1. Introduction
Copper (Cu) is a trace element involved in many physiological processes in the living organism. Importantly, Cu is a cofactor of numerous metabolic enzymes, engaged in the energy production in the respiratory chain, free radical deactivation, the maintenance of connective tissue and the proper function or modulation of nerve conduction as a result of the conversion of dopamine to norepinephrine [
1,
2,
3,
4,
5,
6,
7]. This element also stimulates the circulatory system by iron metabolism, as well as the coagulation process and blood pressure regulation [
2,
6,
7]. In order to ensure the proper course of the above-mentioned physiological processes in the animal’s body, it is therefore very important to provide a constant source of Cu in the diet. For this purpose, the diet is most often supplemented with a mineral mixture containing Cu at a level consistent with dietary recommendations, most often in the form of inorganic salt like CuCO
3 [
8,
9,
10]. However, in recent years, researchers have been giving Cu nanoparticles (CuNPs) more attention due to the specific properties determining their higher bioavailability [
8,
9,
10]. We previously showed that CuNPs included in the diet of rats are better absorbed from the gastrointestinal tract than standard CuCO
3 [
11], but most of all, they stimulate the immune system [
12] and reduce the level of DNA methylation or oxidative damage to proteins and DNA [
13]. Nevertheless, others indicated that the application of CuNPs in animal nutrition, apart from the benefits, may carry a significant risk due to the potential toxicity of CuNPs [
11,
13,
14,
15,
16,
17]. Our results are partly consistent with these reports, as we proved an increase in Cu accumulation in the brain, kidneys and lungs, as well as an intensification of lipid peroxidation or impairment of antioxidant defence [
18]. Due to such a broad physiological effect of CuNPs in the body, indicating positive and negative effects, it may be important to regulate the area of their reactivity.
Dietary fibre is one of the most important components of the diet, apart from nutrients and microelements. It is defined as parts of plants or analogous carbohydrates that are resistant to digestion and absorption in the small intestine of humans and animals, with the participation of endogenous enzymes, and are completely or partially fermented in the large intestine [
19]. Dietary fibre is usually divided into soluble and insoluble fibre. Soluble fibre includes fibre which can absorb water and thus significantly increase its volume. This group includes, above all, viscous forming a gel in contact with water pectin and sticky, swelling psyllium, but also non-viscous inulin of significant prebiotic importance. The best-known example of insoluble fibre is cellulose, which does not show a significant tendency to bind water in the digestive tract [
20]. Reports indicate that a diet rich in dietary fibre supports the gut microbiome and reduces the risk of cardiovascular diseases and colorectal cancer development [
21]. The data also suggest that inclusion of dietary fibre in the diet may modify the processes of mineral absorption in the small intestine [
22,
23,
24,
25,
26]. Moreover, the absorption of micronutrients and their impact on the intestinal tissue largely depends on the type and amount of fibre in the diet, and also minerals homeostasis in the body [
27]. It has been reported that manipulation of the fibre content of the diet may have an indirect effect on Cu bioavailability by altering the bioavailability of mineral antagonists [
28]. The effect of vegetable fibres on copper absorption is varied. It has been reported that dietary α-cellulose did not reduce copper absorption [
29]; hence, we assumed that our choice of α-cellulose as control fibre has been justified. Different dietary fibres substantially effect the small and large intestinal luminal pH values, e.g., through the enhancement of water binding or increased viscosity, as well as the stimulation of microbiota in SCFA production. The increase in intestinal pH affects copper absorbability, presumably because of a diminished concentration of free copper. There is evidence that the fraction of Cu tightly bound to bile remains unabsorbable during its passage through the gastrointestinal tract. Some studies have shown that hemicellulose induced a negative copper balance in adolescent human males, though pectin and intact cellulose were inactive. Other refined fibres and gums, such as locust bean and karaya gums, as well as carboxymethylcellulose, have been shown to be either without effect or beneficial to the trace element balance, including copper [
30].
Therefore, we assumed that the dietary combination of CuNPs with different types of fibre—neutral cellulose (control), prebiotic inulin, sticky pectin or swelling psyllium—would affect the physiological intestinal response, thereby regulating the metabolic effect of CuNPs in the body. The aim of the study was to verify the hypothesis regarding the effect of recommended Cu/kg concentration (6.5 mg) or the two times higher (13.0 mg) level of CuNPs in the diet, in combination with different types of dietary fibre (i.e., cellulose, pectin, inulin or psyllium) on selected biological parameters of intestinal integrity and histology of the small intestine in rats.
4. Discussion
The results of the study did not show that the replacement of the recommended level of Cu by CuNPs affected the diet intake and body weight of experimental rats, which is fully consistent with the results of our previous studies on rats [
11]. There were also no significant changes in the analysed growth parameters in response to the increasing CuNPs level. However, a decrease consumption and body weight were noted in rats whose diet was supplemented with inulin or psyllium. Others also report such observations [
25,
34,
35,
36,
37,
38,
39]. An explanation may be that dietary fibre naturally increases the feeling of satiety, consequently reducing the overall caloric intake [
37]. The observed effects caused by inulin or psyllium may be related to the fact that both forms dissolve well in water, forming a gel in the small intestine, which prolongs the feeling of satiety, thus reducing the intake of energy and weight loss, or to the specific rheological properties of high-fibre foods that require longer chewing [
40]. Adam et al. [
37] prove that soluble dietary fibre of a prebiotic nature stimulates the secretion of satiety hormones, such as glucagon-like peptide-1 (GLP-1) and tyrosine-tyrosine peptide (PYY), by the mucosa of the small intestine, which effectively suppresses appetite and reduces the food intake. Moreover, in their studies, the authors have also shown that the stimulation of the secretion of satiety hormones may occur as a result of the activation of receptors located in the small intestine by an increased amount of signalling short-chain fatty acids (SCFAs) produced during fibre fermentation [
37]. Interestingly, despite the significant role of pectin in dietary intake and body weight reduction [
37], the results of our study did not confirm the significant effect of this form of fibre on the reduction of growth parameters in rats. In turn, our results are consistent with others [
41,
42], and proved that a diet containing pectin is well tolerated, and no changes in the amount of food consumed or body weight were noted. Taken together, the results presented here and elsewhere suggest that the observed effects are correlated with dietary pectin levels. It seems possible that the inclusion of a higher proportion of this type of fibre in the diet could result in a decrease in the growth rate of rats tested.
Further, the replacement of CuCO
3 by CuNPs, as well as a two-fold increase in their level compared to dietary recommendations had no effect on intestinal parameters, such as small intestine weight with contents, and the dry matter, viscosity or pH of the ileum. However, these parameters can be modulated by introducing alternative forms of dietary fibre to the diet. The results of our study showed an increase in the weight of the small intestine with its content in rats fed a diet containing pectin, inulin or psyllium. Our results are consistent with others [
42,
43,
44,
45]. Dongowski et al. [
42] noted an increase in the weight of the ileum tissue and its content in rats fed a diet containing pectin. A similar increase in the weight of the small intestine in rats fed a diet containing pectin for 20 days was also noted by Pirman et al. [
46]. Moreover, Krupa-Kozak et al. [
45] proved that a gluten-free diet supplemented with prebiotic inulin for 6 weeks also showed a significant increase the relative weight of the small intestine in rats. Arjmandi et al. [
43] observed an increase in the relative weight of the small intestine in rats fed a diet containing cholesterol and 5 or 10% psyllium for 21 days, which was stored at 5 °C or 40 °C for 8 months, compared to a control group receiving a diet containing 10% cellulose. Kristensen et al. [
44] also observed analogous changes as a significant increase in the weight of the gastrointestinal tract in total and its individual segments, including the small intestine, in rats receiving a dietary fibre supplement from linseed. Dongowski et al. [
42] explain the obtained effect by the fact that dietary fibre (especially pectin) is not or only slightly degraded in the small intestine of rats, which means that the fibre contained in the gastrointestinal content binds water, thereby increasing its viscosity. The small intestine has a difficult task because it has to transport this sticky content to the lower segments of the digestive tract, which causes its cells to grow, and, as a consequence, its mass also increases. Although our study confirmed an increase in ileal viscosity in rats fed a diet supplemented with pectin and psyllium, histological evaluation did not reveal an overgrowth of the small intestinal wall cells in any of the experimental groups. In our study, an increase in the acidity of the ileum of rats fed a diet with the supplementation of pectin and inulin was also shown. Krupa-Kozak et al. [
45] suggest that an increase in the weight of the small intestine with simultaneous acidification of its environment may indicate that inulin (and perhaps other soluble forms of fibre) may be metabolised to some extent by air-tolerant small intestine bacteria. The increased weight of the small intestine with its content obtained in our research, with a simultaneous reduction in the dry matter of the ileum in rats from all experimental groups, and an increase in the viscosity of the small intestine of rats receiving pectin or psyllium, allows us to assume that the observed changes may be primarily the result of hydration properties of dietary fibre. All three forms of tested dietary fibre (inulin, pectin and psyllium) have a tendency to dissolve in water and bind it, wherein pectin forms very viscous solutions, and psyllium swells when binding water [
37]. Food containing fibre, passing through subsequent sections of the digestive tract, up to the small intestine, binds water while increasing its volume and weight. Taking into account the significant increase in the total weight of the small intestine with the simultaneous decrease in the dry matter of the ileum in all experimental groups, it may be assumed that these differences were affected by the weight of water bound by dietary fibre present in the intestinal content.
In the assessment of the intestinal barrier function, diamine oxidase (DAO) is often considered. This enzyme catalyses the decomposition reaction of histamine in the digestive tract, which is responsible for the development of an allergic reaction. Under physiological conditions, a relatively high level of DAO is found in the small intestine, while in blood plasma it is very low. When the intestinal mucosa is damaged, e.g., as a result of ischemia, hypoxia, contact with a harmful factor present in food or tissue nutrition disorders, an inflammatory reaction is triggered with the release of histamine. Due to the necessity of its deactivation, a decrease in the level of DAO in the intestinal mucosa is observed, resulting in a simultaneous decrease in the level of this indicator in the blood [
47]. Our results indicate that the replacement of Cu in the standard form of CuCO
3 by an equivalent dose of CuNPs had no effect on the DAO level in the blood plasma and small intestine. However, the level of DAO in the blood plasma increased with no changes in the level of this indicator in the small intestine as a result of increasing the CuNPs dietary level. Surprisingly, the results of our study also showed no significant differences in the level of intestinal DAO between the experimental groups and control group fed a diet containing the addition of cellulose, while the value of this indicator was significantly increased in the blood plasma of rats receiving a diet containing the addition of inulin or psyllium. Although the obtained results are ambiguous, they undoubtedly indicate a beneficial effect of both the introduction of a doubled dose of CuNPs compared to the standard recommendation, as well as alternative forms of fibre (inulin and psyllium) on maintaining a proper intestinal barrier.
Lactic acid is the end product of glucose oxidation in anaerobic glycolysis [
48]. It may be synthesised in situ on the intestinal mucosa by anaerobic bacteria colonising the lumen of the large intestine, such as lactobacilli, streptococci and bifidobacteria [
49,
50]. There are reports that this compound may affect a number of metabolic and immunological processes in the body, including mediation in the signalling pathways; the production of pro- and anti-inflammatory mediators by T lymphocytes and macrophages; or by affecting the redox status through the reaction of lactate dehydrogenase inducing reactive oxygen species and acting as an inhibitor of glucose breakdown [
49]. Okada et al. [
51], in studies on mice, also proved that lactate can stimulate the proliferation of enterocytes, which has a positive effect on maintaining the function of the intestinal barrier. However, there are also studies that found an increase in the synthesis of lactic acid in the intestines and the accompanying discharge of this compound into the blood, which may be observed in the case of intestinal hypoxia, resulting in the intensification of anaerobic glucose breakdown. As a consequence, it may lead to dangerous lactic acidosis development [
48]. The results of our study indicate that replacing the traditional CuCO
3 with CuNPs resulted in a reduction in the level of lactic acid in the small intestine and blood serum as well. In light of the above-mentioned reports, this effect may be considered positive. In addition, the increased lactic acid level in the intestine may result in acidification of the environment. In the case of the large intestine, this effect may be desirable, because it protects it, among others, from colonisation by pathogenic bacteria [
52], while, in the case of the small intestine, this effect may be completely different. Digestive enzymes secreted into the lumen of the small intestine require a slightly alkaline environment for their activation and proper functioning [
53]. Acidification of the environment caused by the overproduction of lactic acid in the small intestine may therefore result in a reduction in digestive processes, and thus, in the efficiency of the absorption of nutrients. This also seems to confirm the beneficial effect of CuNPs on the functioning of the small intestine. Moreover, our study also noted a significant DxF interaction for LA. This results from the fact that, in the case of the combined use of pectin or psyllium as a source of fibre with a higher level of CuNPs, a reduced level of this indicator was found in the intestinal wall, which was not observed in the case of the combined use of nanoparticles with cellulose or pectin. It is likely that the use of a higher level of CuNPs with the above-mentioned sources of fibre has a more beneficial effect on intestinal integrity than its combination with cellulose, as also indicated by the increased level of LA in the blood plasma of rats from the cellulose group, together with a higher level of CuNPs in the diet. Increased production of lactic acid in the small intestine is an unfavourable phenomenon, as it can lead to acidosis, which results in serious damage to the intestinal epithelium, resulting in excessive intestinal permeability, referred to as “leaky gut” [
54].
Among all organs, the tissues of the gastrointestinal tract, especially the small intestine, is most exposed to the potentially harmful effects of xenobiotics entering the body through diet. It is assumed that this harmful effect is very often associated with the increased synthesis of free oxygen radicals. These, in turn, contribute to the occurrence of oxidative stress, resulting in damage to cellular macromolecules, including DNA modifications such as the oxidation of nitrogenous bases [
55]. In order to prevent the loss of genome integrity, the body has developed various repair systems, among which a very important role is played the DNA repair pathway by cutting out damaged bases (BER) and replacing them with correct ones. This process is initiated by DNA glycosylases, which include, among others, 8-oxoguanine glycosylase (OGG1). This enzyme removes the most common DNA damages such as 8-hydroxydeoxyguanosine (8-OHdG) and 2,6-diamino-4-hydroxy-5-formamidopyrimidine (FapyGua) [
56]. The results of our previous studies showed no deterioration of the oxidoreductive status of the small intestine in rats due to the replacement of the standard form of Cu by CuNPs [
31]. The highly reactive CuNPs may undergo Fenton and/or Haber–Weiss reactions in the body, resulting in the enhanced production of free radicals, which then damage the genetic material [
31]. The results of the present study partially confirm that the replacement of CuCO
3 by CuNPs resulted in an increase in the level of 8-OHdG in the small intestine, which suggests the intensification of oxidative processes in the examined tissue. Interestingly, the inclusion of the recommended level of CuNPs in the diet of rats contributed to a downregulation of
OGG1 gene expression in the small intestine, which, however, did not translate into a decrease in the amount of functional OGG1 protein. It was also not found that increasing the level of CuNPs in the diet of rats from 6.5 to 13 mg Cu/kg of diet resulted in a deterioration of the oxidoreductive status and weakening of DNA repair mechanisms in the small intestine. However, an increase in the OGG1 protein level was noted, with no changes in
OGG1 gene expression and 8-OHdG content in rats supplemented with inulin. This suggests that excessive oxidation of nitrogenous bases does not occur, and thus, the OGG1 enzyme is not significantly used for their repair. This allows us to assume that among all the tested forms of fibre, inulin best protects DNA against the harmful effects of free radicals, which may potentially be formed as a result of including CuNPs in the diet. The beneficial effect of the combined use of pectin with a higher level of CuNPs is evidenced by the observed DxF interaction for the level of APE-1 and for the expression of the
OGG1 gene in the small intestine, resulting from the fact that only in the case of combining pectin as a source of fibre with the addition of a higher level of CuNPs, an increase in the level of APE-1 and downregulation of
OGG1 were noted. Although an increase in the APE-1 level (apurinic/apyrimidinic endonuclease 1, one of the DNA repair enzymes) may indicate an increased oxidation of this acid [
57]. However, due to the fact that an increased level of 8-OHdG was not observed when a higher level of CuNPs was used together with pectin compared to the use of a lower level of CuNPs with pectin, it should be concluded that the increase in the level of APE-1 was not associated with the induction of the repair mechanism.
In the event of damage to important cellular macromolecules, the programmed cell death may be activated. Caspases play an important role in this multi-stage process [
58]. The results of our study indicate that the replacement of CuCO
3 by CuNPs resulted in a decrease in caspase 3 and caspase 8 levels in the small intestine of rats, which seems to be a very beneficial phenomenon, and proves that this supplement does not induce significant negative changes in the cells of the small intestine. It also allows us to assume that the previously mentioned induction of oxidative DNA damage as a result of including CuNPs in the diet of rats was not serious enough to lead to significant changes in the genome of intestinal cells, which would result in their being directed to the apoptotic pathway. Moreover, it was not found that increasing the level of CuNPs in the diet contributed to the occurrence of negative changes in the level of the tested caspases. Regardless of the level of CuNPs introduced into the rats’ diet, no increase in the synthesis of caspases was found as a result of inulin and psyllium inclusion in the diet, and the addition of pectin reduced the level of caspase 3 in the wall of the small intestine. This suggests that, among the studied forms of fibre, pectin best protects the cells of the small intestine against damage that could lead to their apoptosis.
The intestinal barrier is crucial for maintaining intestinal homeostasis, which determines the proper supply of the body with nutrients and the prevention of intestinal diseases. It consists of the apical cell membrane and intercellular tight junctions (TJ) of enterocytes. Tight junctions are protein complexes resulting from the interaction between members of the claudin family, zonula occludens and MARVEL tight junction proteins (TAMP) [
59]. ZO-1 is a cytoplasmic plaque protein that recruits various signalling molecules and acts as a scaffold for transmembrane TJ proteins [
60]. This protein is encoded by the
ZO-1 gene, which is expressed especially in the intestine, kidneys, liver, lungs and brain [
61]. There are reports that a decrease in ZO-1 protein expression may increase intestinal permeability, which, in turn, may result in the development of intestinal inflammation and even the development of neoplastic lesions [
59,
62]. It is assumed that Trefoil family factor 2 (TFF2), encoded by the gene of the same name, also plays an important role in maintaining a proper intestinal barrier [
63]. The results of our study showed that
TFF2 may be expressed in the wall of the small intestine. Interestingly, the
TFF2 protein is involved in intestinal defence and repair mechanisms, while
TFF2 overexpression is often observed in the case of significant tissue damage, infection or neoplastic changes in the within the digestive tract [
63]. The results of our studies indicate that the replacement of CuCO
3 by CuNPs in the diet of rats resulted in
TFF2 and
ZO-1 downregulation, with no changes in the expression level of
OCLN. However, there was no effect of doubling the level of CuNPs in the diet—in relation to the nutritional recommendations—on the level of expression of the analysed genes of the intestinal barrier in rats. The obtained results can be interpreted in two ways, because the downregulation of
TFF2 gene expression by CuNPs seems to be beneficial, but a decrease in the level of
ZO-1 may indicate that CuNPs increase intestinal permeability and, thus, deteriorate the intestinal barrier. The explanation of the obtained results may be related to the formation of a functional protein, which reduces the expression of the
TFF2 and
ZO-1 mRNA pools. The results of our study also showed that the inclusion of dietary fibre in the form of psyllium in the diet of rats increased the
ZO-1 gene expression in the small intestine, which, in light of reports by other authors, is an effect conducive to maintaining the proper integrity of the intestinal barrier [
59,
62].
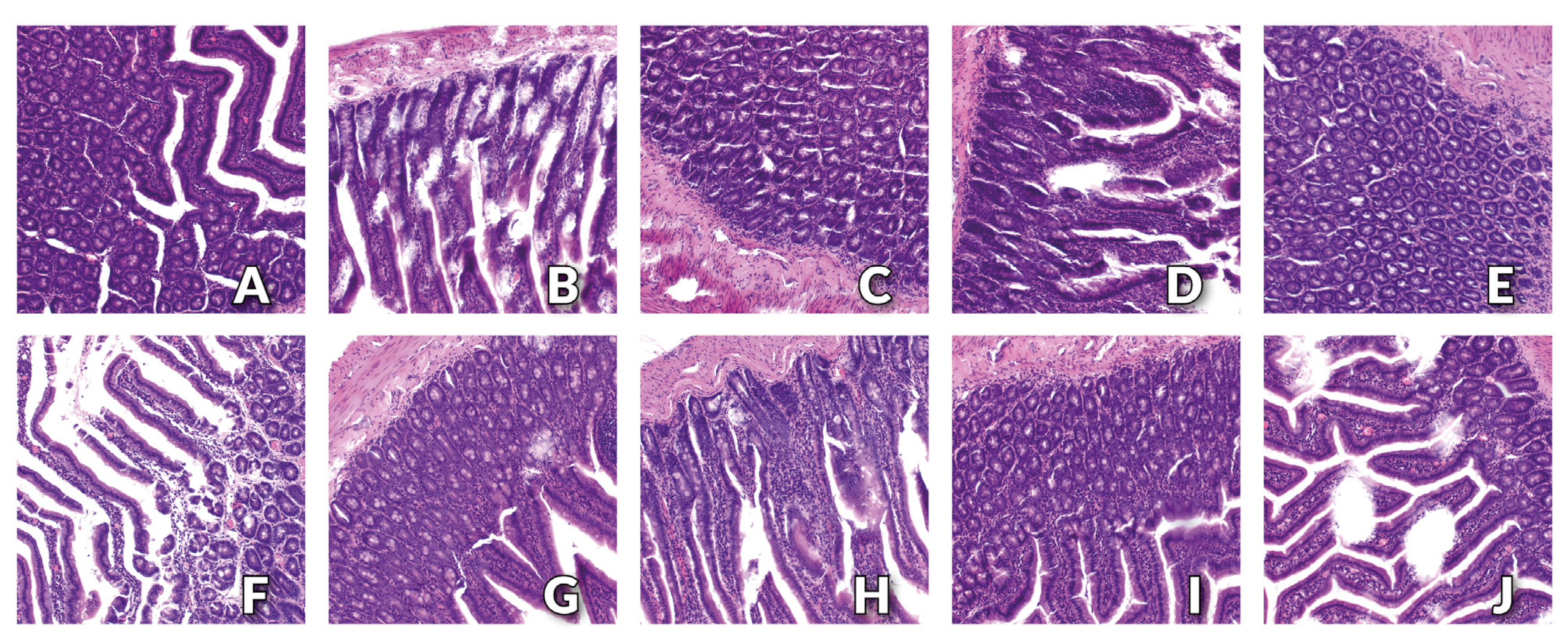
Finally, the results of histopathological examination indicate the normal structure of the small intestine with the presence of small tissue defects in the apical parts of the villi in rats receiving a standard diet containing the recommended level of Cu in the form of CuCO
3 and cellulose as a source of fibre. We observed no adverse changes in the morphology of the small intestine in rats receiving a diet containing the 6.5 mg/kg of CuNPs and the addition of cellulose (control group) or pectin. For rats fed a diet containing the same level of CuNPs but supplemented with inulin or psyllium, single tissue defects were observed at the top of the intestinal villi. Similar changes were intensified by the inclusion of a higher level of CuNPs (13 mg/kg) in the diet, regardless of the form of fibre used, with the most significant and numerous losses of the top part of the intestinal villi observed in the group receiving cellulose in the diet. The obtained results indicate that the replacement of CuCO
3 by CuNPs had a beneficial effect on the morphology of the intestines. In turn, changes in the top parts of the villi observed in groups of rats fed a diet containing an increased level of CuNPs, regardless of the fibre source used, seem to be related to the direct effect of Cu in the form of nanoparticles. However, the lack of any pathological changes at the base of the villi and in the mucosa of the small intestine, regardless of the type of dietary fibre, as well as the lack of deterioration of indicators proving the integrity of the intestinal barrier, do not seem to confirm this assumption. Moreover, our previous studies did not show any pathological changes in the small intestine tissue of healthy, normotensive rats, in which both 50% and 100% of the traditional CuCO
3 was replaced by CuNPs. In the histological image of the small intestine of rats receiving the recommended level of Cu in the diet only in the form of CuCO
3, isolated extensive changes in the base of intestinal villi were found [
31]. In light of the above, it can be assumed that the observed damage to the top parts of the intestinal villi may be the result of a change in the rheology of the gastrointestinal content due to the presence of dietary fibre. Dietary fibre, binding water, increases its volume, and thus, the pressure of intestinal contents on the walls of the small intestine. It is therefore highly probable that the movement of the swollen, viscous chyme through successive fragments of the small intestine was accompanied by greater friction, thus leading to damage to the top parts of the intestinal villi.
Numerous reports in the literature indicate that the use of Cu nanoparticles in animal nutrition may be associated with many risks, such as their increased accumulation in internal organs [
11,
17]. Henson et al. [
64], in an in vitro model study, showed that CuO nanoparticles are much more cytotoxic to rat intestinal epithelial cells (IEC-6) than Cu
2+ ions, and this effect is stronger the higher their dose and the longer the exposure time. The authors observed that increased production of free oxygen radicals resulted in damage to the mitochondrial membrane and reduced viability of small intestine cells exposed to CuO NPs [
64]. Lee et al. [
17] suggest that the toxicity of copper nanoparticles may be a result of the fact that, in the acidic conditions of gastric juice, they can dissociate to Cu
2+ ions. However, the authors did not record a similar phenomenon in intestinal juice [
17]. Henson et al. also confirmed that there was no release of Cu
2+ ions from CuO NPs in the intestinal juice [
64]. According to the authors, this may indicate that the cytotoxicity of CuO NPs towards intestinal cells is not dependent on their dissociation into the ionic form, but on the specific and natural nature of this form of Cu [
64]. The results of our previous studies have also shown that CuNPs are absorbed in the small intestine to a much greater extent than standard forms of this element, such as CuCO
3 [
11], which, in light of the results obtained by Lee et al. [
17] and Henson et al. [
64], should result in damage to the intestinal barrier and the occurrence of pathological changes in the examined tissue. The results of our study, apart from the possibility of intensifying DNA oxidation by CuNPs, did not confirm this assumption, indicating the highly toxic effect of CuNPs disrupting the integrity of the intestinal barrier, even when used in a dose two times higher than the nutritional recommendations for rats. The introduction of twice as high levels of CuNPs into the diet did not have a significant effect on the increased level of this element in the blood plasma (unpublished data). This effect may therefore be related to the direct effect of dietary fibre introduced into the diet. So far, it has not been possible to clearly determine the effect of different types of dietary fibre on the intestinal absorption of minerals. There are reports indicating both a beneficial and negative effect of dietary fibre [
22,
23,
24,
25]. According to Coudray [
27], mineral absorption also largely depends on the type and amount of fibre in the diet, as well as mineral homeostasis in the body. Coudray et al. [
25] showed an increase in the absorption and retention of Cu in the intestines as a result of inulin consumption. In turn, Krzysik et al. [
26] observed a decrease in the absorption of divalent ions as a result of feeding rats a diet containing pectin or cellulose. The decrease in the absorption of trace elements in the small intestine is probably due to the fact that fibre (especially pectin and psyllium) forms a kind of sticky gel in the digestive tract, which is able to bind mainly divalent ions (including Cu
2+) with free carboxyl groups, which reduces their bioavailability [
23,
26,
65]. Furthermore, pectin may also affect the absorption of minerals by stimulating the bacterial production of short-chain fatty acids (SCFA) and acidifying the intestinal lumen, thus creating unfavourable conditions for the absorption process and intensifying the dissociation of CuNPs to divalent ions [
8,
26,
66]. In addition, the presence of dietary fibre in the diet accelerates the intestinal transit, as a result of which the food content is much shorter in the small intestine, which may significantly reduce the amount of nutrients and minerals absorbed into the body [
26,
67]. Taken together, it seems that the supplementation of dietary fibre in the form of pectin, inulin and psyllium reduced the absorption of CuNPs, thus protecting the small intestine and modulating its biological response.


 ,
, 

{kind=link}