, Y.; Han, J.M. Glutamine reliance in cell metabolism. Exp. Mol. Med. 2020, 52, 1496–1516. [Google Scholar] [CrossRef]
- van de Vyver, M.; Myburgh, K.H. Cytokine and satellite cell responses to muscle damage: Interpretation and possible confounding factors in human studies. J. Muscle Res. Cell Motil. 2012, 33, 177–185. [Google Scholar] [CrossRef] [PubMed]
- Girven, M.; Dugdale, H.F.; Owens, D.J.; Hughes, D.C.; Stewart, C.E.; Sharples, A.P. l-glutamine Improves Skeletal Muscle Cell Differentiation and Prevents Myotube Atrophy After Cytokine (TNF-α) Stress Via Reduced p38 MAPK Signal Transduction. J. Cell Physiol. 2016, 231, 2720–2732. [Google Scholar] [CrossRef]
- Gnoni, A.; Longo, S.; Gnoni, G.V.; Giudetti, A.M. Carnitine in human muscle bioenergetics: Can carnitine supplementation improve physical exercise? Molecules 2020, 25, 182. [Google Scholar] [CrossRef]
- Fielding, R.; Riede, L.; Lugo, J.P.; Bellamine, A. L-carnitine supplementation in recovery after exercise. Nutrients 2018, 10, 349. [Google Scholar] [CrossRef]
- Ohara, M.; Ogawa, K.; Suda, G.; Kimura, M.; Maehara, O.; Shimazaki, T.; Suzuki, K.; Nakamura, A.; Umemura, M.; Izumi, T. L-carnitine suppresses loss of skeletal muscle mass in patients with liver cirrhosis. Hepatol. Commun. 2018, 2, 910–922. [Google Scholar] [CrossRef]
- Evans, M.; Guthrie, N.; Pezzullo, J.; Sanli, T.; Fielding, R.A.; Bellamine, A. Efficacy of a novel formulation of L-Carnitine, creatine, and leucine on lean body mass and functional muscle strength in healthy older adults: A randomized, double-blind placebo-controlled study. Nutr. Metab. 2017, 14, 7. [Google Scholar] [CrossRef]
- Amorim, J.A.; Coppotelli, G.; Rolo, A.P.; Palmeira, C.M.; Ross, J.M.; Sinclair, D.A. Mitochondrial and metabolic dysfunction in ageing and age-related diseases. Nat. Rev. Endocrinol. 2022, 18, 243–258. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode | Metabolite | ID | MW | RT | FC | |
|---|---|---|---|---|---|---|
| PSC vs. YC | SC vs. YC | |||||
| neg | 4-Oxoproline | METPA0228 | 129.04169 | 0.849 | −2.39 | − |
| neg | 2,2′-Methylenebis (4-methyl-6-tert-butylphenol | mzc1138 | 340.24016 | 12.551 | − | −3.40 |
| neg | Adipic acid | HMDB0000448 | 146.05714 | 2.275 | −21.80 | −17.95 |
| neg | Citric acid | HMDB0000094 | 192.0265 | 0.788 | −1.41 | − |
| neg | Glutamic acid | HMDB0000148/HMDB0003339 | 147.05221 | 0.603 | −1.34 | − |
| neg | Guanosine | HMDB0000133 | 283.09157 | 1.319 | −1.38 | − |
| neg | N-Acetyl-L-aspartic acid | HMDB0000812 | 175.04736 | 0.755 | +1.46 | − |
| neg | Pantothenic acid | HMDB0000210 | 219.11032 | 2.158 | −2.01 | − |
| neg | Pyridoxine | HMDB0000239 | 169.07315 | 0.826 | −1.42 | − |
| neg | Succinic acid | HMDB0000254 | 118.02569 | 0.842 | −1.55 | −1.58 |
| neg | Uric acid | HMDB0000289 | 168.0276 | 0.837 | −1.67 | − |
| neg | Uridine | HMDB0000296 | 244.06954 | 0.927 | −1.22 | − |
| neg | Xanthine | HMDB0000292 | 152.03263 | 0.986 | −2.05 | − |
| pos | 2-Hydroxycinnamic acid/4-Hydroxycinnamic acid | HMDB0002641/HMDB0002035 | 164.04743 | 0.934 | −1.78 | − |
| pos | Acetylcholine | HMDB0000895 | 145.1103 | 0.891 | −1.18 | − |
| pos | Adenine | HMDB0000034 | 135.05467 | 1.185 | +22.80 | − |
| pos | Adenosine | HMDB0000050 | 267.09665 | 1.242 | +32.75 | − |
| pos | Bis(4-ethylbenzylidene) sorbitol | mzc7437 | 414.20382 | 9.219 | −3.93 | −3.52 |
| pos | Carnitine | HMDB0000062 | 161.10519 | 0.729 | +1.27 | +1.40 |
| pos | Choline | HMDB0000097 | 103.10003 | 0.811 | −1.25 | +1.22 |
| pos | Creatinine | HMDB0000562 | 113.05909 | 0.645 | −1.39 | − |
| pos | Glutamine | HMDB0003423/HMDB0000641 | 146.06919 | 0.605 | −1.40 | − |
| pos | Guanidinosuccinic acid | HMDB0003157 | 175.05853 | 0.977 | −1.48 | −1.45 |
| pos | Guanine | HMDB0000132 | 151.04942 | 1.274 | −1.25 | − |
| pos | Hypoxanthine | HMDB0000157 | 136.03853 | 1.254 | −1.32 | − |
| pos | Indoleacrylic acid | HMDB0000734 | 187.06341 | 2.855 | −1.84 | − |
| pos | Inosine | HMDB0000195 | 268.08055 | 1.249 | −1.54 | − |
| pos | Leucine/Isoleucine/Norleucine | HMDB0000687/HMDB0000172 | 131.09468 | 0.98 | −1.69 | −1.31 |
| pos | Methionine | HMDB0000696 | 149.05104 | 0.79 | −2.12 | −1.40 |
| pos | Octadecanamide | HMDB0034146 | 283.28734 | 13.922 | −1.83 | −2.29 |
| pos | Phenylalanine | HMDB0000159/METPA0264 | 165.07898 | 1.481 | −1.86 | − |
| pos | Proline | HMDB0000162/HMDB0003411 | 115.06359 | 0.701 | −1.49 | − |
| pos | Propionyl carnitine | HMDB0000824 | 217.13122 | 1.488 | −1.89 | − |
| pos | Pyridoxine | HMDB0000239 | 169.074 | 0.853 | −1.59 | −1.43 |
| pos | Pyroglutamic acid | HMDB0000267 | 129.04268 | 0.852 | −2.24 | −1.85 |
| pos | Tyrosine | HMDB0000158/HMDB0006050 | 181.07399 | 0.934 | −1.80 | − |
| pos | Uracil | HMDB0000300 | 112.02756 | 0.932 | −1.16 | − |
| pos | Creatine | HMDB0000064 | 131.06955 | 0.655 | − | +1.26 |
| pos | Docosanamide | HMDB0000583 | 339.34976 | 16.095 | − | −2.34 |
| pos | Erucamide | mzc282 | 337.33412 | 15.515 | − | −1.96 |
| pos | NADH | HMDB0001487 | 663.10909 | 0.943 | − | +2.58 |
| pos | Niacinamide | HMDB0001406 | 122.0482 | 0.67 | − | +1.74 |
| pos | PEG n5 | mzc1819 | 238.14147 | 3.537 | − | −2.58 |
| Mode | Metabolite | ID | MW | RT | FC | |
|---|---|---|---|---|---|---|
| YT50 vs. YC | YT200 vs. YC | |||||
| neg | 2,2′-Methylenebis(4-methyl-6-tert-butylphenol) | mzc1138 | 340.24016 | 12.551 | −2.09 | −2.31 |
| neg | 4-Oxoproline | METPA0228 | 129.04169 | 0.849 | −2.02 | − |
| neg | Adipic acid | HMDB0000448 | 146.05714 | 2.275 | −9.07 | −15.15 |
| neg | N-Acetyl-L-aspartic acid | HMDB0000812 | 175.04736 | 0.755 | +1.55 | − |
| neg | Oxidised glutathione | HMDB0003337 | 612.1527 | 0.892 | − | +1.50 |
| neg | Succinic acid | HMDB0000254 | 118.02569 | 0.842 | − | +1.16 |
| neg | Uridine | HMDB0000296 | 244.06954 | 0.927 | − | +1.14 |
| pos | (8)-Shogaol | HMDB0031463 | 276.1723 | 8.295 | +8.21 | +36.1 |
| pos | Bis(4-ethylbenzylidene) sorbitol | mzc7437 | 414.20382 | 9.219 | −2.93 | −2.73 |
| pos | Carnitine | HMDB0000062 | 161.10519 | 0.729 | +1.30 | − |
| pos | Hypoxanthine | HMDB0000157 | 136.03853 | 1.254 | +1.20 | − |
| pos | Valine | HMDB0000883 | 117.07922 | 0.728 | +1.37 | +1.47 |
| pos | Creatine | HMDB0000064 | 131.06955 | 0.655 | − | −1.18 |
| pos | Cytosine | HMDB0000630 | 111.04353 | 0.667 | − | +1.39 |
| pos | Glutamine | HMDB0003423/HMDB0000641 | 146.06919 | 0.605 | − | +1.24 |
| pos | Niacinamide | HMDB0001406 | 122.0482 | 0.67 | − | +2.13 |
| pos | PEG n7 | mzc1821 | 326.19378 | 4.316 | − | +2.04 |
| pos | Thiamine | HMDB0000235 | 264.10438 | 0.77 | − | +1.47 |
| Mode | Metabolite | ID | MW | RT | FC | |
|---|---|---|---|---|---|---|
| PST50 vs. PSC | PST300 vs. PSC | |||||
| neg | Citric acid | HMDB0000094 | 192.0265 | 0.788 | −1.18 | − |
| neg | Pyridoxine | HMDB0000239 | 169.07315 | 0.826 | +1.36 | − |
| neg | 2,2′-Methylenebis(4-methyl-6-tert-butylphenol) | mzc1138 | 340.24016 | 12.551 | − | −1.84 |
| neg | Glutamic acid | HMDB0000148/HMDB0003339 | 147.05221 | 0.603 | − | −1.24 |
| neg | Oxidised glutathione | HMDB0003337 | 612.1527 | 0.892 | − | +1.42 |
| neg | Succinic acid | HMDB0000254 | 118.02569 | 0.842 | − | +1.17 |
| neg | Uric acid | HMDB0000289 | 168.0276 | 0.837 | − | +1.70 |
| neg | Uridine | HMDB0000296 | 244.06954 | 0.927 | − | +1.18 |
| neg | Xanthine | HMDB0000292 | 152.03263 | 0.986 | − | +1.78 |
| pos | (8)-Shogaol | HMDB0031463 | 276.1723 | 8.295 | − | +34.32 |
| pos | Bis(4-ethylbenzylidene) sorbitol | mzc7437 | 414.20382 | 9.219 | +1.48 | − |
| pos | Carnitine | HMDB0000062 | 161.10519 | 0.729 | −1.24 | −1.29 |
| pos | Creatine | HMDB0000064 | 131.06955 | 0.655 | −1.32 | −1.22 |
| pos | Glutathione | HMDB0062697 | 307.08364 | 0.766 | −3.05 | − |
| pos | Leucine/Isoleucine/Norleucine | HMDB0000687/HMDB0000172 | 131.09468 | 0.98 | +1.36 | − |
| pos | Niacinamide | HMDB0001406 | 122.0482 | 0.67 | +1.01 | +1.50 |
| pos | Phenylalanine | HMDB0000159/METPA0264 | 165.07898 | 1.481 | +1.50 | − |
| pos | Pyridoxine | HMDB0000239 | 169.074 | 0.853 | +1.53 | − |
| pos | Thiamine | HMDB0000235 | 264.10438 | 0.77 | +1.40 | − |
| pos | Glutamine | HMDB0003423/HMDB0000641 | 146.06919 | 0.605 | − | +1.25 |
| Mode | Metabolite | ID | MW | RT | FC | |
|---|---|---|---|---|---|---|
| ST100 vs. SC | ST300 vs. SC | |||||
| neg | 2,2′-Methylenebis(4-methyl-6-tert-butylphenol) | mzc1138 | 340.24016 | 12.551 | +3.77 | |
| neg | 4-Oxoproline | METPA0228 | 129.04169 | 0.849 | − | −1.88 |
| neg | N-Acetyl-L-aspartic acid | HMDB0000812 | 175.04736 | 0.755 | − | −1.51 |
| neg | Oxidised glutathione | HMDB0003337 | 612.1527 | 0.892 | − | +1.73 |
| neg | Uric acid | HMDB0000289 | 168.0276 | 0.837 | −2.51 | −2.58 |
| pos | (8)-Shogaol | HMDB0031463 | 276.1723 | 8.295 | +16.70 | +56.13 |
| pos | Carnitine | HMDB0000062 | 161.10519 | 0.729 | −1.23 | −1.33 |
| pos | Creatine | HMDB0000064 | 131.06955 | 0.655 | −1.28 | − |
| pos | Cytosine | HMDB0000630 | 111.04353 | 0.667 | +1.67 | − |
| pos | Glutathione | HMDB0062697 | 307.08364 | 0.766 | −4.09 | − |
| pos | Octadecanamide | HMDB0034146 | 283.28734 | 13.922 | +1.85 | +2.89 |
| pos | Uracil | HMDB0000300 | 112.02756 | 0.932 | +1.21 | +1.20 |
| pos | Acetylcholine | HMDB0000895 | 145.1103 | 0.891 | − | −1.26 |
| pos | Adenine | HMDB0000034 | 135.05467 | 1.185 | − | +36.70 |
| pos | Adenosine | HMDB0000050 | 267.09665 | 1.242 | − | +45.74 |
| pos | Bis(4-ethylbenzylidene) sorbitol | mzc7437 | 414.20382 | 9.219 | − | +1.61 |
| pos | Docosanamide | HMDB0000583 | 339.34976 | 16.095 | − | +3.06 |
| pos | Erucamide | mzc282 | 337.33412 | 15.515 | − | +2.83 |
| pos | Guanine | HMDB0000132 | 151.04942 | 1.274 | − | −1.28 |
| pos | Hypoxanthine | HMDB0000157 | 136.03853 | 1.254 | − | −1.31 |
| pos | Inosine | HMDB0000195 | 268.08055 | 1.249 | − | −1.35 |
| pos | Oxidised glutathione | HMDB0003337 | 612.15161 | 0.9 | − | +1.74 |
| pos | Pyroglutamic acid | HMDB0000267 | 129.04268 | 0.852 | − | −1.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Sahardi, N.F.N.; Jaafar, F.; Tan, J.K.; Mad Nordin, M.F.; Makpol, S. Elucidating the Pharmacological Properties of Zingiber officinale Roscoe (Ginger) on Muscle Ageing by Untargeted Metabolomic Profiling of Human Myoblasts. Nutrients 2023, 15, 4520. https://doi.org/10.3390/nu15214520
Mohd Sahardi NFN, Jaafar F, Tan JK, Mad Nordin MF, Makpol S. Elucidating the Pharmacological Properties of Zingiber officinale Roscoe (Ginger) on Muscle Ageing by Untargeted Metabolomic Profiling of Human Myoblasts. Nutrients. 2023; 15(21):4520. https://doi.org/10.3390/nu15214520
Chicago/Turabian StyleMohd Sahardi, Nur Fatin Nabilah, Faizul Jaafar, Jen Kit Tan, Mariam Firdhaus Mad Nordin, and Suzana Makpol. 2023. "Elucidating the Pharmacological Properties of Zingiber officinale Roscoe (Ginger) on Muscle Ageing by Untargeted Metabolomic Profiling of Human Myoblasts" Nutrients 15, no. 21: 4520. https://doi.org/10.3390/nu15214520
APA StyleMohd Sahardi, N. F. N., Jaafar, F., Tan, J. K., Mad Nordin, M. F., & Makpol, S. (2023). Elucidating the Pharmacological Properties of Zingiber officinale Roscoe (Ginger) on Muscle Ageing by Untargeted Metabolomic Profiling of Human Myoblasts. Nutrients, 15(21), 4520. https://doi.org/10.3390/nu15214520