

The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake

 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Participants
2.2.1. Resistance Exercise Study
2.2.2. Habitual Coffee Intake Study
2.3. Resistance Exercise Study Design
2.4. Assessment of Habitual Coffee Intake
2.5. Biochemical Analysis
2.6. Genotyping
2.6.1. Resistance Exercise Study
2.6.2. Habitual Coffee Intake Study
2.7. Statistical Analysis
3. Results
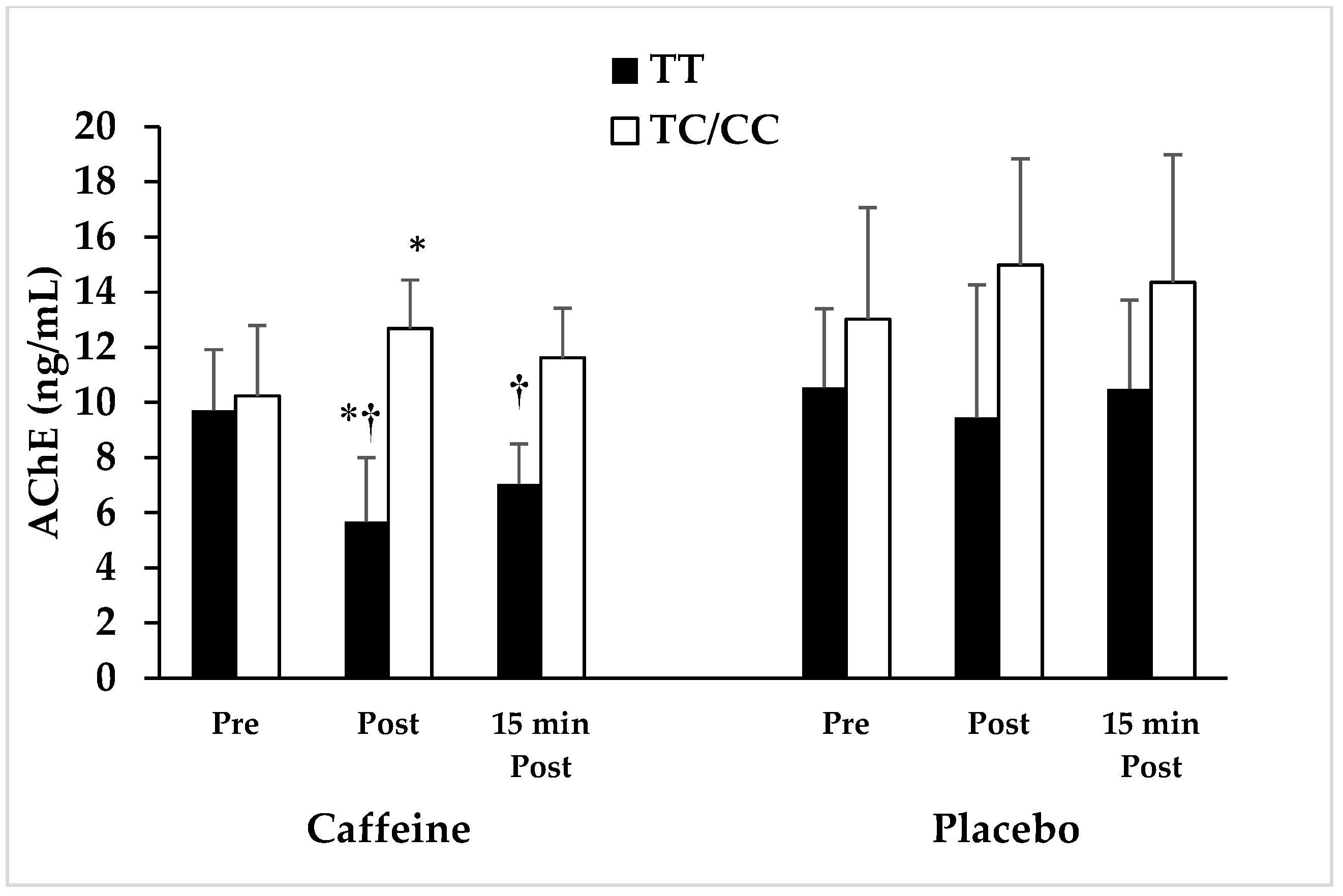
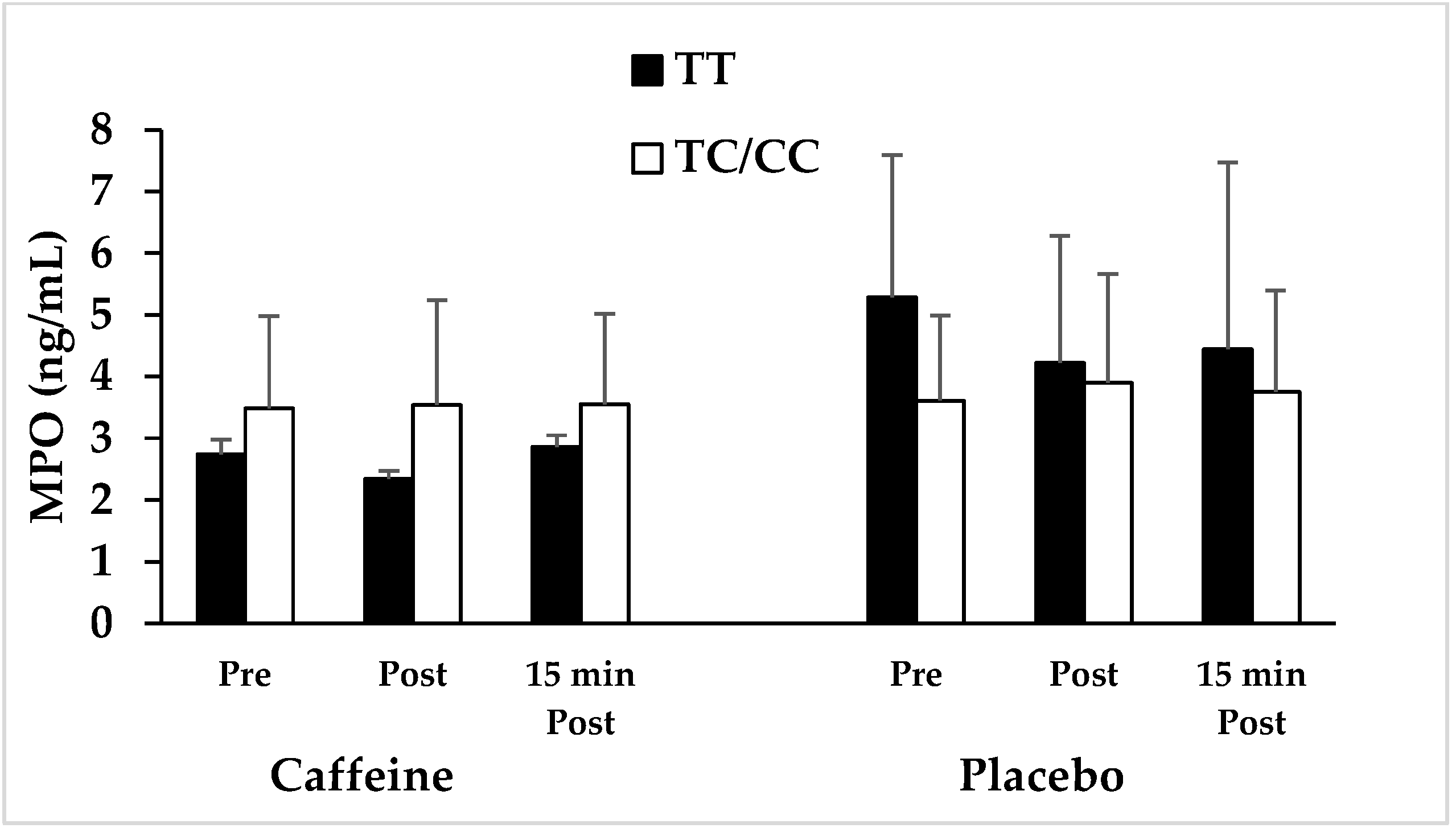
3.1. Association of ADORA2A Variant with Anti-Inflammatory Effects of Caffeine in Response to Intense Resistance Exercise
3.2. Association of ADORA2A Variant with Habitual Coffee Intake
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ginevičienė, V.; Utkus, A.; Pranckevičienė, E.; Semenova, E.A.; Hall, E.C.R.; Ahmetov, I.I. Perspectives in Sports Genomics. Biomedicines 2022, 10, 298. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Hall, E.C.R.; Semenova, E.A.; Pranckevičienė, E.; Ginevičienė, V. Advances in sports genomics. Adv. Clin. Chem. 2022, 107, 215–263. [Google Scholar] [CrossRef]
- Womack, C.J.; Saunders, M.J.; Bechtel, M.K.; Bolton, D.J.; Martin, M.; Luden, N.D.; Dunham, W.; Hancock, M. The influence of a CYP1A2 polymorphism on the ergogenic effects of caffeine. J. Int. Soc. Sports Nutr. 2012, 9, 7. [Google Scholar] [CrossRef]
- Guest, N.S.; Horne, J.; Vanderhout, S.M.; El-Sohemy, A. Sport Nutrigenomics: Personalized Nutrition for Athletic Performance. Front. Nutr. 2019, 6, 8. [Google Scholar] [CrossRef]
- Hall, E.C.R.; Semenova, E.A.; Bondareva, E.A.; Andryushchenko, L.B.; Larin, A.K.; Cięszczyk, P.; Generozov, E.V.; Ahmetov, I.I. Association of Genetically Predicted BCAA Levels with Muscle Fiber Size in Athletes Consuming Protein. Genes 2022, 13, 397. [Google Scholar] [CrossRef]
- Smart, D.; Chipman, J.; Hodges, N. Activity of OGG1 variants in the repair of pro-oxidant-induced 8-oxo-2′-deoxyguanosine. DNA Repair 2006, 5, 1337–1345. [Google Scholar] [CrossRef]
- Mirzaei, B.; Rahmani-Nia, F.; Salehi, Z.; Rahimi, R.; Faraji Saravani, M.; Ahmadi, R.; Mousavi Beneh Hoor, Z. Effects of GSTM1 gene polymorphism on oxidative DNA damage and lipid peroxidation following exhaustive exercise in young wrestlers. Med. Dello Sport 2014, 67, 437–448. [Google Scholar]
- Rahimi, R.; Salehi, Z.; Saravani, M.F.; Hoor, Z.M.B.; Darvishi, S. Is high-intensity resistance exercise-induced oxidative DNA damage related to OGG1 Ser326Cys polymorphism in athletes? Sport Sci. Health 2014, 10, 159–163. [Google Scholar] [CrossRef]
- Ahmetov, I.I.; Naumov, V.A.; Donnikov, A.E.; Maciejewska-Karłowska, A.; Kostryukova, E.S.; Larin, A.K.; Maykova, E.V.; Alexeev, D.G.; Fedotovskaya, O.N.; Generozov, E.V.; et al. SOD2 gene polymorphism and muscle damage markers in elite athletes. Free Radic. Res. 2014, 48, 948–955. [Google Scholar] [CrossRef]
- Zarebska, A.; Jastrzebski, Z.; Ahmetov, I.I.; Zmijewski, P.; Cieszczyk, P.; Leonska-Duniec, A.; Sawczuk, M.; Leznicka, K.; Trybek, G.; Semenova, E.A.; et al. GSTP1 c.313A>G polymorphism in Russian and Polish athletes. Physiol. Genom. 2017, 49, 127–131. [Google Scholar] [CrossRef]
- Shanker, J.; Kakkar, V.V. Implications of genetic polymorphisms in inflammation-induced atherosclerosis. Open Cardiovasc. Med. J. 2010, 4, 30. [Google Scholar] [CrossRef]
- Dionísio, T.J.; Thiengo, C.R.; Brozoski, D.T.; Dionísio, E.J.; Talamoni, G.A.; Silva, R.B.; Garlet, G.P.; Santos, C.F.; Amaral, S.L. The influence of genetic polymorphisms on performance and cardiac and hemodynamic parameters among Brazilian soccer players. Appl. Physiol. Nutr. Metab. 2017, 42, 596–604. [Google Scholar] [CrossRef]
- Simpson, R.E.; Phillis, J.W. Adenosine in exercise adaptation. Br. J. Sport. Med. 1992, 26, 54–58. [Google Scholar] [CrossRef]
- Ralevic, V.; Burnstock, G. Receptors for purines and pyrimidines. Pharmacol. Rev. 1998, 50, 413–492. [Google Scholar]
- Mazziotta, C.; Rotondo, J.C.; Lanzillotti, C.; Campione, G.; Martini, F.; Tognon, M. Cancer biology and molecular genetics of A3 adenosine receptor. Oncogene 2022, 41, 301–308. [Google Scholar] [CrossRef]
- Latini, S.; Pedata, F. Adenosine in the central nervous system: Release mechanisms and extracellular concentrations. J. Neurochem. 2001, 79, 463–484. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Arslan, G.; Halldner, L.; Kull, B.; Schulte, G.; Wasserman, W. Structure and function of adenosine receptors and their genes. Naunyn Schmiedebergs Arch. Pharmacol. 2000, 362, 364–374. [Google Scholar] [CrossRef]
- Okada, M.; Kiryu, K.; Kawata, Y.; Mizuno, K.; Wada, K.; Tasaki, H.; Kaneko, S. Determination of the effects of caffeine and carbamazepine on striatal dopamine release by in vivo microdialysis. Eur. J. Pharmacol. 1997, 321, 181–188. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Chen, J.-F.; Masino, S.A.; Vaugeois, J.-M. Actions of adenosine at its receptors in the CNS: Insights from knockouts and drugs. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 385–412. [Google Scholar] [CrossRef]
- Davis, J.; Zhao, Z.; Stock, H.S.; Mehl, K.A.; Buggy, J.; Hand, G.A. Central nervous system effects of caffeine and adenosine on fatigue. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R399–R404. [Google Scholar] [CrossRef]
- El Yacoubi, M.; Ledent, C.; Parmentier, M.; Costentin, J.; Vaugeois, J.-M. Reduced appetite for caffeine in adenosine A 2A receptor knockout mice. Eur. J. Pharmacol. 2005, 519, 290–291. [Google Scholar] [CrossRef]
- Cronstein, B.; Levin, R.; Philips, M.; Hirschhorn, R.; Abramson, S.; Weissmann, G. Neutrophil adherence to endothelium is enhanced via adenosine A1 receptors and inhibited via adenosine A2 receptors. J. Immunol. 1992, 148, 2201–2206. [Google Scholar] [CrossRef]
- Cronstein, B.N.; Kubersky, S.M.; Weissmann, G.; Hirschhorn, R. Engagement of adenosine receptors inhibits hydrogen peroxide (H2O2−) release by activated human neutrophils. Clin. Immunol. Immunopathol. 1987, 42, 76–85. [Google Scholar] [CrossRef]
- Fredholm, B.B.; Bättig, K.; Holmén, J.; Nehlig, A.; Zvartau, E.E. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol. Rev. 1999, 51, 83–133. [Google Scholar] [PubMed]
- Alsene, K.; Deckert, J.; Sand, P.; de Wit, H. Association between A2a receptor gene polymorphisms and caffeine-induced anxiety. Neuropsychopharmacology 2003, 28, 1694–1702. [Google Scholar] [CrossRef]
- Cornelis, M.C.; El-Sohemy, A.; Campos, H. Genetic polymorphism of the adenosine A2A receptor is associated with habitual caffeine consumption. Am. J. Clin. Nutr. 2007, 86, 240–244. [Google Scholar] [CrossRef]
- Rahimi, R. The effect of CYP1A2 genotype on the ergogenic properties of caffeine during resistance exercise: A randomized, double-blind, placebo-controlled, crossover study. Ir. J. Med. Sci. 2019, 188, 337–345. [Google Scholar] [CrossRef]
- Loy, B.D.; O’Connor, P.J.; Lindheimer, J.B.; Covert, S.F. Caffeine is ergogenic for adenosine A2A receptor gene (ADORA2A) T allele homozygotes: A pilot study. J. Caffeine Res. 2015, 5, 73–81. [Google Scholar] [CrossRef]
- Rogers, P.J.; Hohoff, C.; Heatherley, S.V.; Mullings, E.L.; Maxfield, P.J.; Evershed, R.P.; Deckert, J.; Nutt, D.J. Association of the anxiogenic and alerting effects of caffeine with ADORA2A and ADORA1 polymorphisms and habitual level of caffeine consumption. Neuropsychopharmacology 2010, 35, 1973–1983. [Google Scholar] [CrossRef]
- Morozov, V.I.; Pryatkin, S.A.; Kalinski, M.I.; Rogozkin, V.A. Effect of exercise to exhaustion on myeloperoxidase and lysozyme release from blood neutrophils. Eur. J. Appl. Physiol. 2003, 89, 257–262. [Google Scholar] [CrossRef]
- Morozov, V.I.; Tsyplenkov, P.V.; Golberg, N.D.; Kalinski, M.I. The effects of high-intensity exercise on skeletal muscle neutrophil myeloperoxidase in untrained and trained rats. Eur. J. Appl. Physiol. 2006, 97, 716–722. [Google Scholar] [CrossRef]
- Gleeson, M.; Walsh, N.; Blannin, A.; Robson, P.; Cook, L.; Donnelly, A.E.; Day, S.H. The effect of severe eccentric exercise-induced muscle damage on plasma elastase, glutamine and zinc concentrations. Eur. J. Appl. Physiol. Occup. Physiol. 1998, 77, 543–546. [Google Scholar] [CrossRef]
- Borregaard, N.; Cowland, J.B. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood 1997, 89, 3503–3521. [Google Scholar] [CrossRef]
- Das, U.N. Acetylcholinesterase and butyrylcholinesterase as possible markers of low-grade systemic inflammation. Med. Sci. Monit. 2007, 13, RA214–RA221. [Google Scholar] [CrossRef]
- Stefanello, S.T.; Alex, F.; Soares, R.; Barcelos, R. Caffeine supplementation changes inflammatory biomarkers after exercise. J. Yoga Phys. Ther. 2016, 6, 2. [Google Scholar] [CrossRef]
- Borovikova, L.V.; Ivanova, S.; Zhang, M.; Yang, H. Vagus nerve stimulation attenuates the systemic inflammatory response to endotoxin. Nature 2000, 405, 458–462. [Google Scholar] [CrossRef]
- Barcelos, R.P.; Souza, M.A.; Amaral, G.P.; Stefanello, S.T.; Bresciani, G.; Fighera, M.R.; Soares, F.A.A.; de Vargas Barbosa, N. Caffeine intake may modulate inflammation markers in trained rats. Nutrients 2014, 6, 1678–1690. [Google Scholar] [CrossRef]
- Haskell, C.F.; Kennedy, D.O.; Wesnes, K.A.; Scholey, A.B. Cognitive and mood improvements of cafeine in habitual consumers and habitual non-consumers of cafeine. Psychopharmacology 2005, 179, 813–825. [Google Scholar] [CrossRef]
- Erblang, M.; Drogou, C.; Gomez-Merino, D.; Metlaine, A.; Boland, A.; Deleuze, J.F.; Thomas, C.; Sauvet, F.; Chennaoui, M. The Impact of Genetic Variations in ADORA2A in the Association between Caffeine Consumption and Sleep. Genes 2019, 10, 1021. [Google Scholar] [CrossRef] [PubMed]
- Medrano, R.F.V.; de Oliveira, C.A. Guidelines for the tetraprimer ARMS–PCR technique development. Mol. Biotechnol. 2014, 56, 599–608. [Google Scholar] [CrossRef]
- Semenova, E.A.; Zempo, H.; Miyamoto-Mikami, E.; Kumagai, H.; Larin, A.K.; Sultanov, R.I.; Babalyan, K.A.; Zhelankin, A.V.; Tobina, T.; Shiose, K.; et al. Genome-Wide Association Study Identifies CDKN1A as a Novel Locus Associated with Muscle Fiber Composition. Cells 2022, 11, 3910. [Google Scholar] [CrossRef]
- Kikuchi, N.; Moreland, E.; Homma, H.; Semenova, E.A.; Saito, M.; Larin, A.K.; Kobatake, N.; Yusupov, R.A.; Okamoto, T.; Nakazato, K.; et al. Genes and Weightlifting Performance. Genes 2022, 13, 25. [Google Scholar] [CrossRef]
- Pataky, M.; Womack, C.; Saunders, M.; Goffe, J.; D’lugos, A.; El-Sohemy, A.; Luden, N. Caffeine and 3-km cycling performance: Effects of mouth rinsing, genotype, and time of day. Scand. J. Med. Sci. Sport. 2016, 26, 613–619. [Google Scholar] [CrossRef]
- Pickering, C.; Kiely, J. Are the current guidelines on caffeine use in sport optimal for everyone? Inter-individual variation in caffeine ergogenicity, and a move towards personalised sports nutrition. Sport. Med. 2018, 48, 7–16. [Google Scholar] [CrossRef]
- Thomas, R.; Algrain, H.; Ryan, E.; Popojas, A.; Carrigan, P.; Abdulrahman, A.; Carrillo, A. Influence of a CYP1A2 polymorphism on post-exercise heart rate variability in response to caffeine intake: A double-blind, placebo-controlled trial. Ir. J. Med. Sci. 2017, 186, 285–291. [Google Scholar] [CrossRef]
- Peake, J.; Wilson, G.; Hordern, M.; Suzuki, K.; Yamaya, K.; Nosaka, K.; Mackinnon, L.; Coombes, J.S. Changes in neutrophil surface receptor expression, degranulation, and respiratory burst activity after moderate-and high-intensity exercise. J. Appl. Physiol. 2004, 97, 612–618. [Google Scholar] [CrossRef]
- Haskó, G.; Cronstein, B. Regulation of inflammation by adenosine. Front. Immunol. 2013, 4, 85. [Google Scholar] [CrossRef]
- Ritter, M.; Hohenberger, K.; Alter, P.; Herzum, M.; Tebbe, J.; Maisch, M. Caffeine inhibits cytokine expression in lymphocytes. Cytokine 2005, 30, 177–181. [Google Scholar] [CrossRef]
- UK Biobank GWAS Round 2 Results [Released 1st August 2018]. Available online: http://www.nealelab.is/uk-biobank/ (accessed on 27 February 2023).



{kind=link}
{kind=link}
| Genotype Group | TT (n = 5) | CT/CC (n = 10) | T | p Value | ||
|---|---|---|---|---|---|---|
| M | SD | M | SD | |||
| Age (years) | 21.75 | 3.30 | 20.20 | 1.87 | 1.13 | 0.28 |
| Height (cm) | 179.5 | 3.69 | 178.5 | 4.67 | 0.38 | 0.71 |
| Weight (kg) | 73.87 | 6.54 | 72.47 | 10.38 | 0.24 | 0.80 |
| Soft Lean Mass (kg) | 60.55 | 4.25 | 60.38 | 7.26 | 0.04 | 0.96 |
| Body Fat Mass (kg) | 9.97 | 2.90 | 8.73 | 4.34 | 0.52 | 0.61 |
| % Body Fat | 13.32 | 3.15 | 11.71 | 4.50 | 0.64 | 0.52 |
| Body mass index (kg/m2) | 22.90 | 1.08 | 22.80 | 3.59 | 0.05 | 0.95 |
| Basal Metabolic Rate | 2026.70 | 183.64 | 2055.40 | 193.56 | −0.25 | 0.80 |
| Trait | Group | TT Genotype (n = 5) | CT/CC Genotype (n = 10) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Pre | ∆ | Post | ∆ | 15 m Post | ∆ | Pre | ∆ | Post | ∆ | 15 m Post | ∆ | ||
| MPO (ng/mL) | C | 2.7 ± 0.2 | 48.1 ↓ | 2.3 ± 0.1 | 44.5 ↓ | 2.9 ± 0.2 | 35.6 ↓ | 3.5 ± 1.5 | 3.6 ↓ | 3.5 ± 1.7 | 9.2 ↓ | 3.6 ± 1.5 | 5.3 ↓ |
| P | 5.3 ± 2.3 | 4.2 ± 2.1 | 4.4 ± 3.0 | 3.6 ± 1.4 | 3.9 ± 1.8 | 3.8 ± 1.6 | |||||||
| AChE (ng/mL) | C | 9.7 ± 2.2 | 5.1 ↓ | 5.7 ± 2.3 | 55.2 ↓ * | 7.0 ± 1.5 | 39.4 ↓ * | 10.6 ± 2.6 | 19.0 ↓ | 9.5 ± 1.8 | 36.8 ↓ * | 10.5 ± 1.8 | 26.9 ↓ |
| P | 10.2 ± 2.9 | 12.7 ± 4.8 | 11.6 ± 3.2 | 13.0 ± 4.1 | 15.0 ± 3.2 | 14.4 ± 4.6 | |||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahimi, M.R.; Semenova, E.A.; Larin, A.K.; Kulemin, N.A.; Generozov, E.V.; Łubkowska, B.; Ahmetov, I.I.; Golpasandi, H. The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake. Nutrients 2023, 15, 1634. https://doi.org/10.3390/nu15071634
Rahimi MR, Semenova EA, Larin AK, Kulemin NA, Generozov EV, Łubkowska B, Ahmetov II, Golpasandi H. The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake. Nutrients. 2023; 15(7):1634. https://doi.org/10.3390/nu15071634
Chicago/Turabian StyleRahimi, Mohammad Rahman, Ekaterina A. Semenova, Andrey K. Larin, Nikolay A. Kulemin, Edward V. Generozov, Beata Łubkowska, Ildus I. Ahmetov, and Hadi Golpasandi. 2023. "The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake" Nutrients 15, no. 7: 1634. https://doi.org/10.3390/nu15071634
APA StyleRahimi, M. R., Semenova, E. A., Larin, A. K., Kulemin, N. A., Generozov, E. V., Łubkowska, B., Ahmetov, I. I., & Golpasandi, H. (2023). The ADORA2A TT Genotype Is Associated with Anti-Inflammatory Effects of Caffeine in Response to Resistance Exercise and Habitual Coffee Intake. Nutrients, 15(7), 1634. https://doi.org/10.3390/nu15071634