
Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Materials
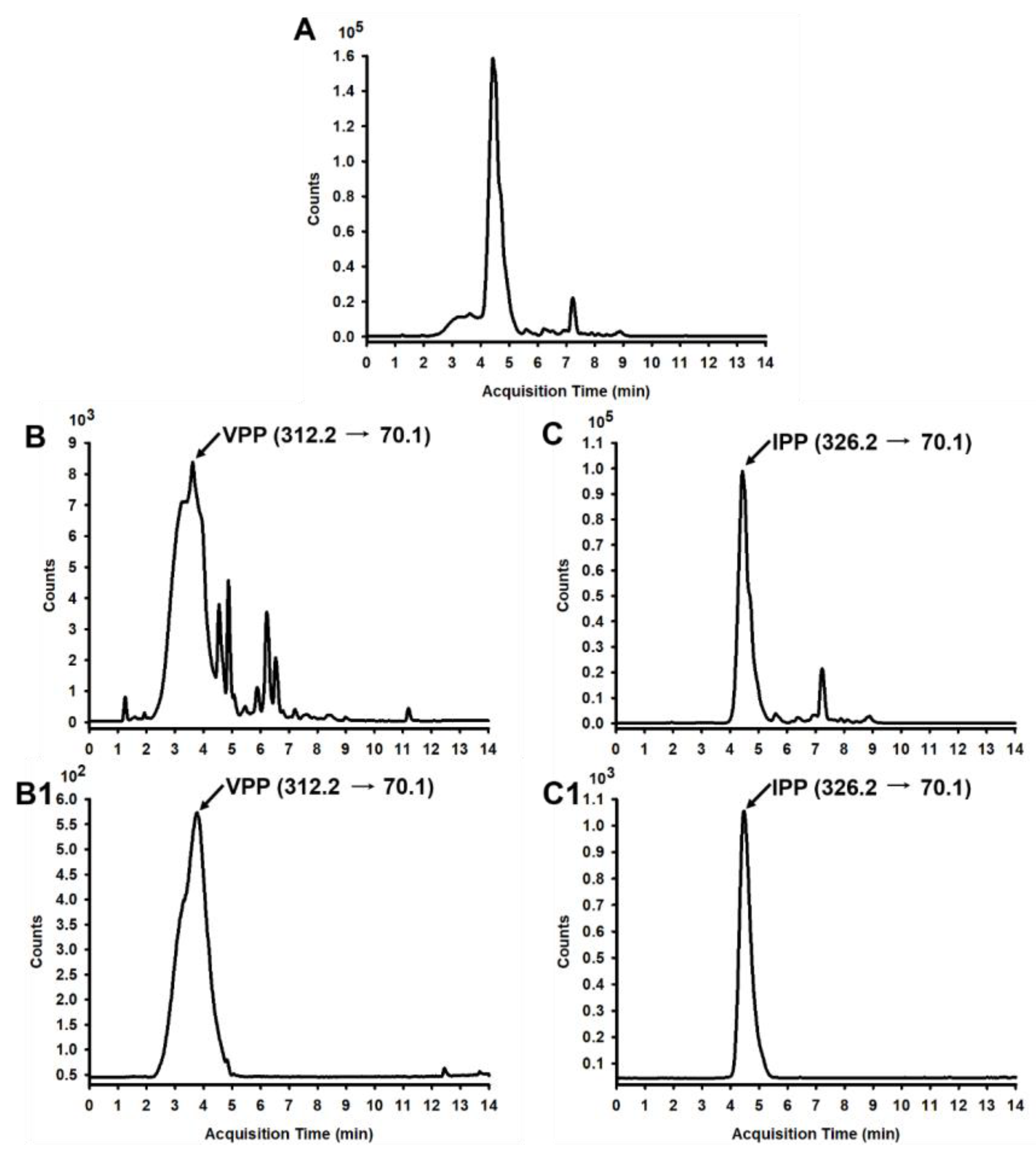
2.2. Preparation of CH and Determination of VPP and IPP
2.3. Animal Experiments and Diet
2.4. Oral Glucose Tolerance Test (OGTT) and Intraperitoneal Insulin Tolerance Test (IPITT)
2.5. Plasma Glucose, Insulin, Lipids, and Homeostasis Model Assessment of Insulin Resistance (HOMA-IR)
2.6. Cell Culture and Adipocyte Inflammation Induction
2.7. Western Blot Analysis
2.8. Morphological and Histopathological Analysis of Liver and Epidydimal Adipose Tissue
2.9. Statistical Analysis
3. Results
3.1. Detection and Quantification of Antihypertensive Tripeptides in Casein Hydrolysates
3.2. Effect of CH Containing Antihypertensive Peptides on the Physiological Parameters of HFD-Induced Obesity Mice
3.3. CH Ameliorated HFD-Induced Insulin Intolerance and Hyperinsulinemia in HFD-Fed C57BL/6J Mice
3.4. Effects of CH Containing Antihypertensive Peptides on the Plasma Lipid Metabolism Parameters of HFD-Induced Obese Mice
3.5. CH Alleviated Chronic Inflammation in HFD-Induced Obese Mice
3.6. Effect of CH Supplementation on the Epididymal Adipose and Liver Tissues in HFD-Induced Obese Mice
3.7. CH Improved the Adipocyte Dysfunction Induced by TNF-α
3.8. Effects of CH on the NF-κB and MAPK Signal Pathway in Impaired 3T3-L1 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- NCD Risk Factor Collaboration. Trends in adult body-mass index in 200 countries from 1975 to 2014: A pooled analysis of 1698 population-based measurement studies with 19·2 million participants. Lancet 2016, 387, 1377–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, M.; Zatterale, F.; Naderi, J.; Parrillo, L.; Formisano, P.; Raciti, G.A.; Beguinot, F.; Miele, C. Adipose tissue dysfunction as determinant of obesity-associated metabolic complications. Int. J. Mol. Sci. 2019, 20, 2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalupahana, N.S.; Moustaid-Moussa, N.; Claycombe, K.J. Immunity as a link between obesity and insulin resistance. Mol. Asp. Med. 2012, 33, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Haase, J.; Weyer, U.; Immig, K.; Klöting, N.; Blüher, M.; Eilers, J.; Bechmann, I.; Gericke, M. Local proliferation of macrophages in adipose tissue during obesity-induced inflammation. Diabetologia 2014, 57, 562–571. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Autieri, M.V.; Scalia, R. Adipose tissue inflammation and metabolic dysfunction in obesity. Am. J. Physiol. Cell Physiol. 2021, 320, c375–c391. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef]
- Serra, M.C.; Ryan, A.S.; Goldberg, A.P. Reduced LPL and subcutaneous lipid storage capacity are associated with metabolic syndrome in postmenopausal women with obesity. Obes. Sci. Pract. 2017, 3, 106–114. [Google Scholar] [CrossRef]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef]
- Banerjee, S.; Talukdar, I.; Banerjee, A.; Gupta, A.; Balaji, A.; Aduri, R. Type II diabetes mellitus and obesity: Common links, existing therapeutics and future developments. J. Biosci. 2019, 44, 150. [Google Scholar] [CrossRef]
- Healy, N.P.; Kirwan, A.M.; McArdle, M.A.; Holohan, K.; Nongonierma, A.B.; Keane, D.; Kelly, S.; Celkova, L.; Lyons, C.L.; McGillicuddy, F.C.; et al. A casein hydrolysate protects mice against high fat diet induced hyperglycemia by attenuating NLRP3 inflammasome-mediated inflammation and improving insulin signaling. Mol. Nutr. Food Res. 2016, 60, 2421–2432. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Weiss, M.; Gonzalez de Mejia, E. Anthocyanins from purple corn ameliorated tumor necrosis factor-α-induced inflammation and insulin resistance in 3T3-L1 adipocytes via activation of insulin signaling and enhanced GLUT4 translocation. Mol. Nutr. Food Res. 2017, 61, 1700362. [Google Scholar] [CrossRef] [PubMed]
- Rebollo-Hernanz, M.; Zhang, Q.; Aguilera, Y.; Martín-Cabrejas, M.A.; de Mejia, E.G. Cocoa shell aqueous phenolic extract preserves mitochondrial function and insulin sensitivity by attenuating inflammation between macrophages and adipocytes in vitro. Mol. Nutr. Food Res. 2019, 63, e1801413. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Ni, Y.; Nagata, N.; Zhuge, F.; Xu, L.; Nagashimada, M.; Yamamoto, S.; Ushida, Y.; Fuke, N.; Suganuma, H.; et al. Lycopene alleviates obesity-induced inflammation and insulin resistance by regulating M1/M2 status of macrophages. Mol. Nutr. Food Res. 2019, 63, e1900602. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, P.; Zhang, Y.; Shou, Q.; Li, H.; Zhu, Y.; He, L.; Chen, J.; Jiao, J. Eicosapentaenoic and docosahexaenoic acids differentially alter gut microbiome and reverse high-fat diet-induced insulin resistance. Mol. Nutr. Food Res. 2020, 64, e1900946. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Aluko, R.E. Food protein-derived bioactive peptides: Production, processing, and potential health benefits. J. Food Sci. 2012, 77, R11–R24. [Google Scholar] [CrossRef]
- Lemes, A.C.; Sala, L.; Ores Jda, C.; Braga, A.R.; Egea, M.B.; Fernandes, K.F. A review of the latest advances in encrypted bioactive peptides from protein-rich waste. Int. J. Mol. Sci. 2016, 17, 950. [Google Scholar] [CrossRef] [Green Version]
- Marcone, S.; Belton, O.; Fitzgerald, D.J. Milk-derived bioactive peptides and their health promoting effects: A potential role in atherosclerosis. Br. J. Clin. Pharmacol. 2017, 83, 152–162. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Song, J.; Du, M.; Mao, X. Bovine α-lactalbumin hydrolysates (α-LAH) ameliorate adipose insulin resistance and inflammation in high-fat diet-fed C57BL/6J Mice. Nutrients 2018, 10, 242. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Okubo, A.; Yamazaki, S.; Takano, T. Purification and characterization of angiotensin I-converting enzyme inhibitors from sour milk. J. Dairy Sci. 1995, 78, 777–783. [Google Scholar] [CrossRef]
- Sipola, M.; Finckenberg, P.; Korpela, R.; Vapaatalo, H.; Nurminen, M.L. Effect of long-term intake of milk products on blood pressure in hypertensive rats. J. Dairy Res. 2002, 69, 103–111. [Google Scholar] [CrossRef]
- Aihara, K.; Kajimoto, O.; Hirata, H.; Takahashi, R.; Nakamura, Y. Effect of powdered fermented milk with Lactobacillus helveticus on subjects with high-normal blood pressure or mild hypertension. J. Am. Coll. Nutr. 2005, 24, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Bu, T.; Zheng, J.; Liu, L.; He, G.; Wu, J. Preparation, bioavailability, and mechanism of emerging activities of Ile-Pro-Pro and Val-Pro-Pro. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1097–1110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boelsma, E.; Kloek, J. Lactotripeptides and antihypertensive effects: A critical review. Br. J. Nutr. 2009, 101, 776–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, T.; Ohki, K.; Kawagishi, R.; Kajimoto, Y.; Mizuno, S.; Nakamura, Y.; Kitakaze, M. Casein hydrolysate containing the antihypertensive tripeptides Val-Pro-Pro and Ile-Pro-Pro improves vascular endothelial function independent of blood pressure-lowering effects: Contribution of the inhibitory action of angiotensin-converting enzyme. Hypertens. Res. 2007, 30, 489–496. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, D.; Kassan, M.; Contreras Mdel, M.; Carrón, R.; Recio, I.; Montero, M.J.; Sevilla, M. Long-term intake of a milk casein hydrolysate attenuates the development of hypertension and involves cardiovascular benefits. Pharmacol. Res. 2011, 63, 398–404. [Google Scholar] [CrossRef]
- Aihara, K.; Osaka, M.; Yoshida, M. Oral administration of the milk casein-derived tripeptide Val-Pro-Pro attenuates high-fat diet-induced adipose tissue inflammation in mice. Br. J. Nutr. 2014, 112, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Sakamoto, Y.; Toh, M.; Ohara, N.; Hatanaka, Y.; Naka, A.; Kishimoto, Y.; Kondo, K.; Iida, K. Milk-derived peptide Val-Pro-Pro (VPP) inhibits obesity-induced adipose inflammation via an angiotensin-converting enzyme (ACE) dependent cascade. Mol. Nutr. Food Res. 2015, 59, 2502–2510. [Google Scholar] [CrossRef]
- Liu, L.; Zheng, J.; Zhou, M.; Li, S.; He, G.; Wu, J. Peptide analogues of VPP and IPP with improved glucose uptake activity in L6 myotubes can be released from cereal proteins. J. Agric. Food Chem. 2021, 69, 2875–2883. [Google Scholar] [CrossRef]
- Sun, X.; Zhao, H.; Liu, Z.; Sun, X.; Zhang, D.; Wang, S.; Xu, Y.; Zhang, G.; Wang, D. Modulation of gut microbiota by fucoxanthin during alleviation of obesity in high-fat diet-fed mice. J. Agric. Food Chem. 2020, 68, 5118–5128. [Google Scholar] [CrossRef]
- Zebisch, K.; Voigt, V.; Wabitsch, M.; Brandsch, M. Protocol for effective differentiation of 3T3-L1 cells to adipocytes. Anal. Biochem. 2012, 425, 88–90. [Google Scholar] [CrossRef]
- Mizuno, S.; Nishimura, S.; Matsuura, K.; Gotou, T.; Yamamoto, N. Release of short and proline-rich antihypertensive peptides from casein hydrolysate with an Aspergillus oryzae protease. J. Dairy Sci. 2004, 87, 3183–3188. [Google Scholar] [CrossRef] [Green Version]
- Gregor, M.F.; Hotamisligil, G.S. Inflammatory mechanisms in obesity. Annu. Rev. Immunol. 2011, 29, 415–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, S.; Wu, J. Milk-derived tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) promote adipocyte differentiation and inhibit inflammation in 3T3-F442A cells. PLoS ONE 2015, 10, e0117492. [Google Scholar] [CrossRef] [Green Version]
- Chakrabarti, S.; Jahandideh, F.; Davidge, S.T.; Wu, J. Milk-Derived Tripeptides IPP (Ile-Pro-Pro) and VPP (Val-Pro-Pro) Enhance Insulin Sensitivity and Prevent Insulin Resistance in 3T3-F442A Preadipocytes. J. Agric. Food Chem. 2018, 66, 10179–10187. [Google Scholar] [CrossRef] [PubMed]
- Matsuura, K.; Mennear, J.H.; Maeno, M.; Bernard, B.K. Studies of the toxicological potential of tripeptides (L-valyl-L-prolyl-L-proline and L-isoleucyl-L-prolyl-L-proline): VII. Micronucleus test of tripeptides-containing casein hydrolysate and Lactobacillus helveticus-fermented milk powders in rats and mice. Int. J. Toxicol. 2005, 24 (Suppl. 4), 91–96. [Google Scholar] [CrossRef] [PubMed]
- Jahandideh, F.; Zani, S.C.C.; Son, M.; Proctor, S.D.; Davidge, S.T.; Chan, C.B.; Wu, J. Egg white hydrolysate enhances insulin sensitivity in high-fat diet-induced insulin-resistant rats via Akt activation. Br. J. Nutr. 2019, 122, 14–24. [Google Scholar] [CrossRef]
- Liu, J.; Li, Y.; Yang, P.; Wan, J.; Chang, Q.; Wang, T.T.Y.; Lu, W.; Zhang, Y.; Wang, Q.; Yu, L.L. Gypenosides reduced the risk of overweight and insulin resistance in C57BL/6J mice through modulating adipose thermogenesis and gut microbiota. J. Agric. Food Chem. 2017, 65, 9237–9246. [Google Scholar] [CrossRef] [Green Version]
- Coppack, S.W. Pro-inflammatory cytokines and adipose tissue. Proc. Nutr. Soc. 2001, 60, 349–356. [Google Scholar] [CrossRef] [Green Version]
- Hagita, S.; Osaka, M.; Shimokado, K.; Yoshida, M. Adipose inflammation initiates recruitment of leukocytes to mouse femoral artery: Role of adipo-vascular axis in chronic inflammation. PLoS ONE 2011, 6, e19871. [Google Scholar] [CrossRef] [Green Version]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vascul. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef]
- Bays, H.E.; González-Campoy, J.M.; Bray, G.A.; Kitabchi, A.E.; Bergman, D.A.; Schorr, A.B.; Rodbard, H.W.; Henry, R.R. Pathogenic potential of adipose tissue and metabolic consequences of adipocyte hypertrophy and increased visceral adiposity. Expert Rev. Cardiovasc. Ther. 2008, 6, 343–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.W.; Liu, K.L.; Lin, Y.R.; Kuo, W.C. The mechanisms of carnosic acid attenuates tumor necrosis factor-α-mediated inflammation and insulin resistance in 3T3-L1 adipocytes. Mol. Nutr. Food Res. 2014, 58, 654–664. [Google Scholar] [CrossRef] [PubMed]
- Lafontan, M. Adipose tissue and adipocyte dysregulation. Diabetes Metab. 2014, 40, 16–28. [Google Scholar] [CrossRef]
- Yim, M.J.; Hosokawa, M.; Mizushina, Y.; Yoshida, H.; Saito, Y.; Miyashita, K. Suppressive effects of Amarouciaxanthin A on 3T3-L1 adipocyte differentiation through down-regulation of PPARγ and C/EBPα mRNA expression. J. Agric. Food Chem. 2011, 59, 1646–1652. [Google Scholar] [CrossRef]
- Majumder, K.; Mine, Y.; Wu, J. The potential of food protein-derived anti-inflammatory peptides against various chronic inflammatory diseases. J. Sci. Food Agric. 2016, 96, 2303–2311. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 92, 689–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, K.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Structure and activity study of egg protein ovotransferrin derived peptides (IRW and IQW) on endothelial inflammatory response and oxidative stress. J. Agric. Food Chem. 2013, 61, 2120–2129. [Google Scholar] [CrossRef] [PubMed]
- Herlaar, E.; Brown, Z. p38 MAPK signalling cascades in inflammatory disease. Mol. Med. Today 1999, 5, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Masuda, O.; Nakamura, Y.; Takano, T. Antihypertensive peptides are present in aorta after oral administration of sour milk containing these peptides to spontaneously hypertensive rats. J. Nutr. 1996, 126, 3063–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, K.; Nakamura, T.; Kamiie, J.; Takahashi, T.; Yamamoto, N. Accumulation of ACE inhibitory tripeptides, Val-Pro-Pro and Ile-Pro-Pro, in vascular endothelial cells. Biosci. Biotechnol. Biochem. 2012, 76, 1792–1795. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Intake, Body Weight, Organ Weight | Chow Diet | HFD | HFD + 4% CH |
|---|---|---|---|
| Initial body weight (g) | 27.05 ± 2.00 a | 31.77 ± 2.89 b | 30.96 ± 2.55 b |
| Final body weight (g) | 31.03 ± 2.98 a | 35.29 ± 4.60 b | 35.67 ± 3.28 b |
| Body weight change (g) | 3.26 ± 1.35 a | 4.04 ± 2.41 b | 4.97 ± 2.05 b |
| Food intake (g) | 3.42 ± 0.32 a | 2.36 ± 0.17 b | 2.32 ± 0.17 b |
| Energy intake (g) | 12.69 ± 1.20 a | 12.37 ± 0.89 a | 12.17 ± 0.92 a |
| Organ weight (g) | |||
| Liver | 0.99 ± 0.11 a | 0.98 ± 0.14 a | 1.03 ± 0.15 a |
| Kidney | 0.32 ± 0.03 a | 0.36 ± 0.03 a | 0.34 ± 0.02 a |
| Muscle | 0.35 ± 0.10 a | 0.38 ± 0.09 a | 0.37 ± 0.05 a |
| Epididymal fat | 0.74 ± 0.21 a | 1.28 ± 0.44 b | 1.24 ± 0.42 b |
| Mesenteric fat | 0.23 ± 0.11 a | 0.27 ± 0.13 a | 0.30 ± 0.11 a |
| Brown fat | 0.12 ± 0.05 a | 0.14 ± 0.06 a | 0.12 ± 0.04 a |
| Subcutaneous fat | 0.35 ± 0.11 a | 0.76 ± 0.54 b | 0.75 ± 0.38 b |
| Perinephric fat | 0.23 ± 0.08 a | 0.48 ± 0.21 b | 0.49 ± 0.22 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Yu, S.; Bu, T.; He, G.; Li, S.; Wu, J. Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway. Nutrients 2023, 15, 1813. https://doi.org/10.3390/nu15081813
Liu L, Yu S, Bu T, He G, Li S, Wu J. Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway. Nutrients. 2023; 15(8):1813. https://doi.org/10.3390/nu15081813
Chicago/Turabian StyleLiu, Ling, Songfeng Yu, Tingting Bu, Guoqing He, Shanshan Li, and Jianping Wu. 2023. "Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway" Nutrients 15, no. 8: 1813. https://doi.org/10.3390/nu15081813
APA StyleLiu, L., Yu, S., Bu, T., He, G., Li, S., & Wu, J. (2023). Casein Hydrolysate Alleviates Adipose Chronic Inflammation in High Fat-Diet Induced Obese C57BL/6J Mice through MAPK Pathway. Nutrients, 15(8), 1813. https://doi.org/10.3390/nu15081813