
Effects of Caloric Restriction on Spatial Object Recognition Memory, Hippocampal Neuron Loss and Neuroinflammation in Aged Rats

 ,
, 
 ,
, 
Abstract
1. Introduction
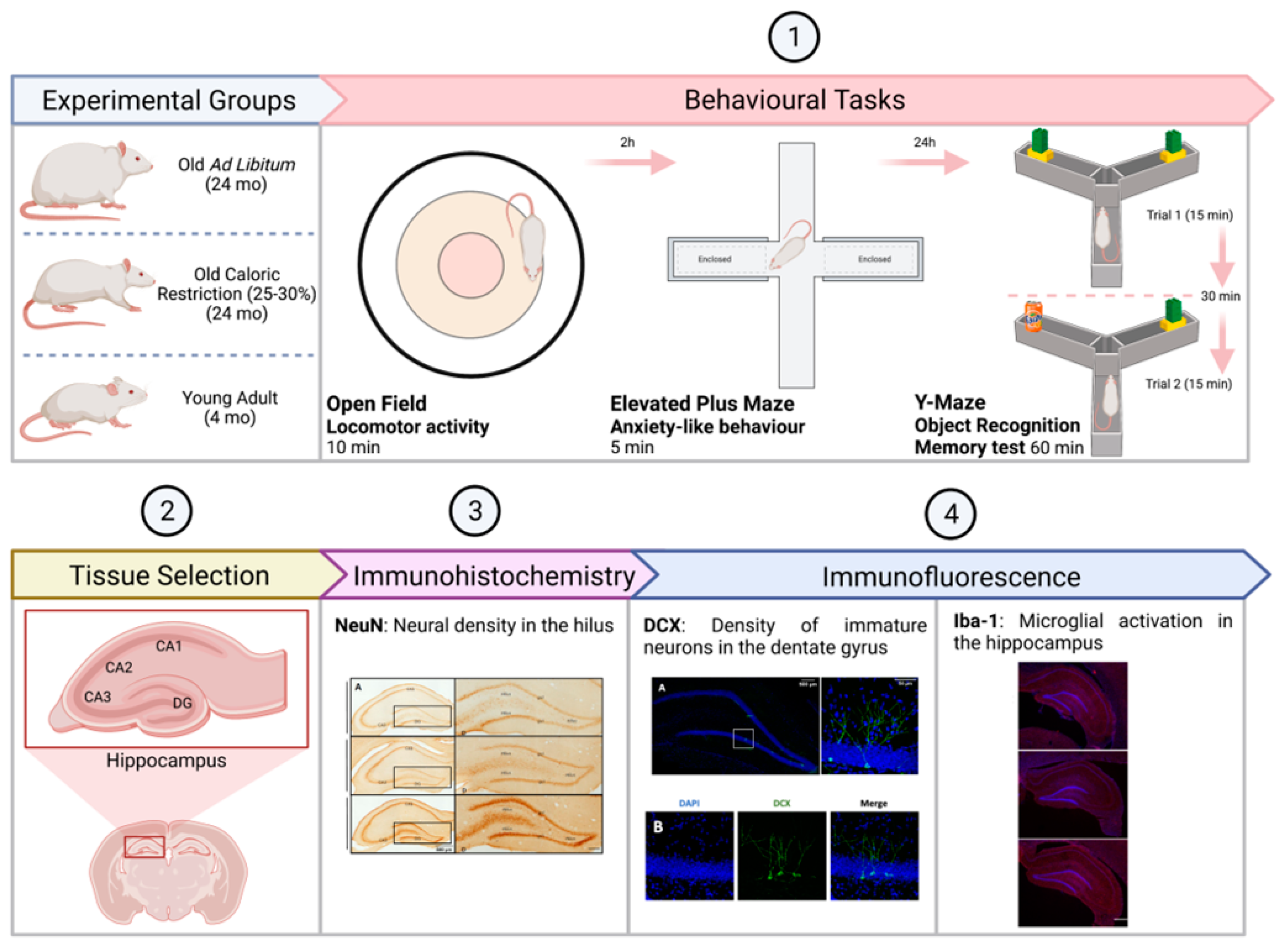
2. Materials and Methods
2.1. Subjects
2.2. Measures
2.2.1. Behavioural Tasks
2.2.2. Protocols for Immunolabeling
2.3. Statistical Analysis
3. Results
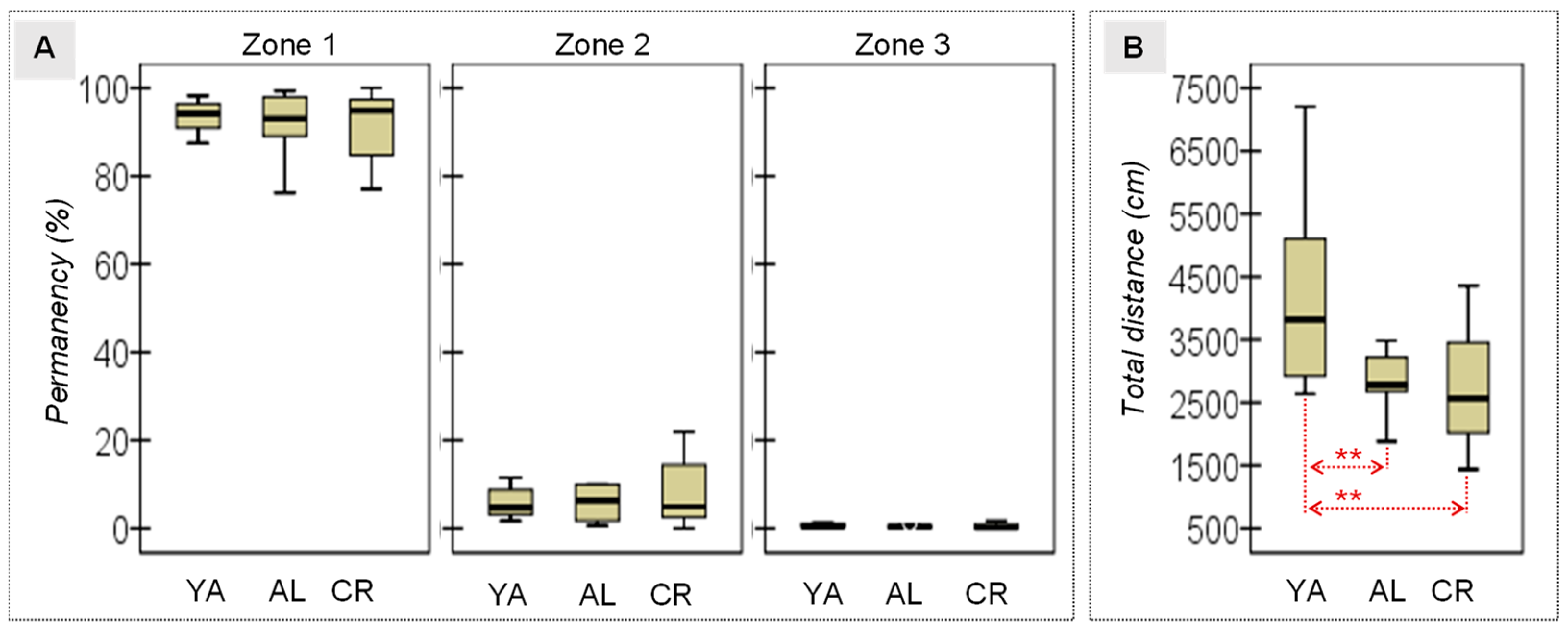
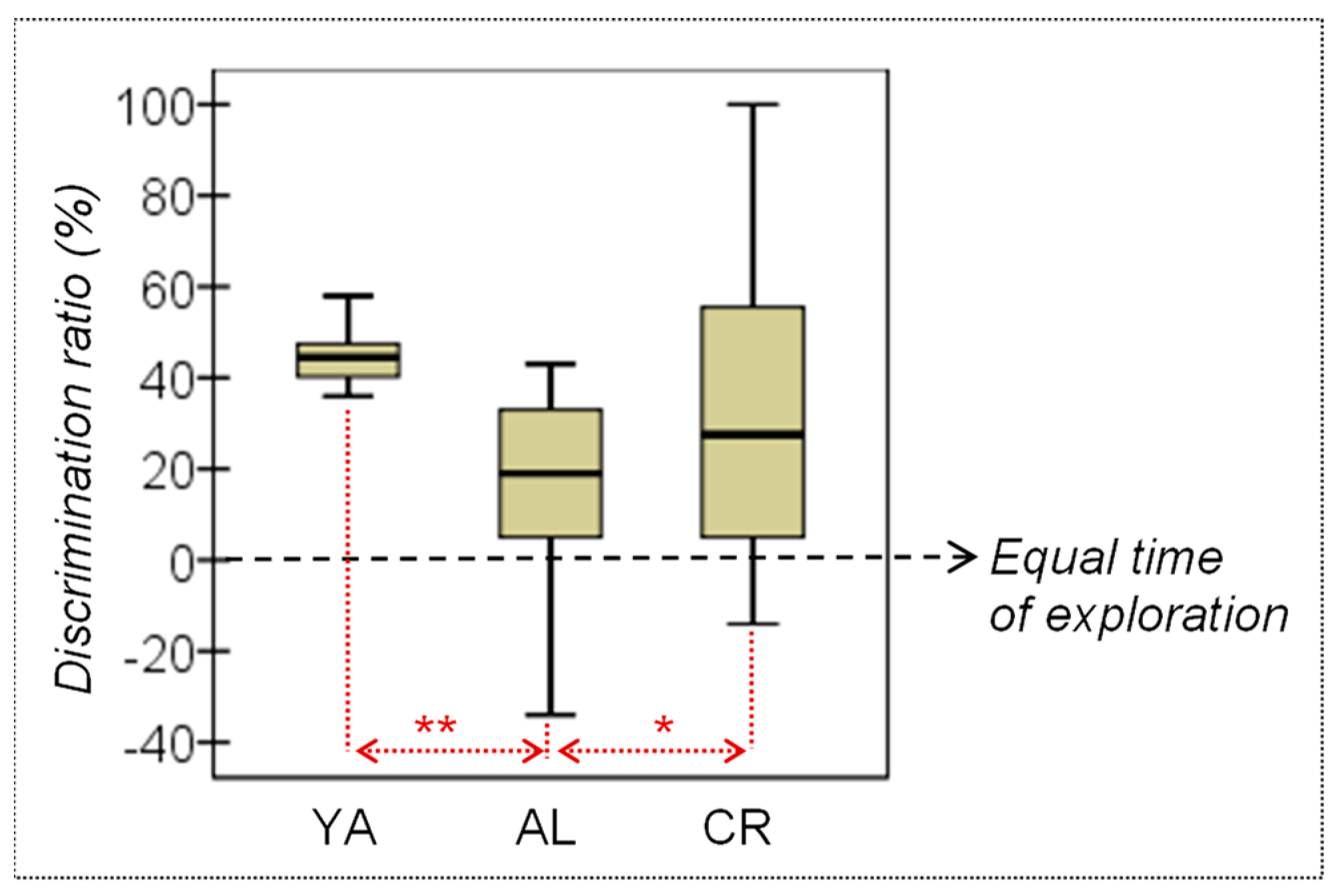
3.1. Behavioral Measures
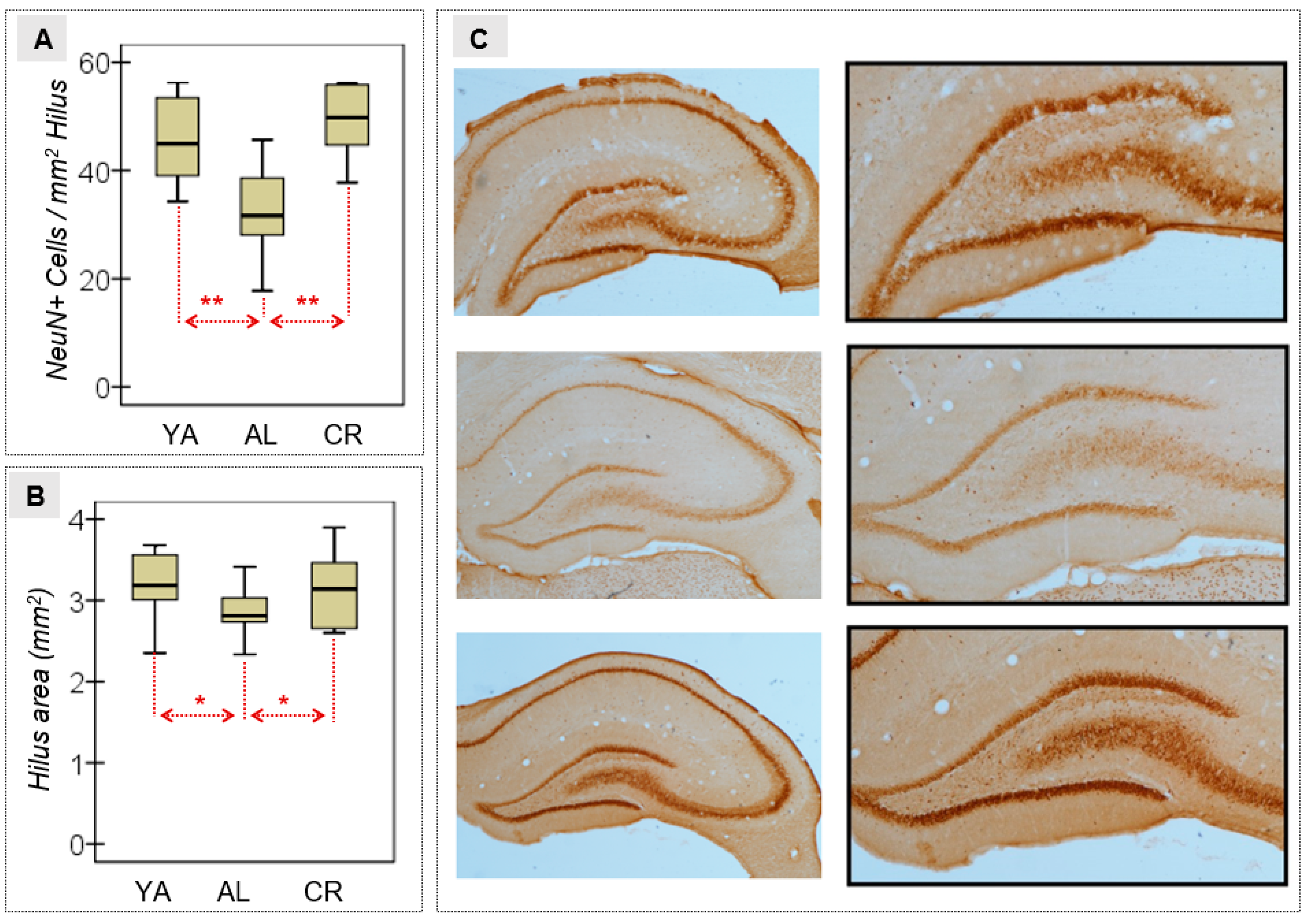
3.2. Neural Density in the Hilus of the DG
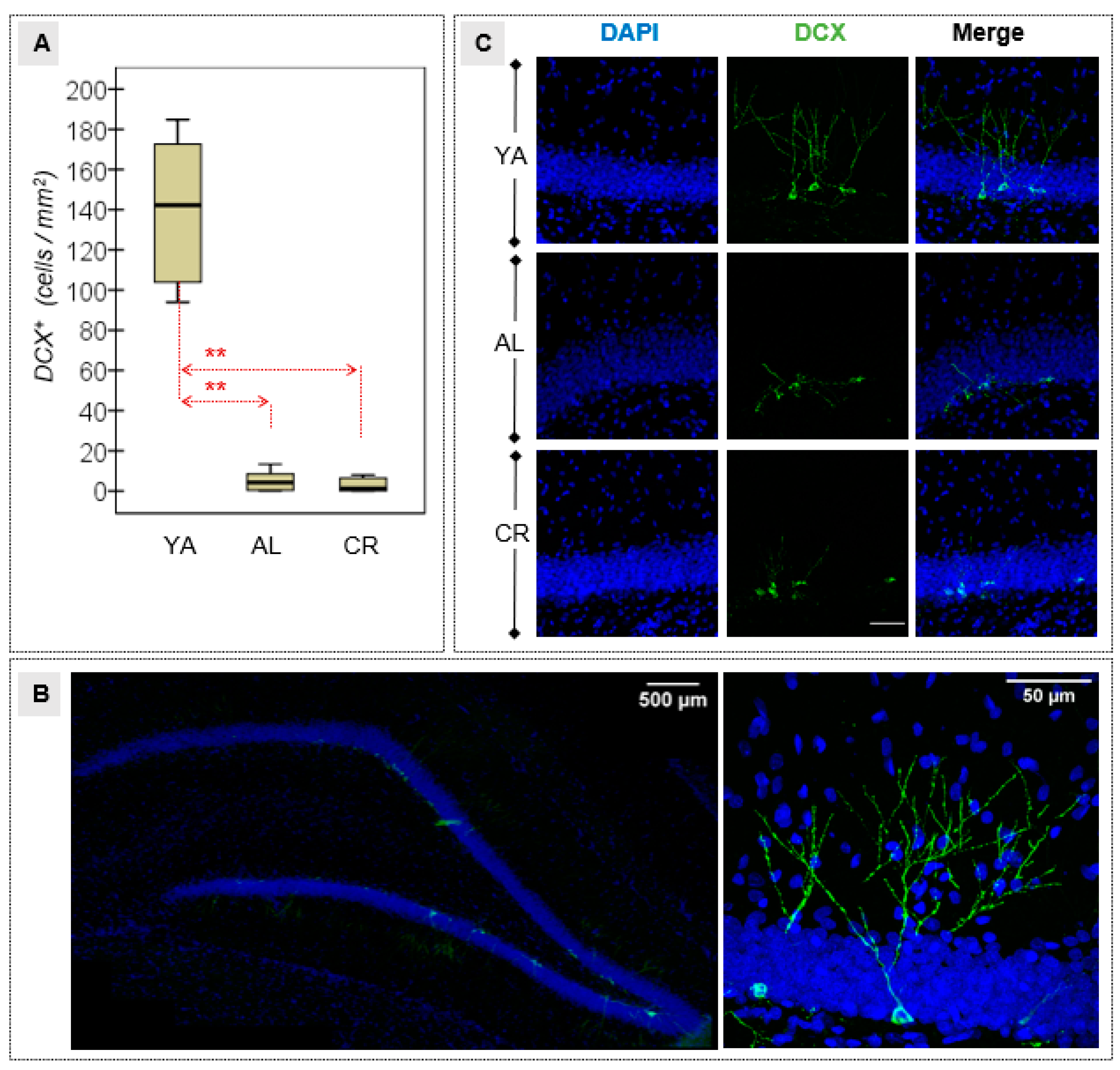
3.3. Density of Immature Neurons in the DG
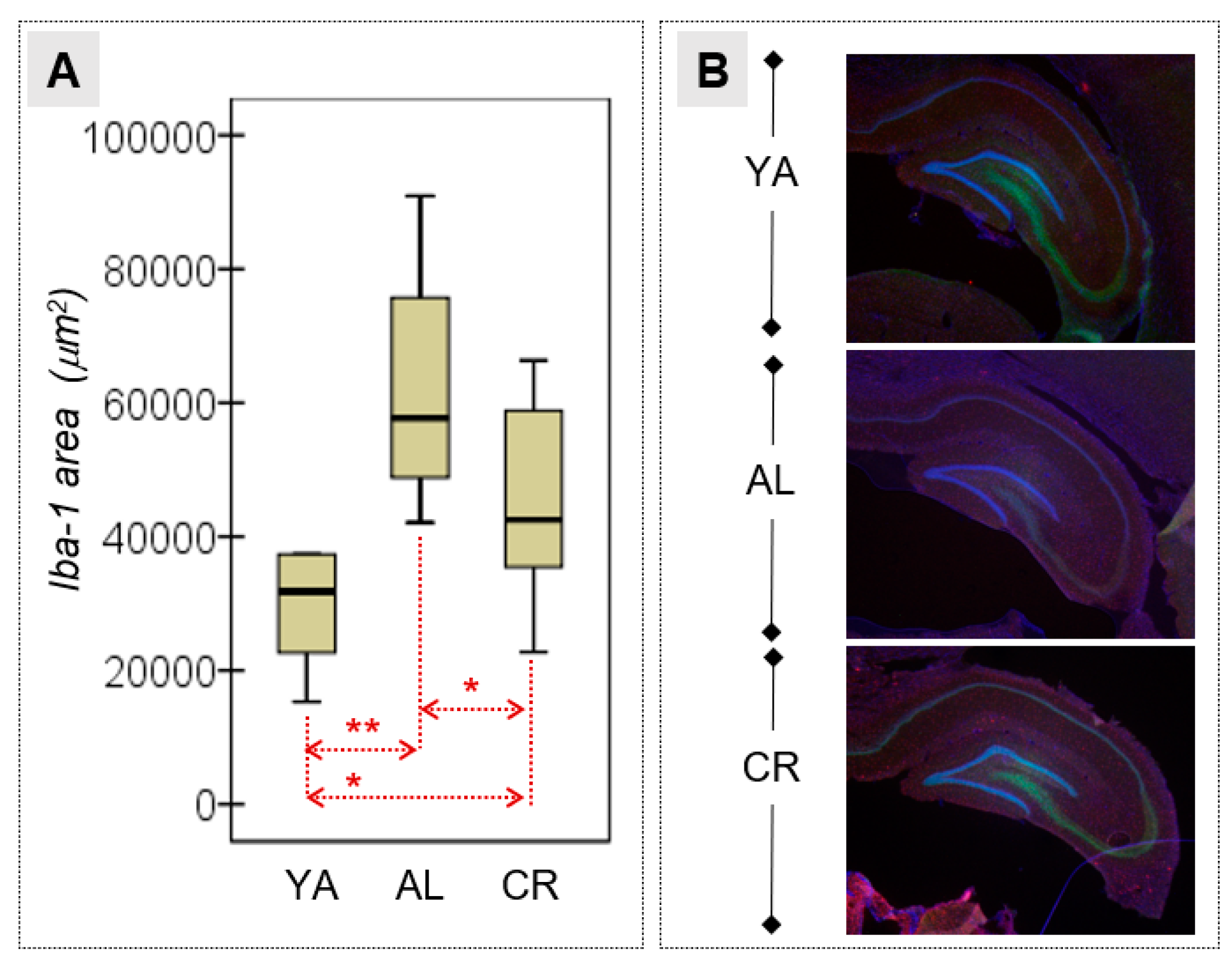
3.4. Microglial Activation in the HPC
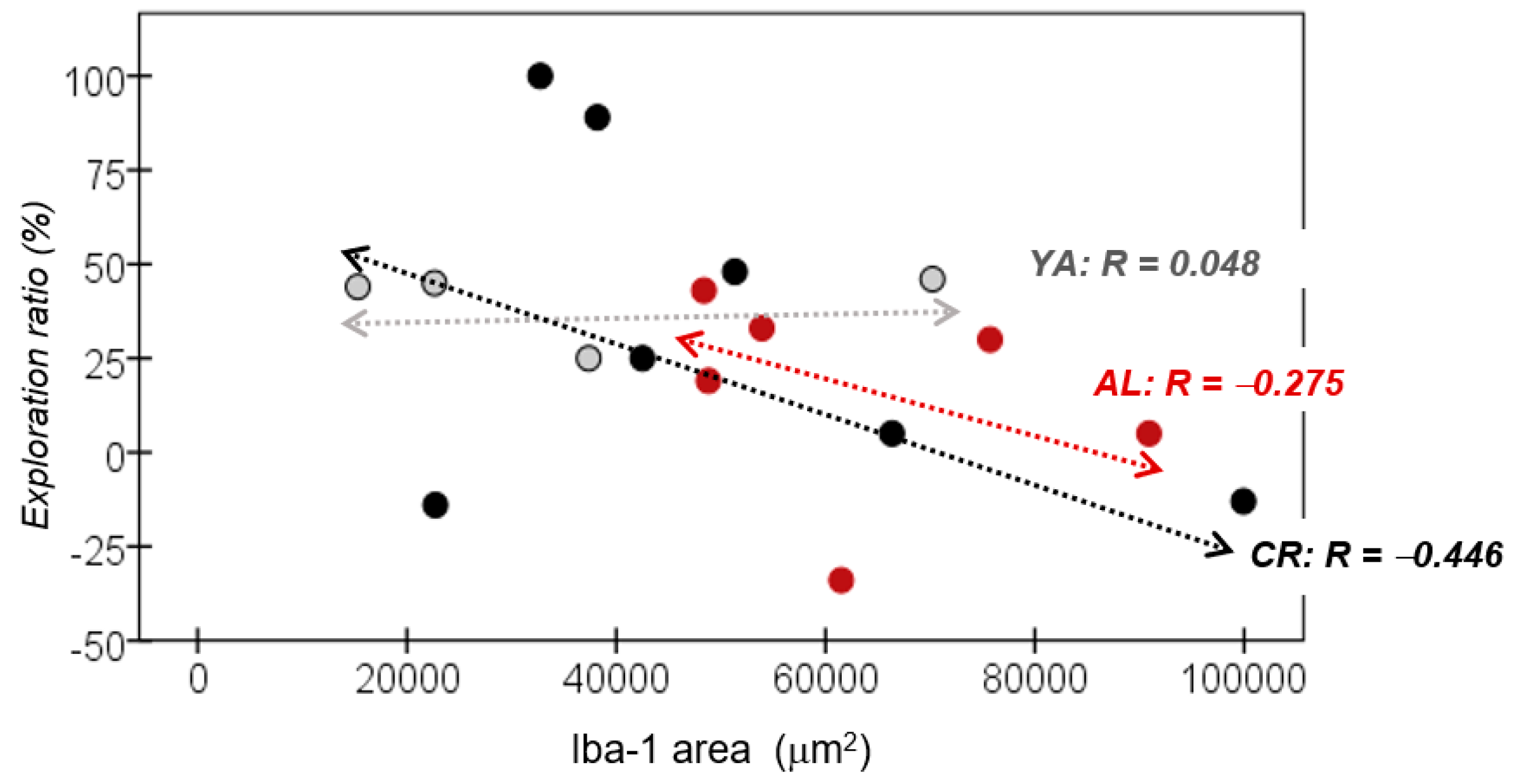
3.5. Correlation between Recognition Memory and Iba-1 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Bettio, L.E.B.; Rajendran, L.; Gil-Mohapel, J. The Effects of Aging in the Hippocampus and Cognitive Decline. Neurosci. Biobehav. Rev. 2017, 79, 66–86. [Google Scholar] [CrossRef]
- Dahan, L.; Rampon, C.; Florian, C. Age-Related Memory Decline, Dysfunction of the Hippocampus and Therapeutic Opportunities. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 102, 109943. [Google Scholar] [CrossRef] [PubMed]
- Rojic-Becker, D.; Portero-Tresserra, M.; Martí-Nicolovius, M.; Vale-Martínez, A.; Guillazo-Blanch, G. Caloric Restriction Modulates the Monoaminergic and Glutamatergic Systems in the Hippocampus, and Attenuates Age-Dependent Spatial Memory Decline. Neurobiol. Learn. Mem. 2019, 166, 107107. [Google Scholar] [CrossRef] [PubMed]
- Lana, D.; Ugolini, F.; Nosi, D.; Wenk, G.L.; Giovannini, M.G. The Emerging Role of the Interplay Among Astrocytes, Microglia, and Neurons in the Hippocampus in Health and Disease. Front. Aging Neurosci. 2021, 13, 651973. [Google Scholar] [CrossRef]
- Dugan, B.; Conway, J.; Duggal, N.A. Inflammaging as a Target for Healthy Ageing. Age Ageing 2023, 52, afac328. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, I.; Howard, S.R.; Stone, J.C.; Monfils, M.H.; Tomanek, B.; Brooks, W.M.; Sutherland, R.J. The Aging Hippocampus: A Multi-Level Analysis in the Rat. Neuroscience 2006, 139, 1173–1185. [Google Scholar] [CrossRef]
- Apple, D.M.; Solano-Fonseca, R.; Kokovay, E. Neurogenesis in the Aging Brain. Biochem. Pharmacol. 2017, 141, 77–85. [Google Scholar] [CrossRef]
- Portero-Tresserra, M.; Martí-Nicolovius, M.; Tarrés-Gatius, M.; Candalija, A.; Guillazo-Blanch, G.; Vale-Martínez, A. Intra-Hippocampal D-Cycloserine Rescues Decreased Social Memory, Spatial Learning Reversal, and Synaptophysin Levels in Aged Rats. Psychopharmacology 2018, 235, 1463–1477. [Google Scholar] [CrossRef]
- Baptista, P.; Andrade, J.P. Adult Hippocampal Neurogenesis: Regulation and Possible Functional and Clinical Correlates. Front. Neuroanat. 2018, 12, 44. [Google Scholar] [CrossRef]
- Yang, T.T.; Lo, C.P.; Tsai, P.S.; Wu, S.Y.; Wang, T.F.; Chen, Y.W.; Jiang-Shieh, Y.F.; Kuo, Y.M. Aging and Exercise Affect Hippocampal Neurogenesis via Different Mechanisms. PLoS ONE 2015, 10, e0132152. [Google Scholar] [CrossRef] [PubMed]
- Altman, J. Are New Neurons Formed in the Brains of Adult Mammals? Science 1962, 135, 1127–1128. [Google Scholar] [CrossRef] [PubMed]
- Gould, E.; Reeves, A.J.; Graziano, M.S.A.; Gross, C.G. Neurogenesis in the Neocortex of Adult Primates. Science 1999, 286, 548–552. [Google Scholar] [CrossRef] [PubMed]
- Kelly, P.H.; Bondolfi, L.; Hunziker, D.; Schlecht, H.P.; Carver, K.; Maguire, E.; Abramowski, D.; Wiederhold, K.H.; Sturchler-Pierrat, C.; Jucker, M.; et al. Progressive Age-Related Impairment of Cognitive Behavior in APP23 Transgenic Mice. Neurobiol. Aging 2003, 24, 365–378. [Google Scholar] [CrossRef]
- Encinas, J.M.; Sierra, A.; Valcárcel-Martín, R.; Martín-Suárez, S. A Developmental Perspective on Adult Hippocampal Neurogenesis. Int. J. Dev. Neurosci. 2013, 31, 640–645. [Google Scholar] [CrossRef]
- Barcia, C.; Ros, C.M.; Annese, V.; de Sauvage, M.A.C.; Ros-Bernal, F.; Gómez, A.; Yuste, J.E.; Campuzano, C.M.; de Pablos, V.; Fernandez-Villalba, E.; et al. ROCK/Cdc42-Mediated Microglial Motility and Gliapse Formation Lead to Phagocytosis of Degenerating Dopaminergic Neurons in Vivo. Sci. Rep. 2012, 2, 809. [Google Scholar] [CrossRef]
- Nayak, D.; Roth, T.L.; McGavern, D.B. Microglia Development and Function. Annu. Rev. Immunol. 2014, 32, 367–402. [Google Scholar] [CrossRef]
- Wang, S.; Liu, J.; Tian, L.; Chen, L.; Wang, J.; Tang, Q.; Zhang, F.; Zhou, Z. Increased Insular Cortical Thickness Associated with Symptom Severity in Male Youths with Internet Gaming Disorder: A Surface-Based Morphometric Study. Front. Psychiatry 2018, 9, 99. [Google Scholar] [CrossRef]
- Yegla, B.; Foster, T.C.; Kumar, A. Behavior Model for Assessing Decline in Executive Function During Aging and Neurodegenerative Diseases. Methods Mol. Biol. 2019, 2011, 441–449. [Google Scholar] [CrossRef]
- Seib, D.R.M.; Martin-Villalba, A. Neurogenesis in the Normal Ageing Hippocampus: A Mini-Review. Gerontology 2015, 61, 327–335. [Google Scholar] [CrossRef]
- Kozareva, D.A.; Cryan, J.F.; Nolan, Y.M. Born This Way: Hippocampal Neurogenesis across the Lifespan. Aging Cell 2019, 18, e13007. [Google Scholar] [CrossRef]
- Spencer, N.G.; Lovell, D.P.; Elderfield, K.; Austen, B.; Howe, F.A. Can MRI T1 Be Used to Detect Early Changes in 5xFAD Alzheimer’s Mouse Brain? MAGMA 2017, 30, 153–163. [Google Scholar] [CrossRef]
- Poulose, S.M.; Miller, M.G.; Scott, T.; Shukitt-Hale, B. Nutritional Factors Affecting Adult Neurogenesis and Cognitive Function. Adv. Nutr. 2017, 8, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Gillespie, Z.E.; Pickering, J.; Eskiw, C.H. Better Living through Chemistry: Caloric Restriction (CR) and CR Mimetics Alter Genome Function to Promote Increased Health and Lifespan. Front. Genet. 2016, 7, 142. [Google Scholar] [CrossRef] [PubMed]
- Hadem, I.K.H.; Majaw, T.; Kharbuli, B.; Sharma, R. Beneficial Effects of Dietary Restriction in Aging Brain. J. Chem. Neuroanat. 2019, 95, 123–133. [Google Scholar] [CrossRef] [PubMed]
- van Cauwenberghe, C.; Vandendriessche, C.; Libert, C.; Vandenbroucke, R.E. Caloric Restriction: Beneficial Effects on Brain Aging and Alzheimer’s Disease. Mamm. Genome 2016, 27, 300–319. [Google Scholar] [CrossRef] [PubMed]
- Portero-Tresserra, M.; Rojic-Becker, D.; Vega-Carbajal, C.; Guillazo-Blanch, G.; Vale-Martínez, A.; Martí-Nicolovius, M. Caloric Restriction Modulates the Monoaminergic System and Metabolic Hormones in Aged Rats. Sci. Rep. 2020, 10, 19299. [Google Scholar] [CrossRef]
- Gallagher, M.; Stocker, A.M.; Koh, M.T. Mindspan: Lessons from Rat Models of Neurocognitive Aging. ILAR J. 2011, 52, 32–40. [Google Scholar] [CrossRef]
- Ingram, D.K.; de Cabo, R. Calorie Restriction in Rodents: Caveats to Consider. Ageing Res. Rev. 2017, 39, 15–28. [Google Scholar] [CrossRef]
- Speakman, J.R.; Mitchell, S.E. Caloric Restriction. Mol. Aspects Med. 2011, 32, 159–221. [Google Scholar] [CrossRef]
- Rojic-Becker, D.; Portero-Tresserra, M.; Martí-Nicolovius, M.; Vale-Martínez, A.; Guillazo-Blanch, G. Effects of Caloric Restriction on Monoaminergic Neurotransmission, Peripheral Hormones, and Olfactory Memory in Aged Rats. Behav. Brain Res. 2021, 409, 113328. [Google Scholar] [CrossRef] [PubMed]
- Pellow, S.; Chopin, P.; File, S.E.; Briley, M. Validation of Open:Closed Arm Entries in an Elevated plus-Maze as a Measure of Anxiety in the Rat. J. Neurosci. Methods 1985, 14, 149–167. [Google Scholar] [CrossRef] [PubMed]
- Walf, A.A.; Frye, C.A. The Use of the Elevated plus Maze as an Assay of Anxiety-Related Behavior in Rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Lissner, L.J.; Wartchow, K.M.; Toniazzo, A.P.; Gonçalves, C.A.; Rodrigues, L. Object Recognition and Morris Water Maze to Detect Cognitive Impairment from Mild Hippocampal Damage in Rats: A Reflection Based on the Literature and Experience. Pharmacol. Biochem. Behav. 2021, 210, 173273. [Google Scholar] [CrossRef] [PubMed]
- Dellu, F.; Mayo, W.; Cherkaoui, J.; Moal, M.L.; Simon, H. A Two-Trial Memory Task with Automated Recording: Study in Young and Aged Rats. Brain Res. 1992, 588, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Conrad, C.D.; Galea, L.A.M.; Kuroda, Y.; McEwen, B.S. Chronic Stress Impairs Rat Spatial Memory on the Y Maze, and This Effect Is Blocked by Tianeptine Pretreatment. Behav. Neurosci. 1996, 110, 1321–1334. [Google Scholar] [CrossRef] [PubMed]
- Preibisch, S.; Saalfeld, S.; Tomancak, P. Globally Optimal Stitching of Tiled 3D Microscopic Image Acquisitions. Bioinformatics 2009, 25, 1463–1465. [Google Scholar] [CrossRef]
- Blanca, M.J.; Alarcón, R.; Arnau, J.; Bono, R.; Bendayan, R. Non-Normal Data: Is ANOVA Still a Valid Option? Psicothema 2017, 29, 552–557. [Google Scholar] [CrossRef]
- Levine, T.R.; Hullet, C.R. Eta Squared, Partial Eta Squared, and Misreporting of Effect Size in Communication Research. Hum. Commun. Res. 2002, 28, 612–625. [Google Scholar] [CrossRef]
- Rosnow, R.L.; Rosenthal, R. Computing Contrasts, Effect Sizes, and Counternulls on Other People’s Published Data: General Procedures for Research Consumers. Psychol. Methods 1996, 1, 331–340. [Google Scholar] [CrossRef]
- Dong, W.; Wang, R.; Ma, L.N.; Xu, B.L.; Zhang, J.S.; Zhao, Z.W.; Wang, Y.L.; Zhang, X. Autophagy Involving Age-Related Cognitive Behavior and Hippocampus Injury Is Modulated by Different Caloric Intake in Mice. Int. J. Clin. Exp. Med. 2015, 8, 11843–11853. [Google Scholar] [PubMed]
- Lueptow, L.M. Novel Object Recognition Test for the Investigation of Learning and Memory in Mice. J. Vis. Exp. 2017, 2017, 55718. [Google Scholar] [CrossRef]
- Warburton, E.C.; Brown, M.W. Neural Circuitry for Rat Recognition Memory. Behav. Brain Res. 2015, 285, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Cohen, S.J.; Stackman, R.W. Assessing Rodent Hippocampal Involvement in the Novel Object Recognition Task. A Review. Behav. Brain Res. 2015, 285, 105–117. [Google Scholar] [CrossRef]
- Richardson, A.E.; Tomasulo, M.M.V. Influence of Acute Stress on Spatial Tasks in Humans. Physiol. Behav. 2011, 103, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Casadesus, G.; Cantuti-Castelvetri, I.; Joseph, J.A. Effect of Age on Object Exploration, Habituation, and Response to Spatial and Nonspatial Change. Behav. Neurosci. 2001, 115, 1059–1064. [Google Scholar] [CrossRef]
- Gardner, R.S.; Gold, P.E.; Korol, D.L. Inactivation of the Striatum in Aged Rats Rescues Their Ability to Learn a Hippocampus-Sensitive Spatial Navigation Task. Neurobiol. Learn. Mem. 2020, 172, 107231. [Google Scholar] [CrossRef]
- Arias-Cavieres, A.; Adasme, T.; Sánchez, G.; Muñoz, P.; Hidalgo, C. Aging Impairs Hippocampal- Dependent Recognition Memory and LTP and Prevents the Associated RyR Up-Regulation. Front. Aging Neurosci. 2017, 9, 111. [Google Scholar] [CrossRef]
- Platano, D.; Fattoretti, P.; Balietti, M.; Bertoni-Freddari, C.; Aicardi, G. Long-Term Visual Object Recognition Memory in Aged Rats. Rejuvenation Res. 2008, 11, 333–339. [Google Scholar] [CrossRef]
- Canatelli-Mallat, M.; Chiavellini, P.; Lehmann, M.; Goya, R.G.; Morel, G.R. Age-Related Loss of Recognition Memory and Its Correlation with Hippocampal and Perirhinal Cortex Changes in Female Sprague Dawley Rats. Behav. Brain Res. 2022, 435, 114026. [Google Scholar] [CrossRef]
- Fraundorf, S.H.; Hourihan, K.L.; Peters, R.A.; Benjamin, A.S. Aging and Recognition Memory: A Meta-Analysis. Psychol. Bull. 2019, 145, 339–371. [Google Scholar] [CrossRef]
- Wahl, D.; Solon-Biet, S.M.; Wang, Q.P.; Wali, J.A.; Pulpitel, T.; Clark, X.; Raubenheimer, D.; Senior, A.M.; Sinclair, D.A.; Cooney, G.J.; et al. Comparing the Effects of Low-Protein and High-Carbohydrate Diets and Caloric Restriction on Brain Aging in Mice. Cell Rep. 2018, 25, 2234–2243.e6. [Google Scholar] [CrossRef] [PubMed]
- Malin, D.H.; Lee, D.R.; Goyarzu, P.; Chang, Y.H.; Ennis, L.J.; Beckett, E.; Shukitt-Hale, B.; Joseph, J.A. Short-Term Blueberry-Enriched Diet Prevents and Reverses Object Recognition Memory Loss in Aging Rats. Nutrition 2011, 27, 338–342. [Google Scholar] [CrossRef] [PubMed]
- Goyarzu, P.; Malin, D.H.; Lau, F.C.; Taglialatela, G.; Moon, W.D.; Jennings, R.; Moy, E.; Moy, D.; Lippold, S.; Shukitt-Hale, B.; et al. Blueberry Supplemented Diet: Effects on Object Recognition Memory and Nuclear Factor-Kappa B Levels in Aged Rats. Nutr. Neurosci. 2004, 7, 75–83. [Google Scholar] [CrossRef]
- Brownlow, M.L.; Joly-Amado, A.; Azam, S.; Elza, M.; Selenica, M.L.; Pappas, C.; Small, B.; Engelman, R.; Gordon, M.N.; Morgan, D. Partial Rescue of Memory Deficits Induced by Calorie Restriction in a Mouse Model of Tau Deposition. Behav. Brain Res. 2014, 271, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Shen, Q.; Dong, S.; Xu, Z.; Tsien, J.Z.; Hu, Y. Calorie Restriction Ameliorates Neurodegenerative Phenotypes in Forebrain-Specific Presenilin-1 and Presenilin-2 Double Knockout Mice. Neurobiol. Aging 2008, 29, 1502–1511. [Google Scholar] [CrossRef]
- Gámiz, F.; Gallo, M. Spontaneous Object Recognition Memory in Aged Rats: Complexity versus Similarity. Learn. Mem. 2012, 19, 444–448. [Google Scholar] [CrossRef]
- de Lima, M.N.M.; Laranja, D.C.; Caldana, F.; Bromberg, E.; Roesler, R.; Schröder, N. Reversal of Age-Related Deficits in Object Recognition Memory in Rats with l-Deprenyl. Exp. Gerontol. 2005, 40, 506–511. [Google Scholar] [CrossRef]
- Bergado, J.A.; Almaguer, W.; Rojas, Y.; Capdevila, V.; Frey, J.U. Spatial and Emotional Memory in Aged Rats: A Behavioral-Statistical Analysis. Neuroscience 2011, 172, 256–269. [Google Scholar] [CrossRef]
- Chao, O.Y.; Nikolaus, S.; Yang, Y.M.; Huston, J.P. Neuronal Circuitry for Recognition Memory of Object and Place in Rodent Models. Neurosci. Biobehav. Rev. 2022, 141, 104855. [Google Scholar] [CrossRef]
- Nadel, L.; Hupbach, A.; Gomez, R.; Newman-Smith, K. Memory Formation, Consolidation and Transformation. Neurosci. Biobehav. Rev. 2012, 36, 1640–1645. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.M. The Role of the Hippocampus in Recognition Memory. Cortex 2017, 93, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Tanimizu, T.; Kono, K.; Kida, S. Brain Networks Activated to Form Object Recognition Memory. Brain Res. Bull. 2018, 141, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Miranda, M.; Bekinschtein, P. Plasticity Mechanisms of Memory Consolidation and Reconsolidation in the Perirhinal Cortex. Neuroscience 2018, 370, 46–61. [Google Scholar] [CrossRef]
- Dong, W.; Wang, R.; Ma, L.N.; Xu, B.L.; Zhang, J.S.; Zhao, Z.W.; Wang, Y.L.; Zhang, X. Influence of Age-Related Learning and Memory Capacity of Mice: Different Effects of a High and Low Caloric Diet. Aging Clin. Exp. Res. 2016, 28, 303–311. [Google Scholar] [CrossRef]
- Kaptan, Z.; Akgün-Dar, K.; Kapucu, A.; Dedeakayoʇullari, H.; Batu, Ş.; Üzüm, G. Long Term Consequences on Spatial Learning-Memory of Low-Calorie Diet during Adolescence in Female Rats; Hippocampal and Prefrontal Cortex BDNF Level, Expression of NeuN and Cell Proliferation in Dentate Gyrus. Brain Res. 2015, 1618, 194–204. [Google Scholar] [CrossRef] [PubMed]
- Bassani, T.B.; Bonato, J.M.; Machado, M.M.F.; Cóppola-Segovia, V.; Moura, E.L.R.; Zanata, S.M.; Oliveira, R.M.M.W.; Vital, M.A.B.F. Decrease in Adult Neurogenesis and Neuroinflammation Are Involved in Spatial Memory Impairment in the Streptozotocin-Induced Model of Sporadic Alzheimer’s Disease in Rats. Mol. Neurobiol. 2018, 55, 4280–4296. [Google Scholar] [CrossRef] [PubMed]
- Gemma, C.; Bachstetter, A.D.; Bickford, P.C. Neuron-Microglia Dialogue and Hippocampal Neurogenesis in the Aged Brain. Aging Dis. 2010, 1, 232. [Google Scholar]
- Sung, P.S.; Lin, P.Y.; Liu, C.H.; Su, H.C.; Tsai, K.J. Neuroinflammation and Neurogenesis in Alzheimer’s Disease and Potential Therapeutic Approaches. Int. J. Mol. Sci. 2020, 21, 701. [Google Scholar] [CrossRef]
- Ledreux, A.; Wang, X.; Schultzberg, M.; Granholm, A.C.; Freeman, L.R. Detrimental Effects of a High Fat/High Cholesterol Diet on Memory and Hippocampal Markers in Aged Rats. Behav. Brain Res. 2016, 312, 294–304. [Google Scholar] [CrossRef]
- Yegla, B.; Foster, T. Effect of Systemic Inflammation on Rat Attentional Function and Neuroinflammation: Possible Protective Role for Food Restriction. Front. Aging Neurosci. 2019, 11, 296. [Google Scholar] [CrossRef] [PubMed]
- Perry, V.H.; Nicoll, J.A.R.; Holmes, C. Microglia in Neurodegenerative Disease. Nat. Rev. Neurol. 2010, 6, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Streit, W.J.; Mrak, R.E.; Sue, W.; Griffin, T. Microglia and Neuroinflammation: A Pathological Perspective. J. Neuroinflamm. 2004, 1, 14. [Google Scholar] [CrossRef]
- Fuentes, E.; Venegas, B.; Muñoz-Arenas, G.; Moran, C.; Vazquez-Roque, R.A.; Flores, G.; Treviño, S.; Diaz, A.; Guevara, J. High-Carbohydrate and Fat Diet Consumption Causes Metabolic Deterioration, Neuronal Damage, and Loss of Recognition Memory in Rats. J. Chem. Neuroanat. 2023, 129, 102237. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Aimone, J.B.; Gage, F.H. New Neurons and New Memories: How Does Adult Hippocampal Neurogenesis Affect Learning and Memory? Nat. Rev. Neurosci. 2010, 11, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Rapp, P.R.; Gallagher, M. Preserved Neuron Number in the Hippocampus of Aged Rats with Spatial Learning Deficits. Proc. Natl. Acad. Sci. USA 1996, 93, 9926–9930. [Google Scholar] [CrossRef]
- Gräff, J.; Tsai, L.H. Histone Acetylation: Molecular Mnemonics on the Chromatin. Nat. Rev. Neurosci. 2013, 14, 97–111. [Google Scholar] [CrossRef]
- Morel, G.R.; Andersen, T.; Pardo, J.; Zuccolilli, G.O.; Cambiaggi, V.L.; Hereñú, C.B.; Goya, R.G. Cognitive Impairment and Morphological Changes in the Dorsal Hippocampus of Very Old Female Rats. Neuroscience 2015, 303, 189–199. [Google Scholar] [CrossRef]
- Park, J.H.; Glass, Z.; Sayed, K.; Michurina, T.V.; Lazutkin, A.; Mineyeva, O.; Velmeshev, D.; Ward, W.F.; Richardson, A.; Enikolopov, G. Calorie Restriction Alleviates the Age-Related Decrease in Neural Progenitor Cell Division in the Aging Brain. Eur. J. Neurosci. 2013, 37, 1987–1993. [Google Scholar] [CrossRef]
- Lee, J.; Seroogy, K.B.; Mattson, M.P. Dietary Restriction Enhances Neurotrophin Expression and Neurogenesis in the Hippocampus of Adult Mice. J. Neurochem. 2002, 80, 539–547. [Google Scholar] [CrossRef]
- Lagace, D.C.; Fischer, S.J.; Eisch, A.J. Gender and Endogenous Levels of Estradiol Do Not Influence Adult Hippocampal Neurogenesis in Mice. Hippocampus 2007, 17, 175–180. [Google Scholar] [CrossRef] [PubMed]
- Apple, D.M.; Mahesula, S.; Fonseca, R.S.; Zhu, C.; Kokovay, E. Calorie Restriction Protects Neural Stem Cells from Age-Related Deficits in the Subventricular Zone. Aging 2019, 11, 115–126. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YA | AL | CR | Factor Group | YA vs. AL | YA vs. CR | AL vs. CR | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | F | df | p | η2 | p | |d| | P | |d| | p | |d| | |
| 1 Behavioral measures | ||||||||||||||||
| OF: Time zone-1 (%) | 92.09 | 7.91 | 89.95 | 10.66 | 89.33 | 13.03 | 0.21 | 2/31 | 0.809 | 0.014 | 0.654 | 0.23 | 0.535 | 0.26 | 0.898 | 0.05 |
| OF: Time zone-2 (%) | 7.09 | 7.01 | 9.58 | 10.46 | 10.21 | 12.55 | 0.31 | 2/31 | 0.739 | 0.020 | 0.585 | 0.28 | 0.461 | 0.31 | 0.891 | 0.05 |
| OF: Time zone-3 (%) | 0.83 | 1.03 | 0.47 | 0.43 | 0.46 | 0.53 | 0.92 | 2/31 | 0.410 | 0.058 | 0.285 | 0.45 | 0.232 | 0.45 | 0.967 | 0.03 |
| OF: Distance (cm) | 4224.7 | 1557.4 | 2965.6 | 711.9 | 2726.9 | 916.8 | 5.69 | 2/31 | 0.008 * | 0.275† | 0.019 * | 1.04† | 0.003 * | 1.17† | 0.643 | 0.29 |
| EPM: Time open arm (%) | 19.44 | 15.01 | 23.56 | 20.30 | 33.50 | 25.72 | 1.43 | 2/31 | 0.256 | 0.087 | 0.595 | 0.23 | 0.127 | 0.67 † | 0.321 | 0.43 |
| SOR: Discriminat.ratio (%) | 44.90 | 10.50 | 16.22 | 23.51 | 32.25 | 37.41 | 3.02 | 2/31 | 0.064 | 0.167† | 0.020 * | 1.58† | 0.251 | 0.46 | 0.180 | 0.51† |
| YA | AL | CR | Factor Group | YA vs. AL | YA vs. CR | AL vs. CR | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | F | df | p | η2 | p | |d| | p | |d| | p | |d| | |
| 1 Neural density | ||||||||||||||||
| NeuN+ (cells/mm2) | 45.50 | 8.62 | 32.66 | 9.89 | 48.99 | 7.54 | 6.29 | 2/16 | 0.010 * | 0.440† | 0.019 * | 1.38† | 0.503 | 0.43 | 0.004 | 1.86† |
| Hilus area (mm2) | 3.16 | 0.48 | 2.87 | 0.35 | 3.15 | 0.50 | 0.94 | 2/16 | 0.409 | 0.106† | 0.248 | 0.70† | 0.964 | 0.02 | 0.267 | 0.66† |
| 2 Immature neurons | ||||||||||||||||
| NeuroDCX | 139.18 | 39.17 | 4.93 | 5.01 | 2.94 | 3.47 | 94.10 | 2/20 | 0.001 * | 0.904† | 0.001 * | 4.81† | 0.001 * | 4.90† | 0.856 | 0.46 |
| 3 Microglial activation | ||||||||||||||||
| Iba-1 area (μm2) | 35,458.8 | 21,180.5 | 63,205.4 | 16,976.0 | 50,533.2 | 25,811.3 | 2.18 | 2/15 | 0.148 | 0.225† | 0.054 | 1.45† | 0.259 | 0.64† | 0.316 | 0.58† |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portero-Tresserra, M.; Galofré-López, N.; Pallares, E.; Gimenez-Montes, C.; Barcia, C.; Granero, R.; Rojic-Becker, D.; Vale-Martínez, A.; Martí-Nicolovius, M.; Guillazo-Blanch, G. Effects of Caloric Restriction on Spatial Object Recognition Memory, Hippocampal Neuron Loss and Neuroinflammation in Aged Rats. Nutrients 2023, 15, 1572. https://doi.org/10.3390/nu15071572
Portero-Tresserra M, Galofré-López N, Pallares E, Gimenez-Montes C, Barcia C, Granero R, Rojic-Becker D, Vale-Martínez A, Martí-Nicolovius M, Guillazo-Blanch G. Effects of Caloric Restriction on Spatial Object Recognition Memory, Hippocampal Neuron Loss and Neuroinflammation in Aged Rats. Nutrients. 2023; 15(7):1572. https://doi.org/10.3390/nu15071572
Chicago/Turabian StylePortero-Tresserra, Marta, Neus Galofré-López, Elisabet Pallares, Claudia Gimenez-Montes, Carlos Barcia, Roser Granero, Divka Rojic-Becker, Anna Vale-Martínez, Margarita Martí-Nicolovius, and Gemma Guillazo-Blanch. 2023. "Effects of Caloric Restriction on Spatial Object Recognition Memory, Hippocampal Neuron Loss and Neuroinflammation in Aged Rats" Nutrients 15, no. 7: 1572. https://doi.org/10.3390/nu15071572
APA StylePortero-Tresserra, M., Galofré-López, N., Pallares, E., Gimenez-Montes, C., Barcia, C., Granero, R., Rojic-Becker, D., Vale-Martínez, A., Martí-Nicolovius, M., & Guillazo-Blanch, G. (2023). Effects of Caloric Restriction on Spatial Object Recognition Memory, Hippocampal Neuron Loss and Neuroinflammation in Aged Rats. Nutrients, 15(7), 1572. https://doi.org/10.3390/nu15071572