1. Introduction
Polyphenols are found in foods and beverages such as coffee, red wine, tea, and certain fruits and vegetables [
1]. They have antioxidant properties [
1,
2,
3] and may have beneficial effects on human health. Dark chocolate is rich in cacao polyphenols, which contain abundant epicatechins, catechins, and their oligomeric procyanidins, and are associated with the improvement in endothelial vascular function [
4,
5,
6]. The association between cacao polyphenols and cognitive function has also been investigated. Some behavioral studies have shown that a single ingestion of cacao polyphenols improves cognitive performance [
7,
8], inhibits increased subjective fatigue [
8], and helps to maintain concentration and mental stability during long-lasting cognitive tasks [
9]. Chronic intake of cacao polyphenols has also been reported to improve cognitive performance in terms of verbal learning, memory, and attention in young adults [
10].
Cacao polyphenols have also been reported to improve blood flow in both the peripheral and central nervous systems and to enhance brain activity related to cognitive performance. A previous study reported increased cerebral blood volume in the hippocampal dentate gyrus after 3 months of continuous high-cacao polyphenol food consumption in older adults [
11]. The study also showed an improvement in memory performance compared with that when consuming a low-cacao polyphenol food [
11]. In another study, a single dose of a cacao polyphenol-enriched cocoa drink resulted in increased central nervous system blood flow, with a peak increase in blood flow at 2 h after ingestion and a return to baseline at 6 h [
12]. Some neuroimaging studies have shown that brain activity relevant to memory and/or attention tasks, particularly in the prefrontal and parietal regions, is improved by consuming high cacao flavanols sub-chronically (for approximately 1 week) [
12,
13]. A more recent study reported that acute cacao polyphenol consumption improved reaction time in the Flanker test and increased brain activity in the supramarginal and inferior frontal gyri in individuals with type 1 diabetes and matched controls [
14]. Another study has reported that dark chocolate intake decreased the delta and theta absolute power and increased the alpha and beta absolute power of electrical activity in most brain regions [
15]. These findings suggest that even a single ingestion of cacao polyphenol contributes to brain functions associated with cognitive tasks.
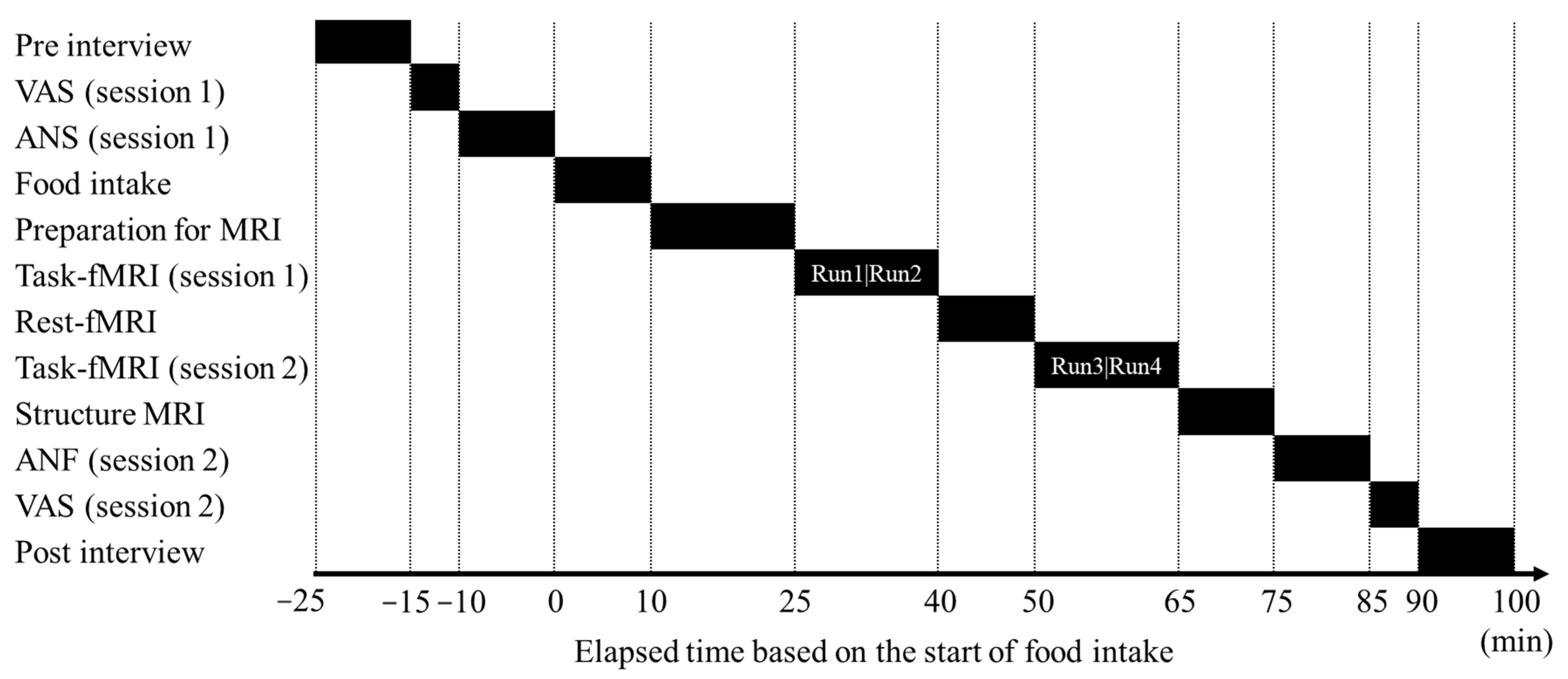
We have also previously investigated how an acute intake of chocolate with high cacao polyphenol content affects cognitive function during tasks that require high cognitive effort in healthy individuals. In that study, we used a “traffic light test” as a continuous and effortful task that demands high cognitive efforts with inhibition to a Stroop stimulus and selective attention to a disturbing stimulus, and we showed that a single intake of chocolate with a high concentration of cacao polyphenols (HC chocolate) helped maintain cognitive performance and concentration during continuous and effortful tasks compared with chocolate with a low concentrations of cacao polyphenols (LC chocolate) [
16]. However, it was unclear whether the acute intake of HC chocolate increases brain activity efficiency during continuous and effortful tasks. In this study, we measured brain activity during a similar traffic light test and examined whether the difference in brain activity between early and late time points after ingestion depended on the cacao polyphenol concentration.
4. Discussion
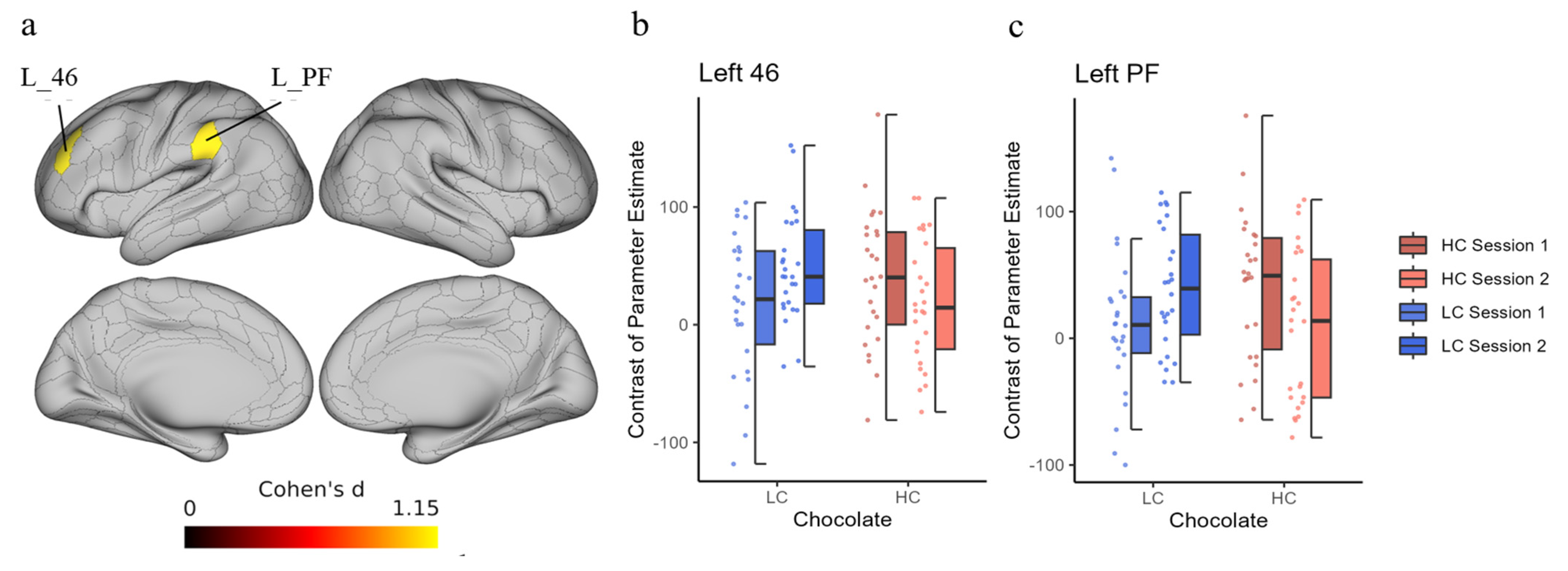
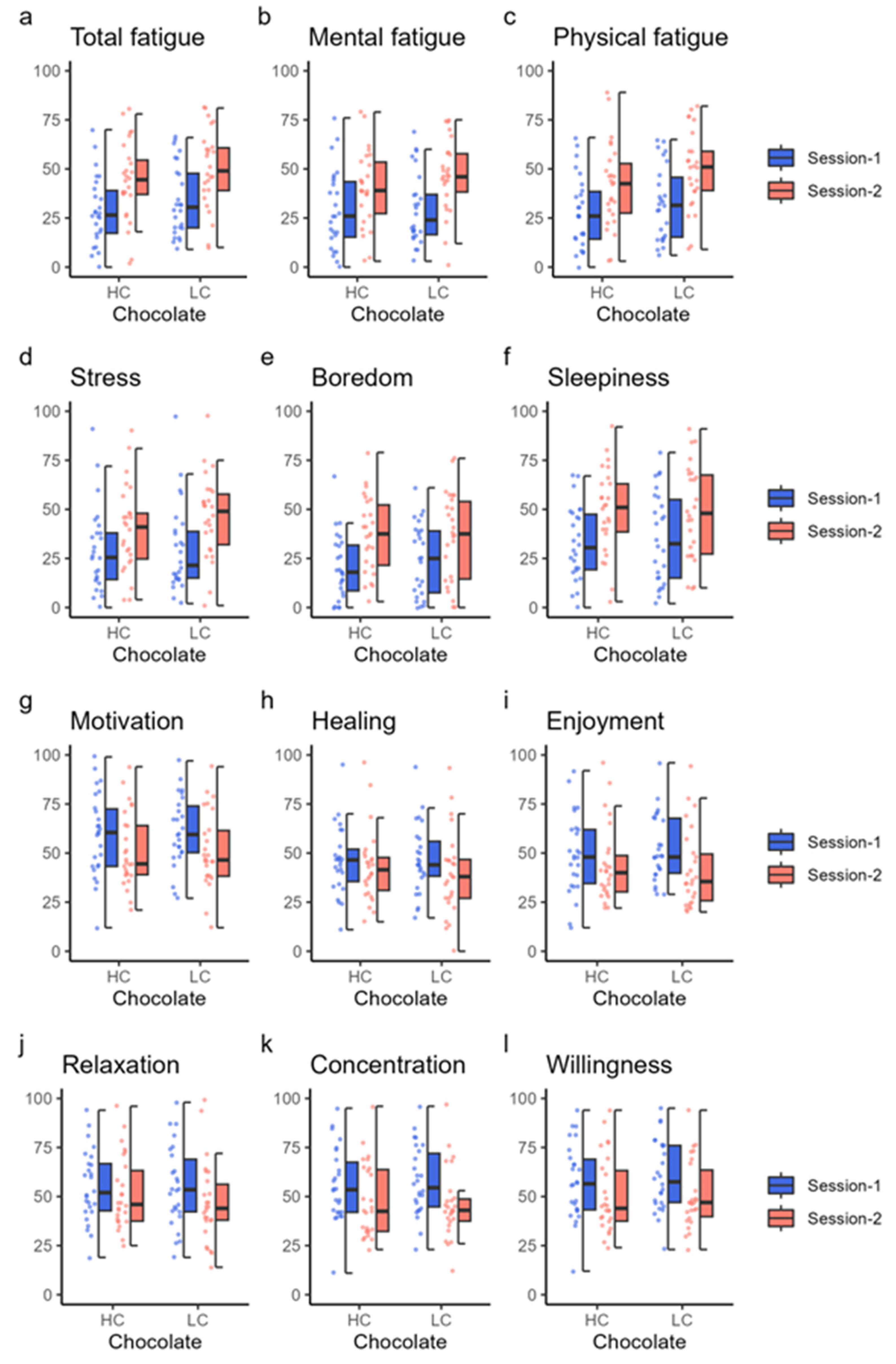
In this fMRI study, we observed that a single intake of HC chocolate enhanced the efficient use of cognitive resources to reduce the cost of brain activity during continuous and effortful tasks. When comparing brain activities during two consecutive cognitive task sessions with a short break in between, significant differences were observed between the HC and LC chocolate intake conditions in the differences in activation between session 1 and session 2 in the left BA46 (DLPFFC) and left BA40 (PF) ROIs. This indicates that brain activity in session 2 when HC chocolate was ingested was lower than that in session 2 when LC chocolate was ingested, although there were no significant changes in cognitive task performance.
Several recent studies have reported that cognitive efficiency can be increased by reducing the consumption of cognitive resources. For example, professional soccer players who play physical sports are reported to efficiently control their foot movements by significantly reducing the consumption of cognitive resources in the motor cortex of the brain [
44]. Another study reported decreased brain activity in the DLPFC during attentional tasks after cognitive training [
45]. Additionally, many previous fMRI studies have shown activation with less BOLD effects in quite discrete and task-dependent brain regions after acquiring skills in instrument [
46] and computer game performance [
47]. Thus, long-term training and upskilling result in energy-saving effects on neural activities. In this context, the novel finding of our study was that the cognitive resource-saving effect of HC chocolate intake was observed only 50–65 min after consumption.
In this study and a recent behavioral study [
16], we used a traffic light task that requires high cognitive effort by combining a Stroop task [
31] and a selective attention task. Stroop trials typically require attention to less salient stimulus attributes while ignoring more salient or automatically processed stimulus attributes. According to a meta-analysis by Cieslik et al. [
48], the Stroop task primarily involves brain regions within the frontoparietal executive control network. Another study suggested that cognitive conflict due to color/word mismatch in Stroop trials activates the conflict monitoring in the anterior cingulate gyrus, which in turn activates executive control of the DLPFC to resolve the conflict. DLPFC activation may lead to increased attention in subsequent trials, resulting in better performance [
49]. Moreover, the traffic light task requires additional executive control because selective attention is needed to suppress attention to interfering stimuli that are irrelevant to the task. When multiple tasks are simultaneously performed, each task’s performance decreases compared with when a single assignment is performed. This decrease may be caused by interference from other tasks that are simultaneously performed (dual-task interference effect), which reduces the cognitive resources directed toward the performance of the target task (task performance effect). Mizuno et al. [
50] found that dual tasks increase parieto-frontal associations compared with single tasks, suggesting their association with demands for cognitive resources. Additionally, several studies have demonstrated a link between cognitive load/demand and brain activity, even in non-dual tasks, indicating that increased cognitive load in working memory tasks decreases performance but increases activity in brain regions associated with working memory [
51]. Furthermore, cortical responses indicate that the same brain regions “work harder” rather than an increased network of brain regions [
52]. Due to these specific features of the cognitive task, rather than a heavier load, we observed a difference in behavioral and fMRI data between HC and LC chocolate intake.
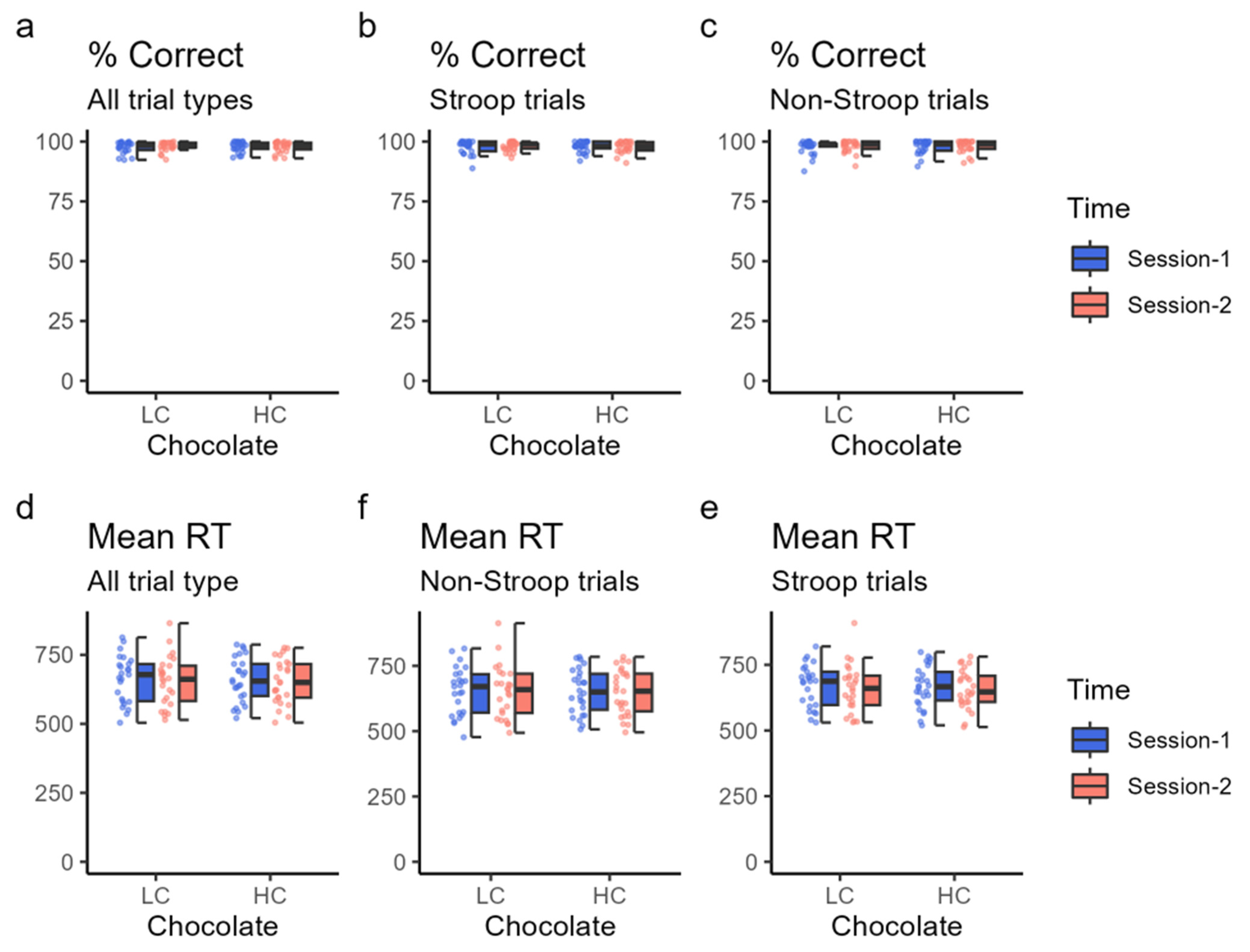
In this study that focused on task-related performance, LC chocolate intake decreased the overall correct response rates and correct response rates in Stroop trials in session 2 compared with session 1; however, HC chocolate intake did not have such a significant decrease in session 2, suggesting that HC chocolate consumption helps maintain performance and concentration for continuous and effortful cognitive tasks. The difference between the results from the behavioral and fMRI studies, especially for the decline of correct response rates in the second half of the LC chocolate intake group, may be explained by the different postures [
53] during the studies: the sitting and supine positions in the behavioral and fMRI studies, respectively, and head restraint [
54] in the fMRI study. Both factors are known to change regional cerebral blood flow and therefore alter task-related activation in the brain. Nevertheless, both results strengthened the effects of HC chocolate intake on brain function efficiency related to dual-cognitive tasks.
From a mechanistic point of view, we contemplated the reasons for HC chocolate mainly exhibiting such beneficial effects on neuronal activity- and energy consumption-saving and whether their actions derived from the effect on regional cerebral blood flow and synaptic plasticity, or a change in the efficiency of mitochondrial electron transport. Regarding the activation of brain regions, Magnore et al. [
55] reviewed the activation of brain regions relevant to chocolate from various perspectives. Chocolate consumption stimulates various brain areas, especially chemosensory areas, including the insula, prefrontal region, and caudomedial and caudolateral orbitofrontal cortices [
56]. fMRI studies have reported that the insular, human gustatory, and orbitofrontal cortices are central regions associated with taste [
57,
58]. Similarly, chocolate color modulates brain activity with a significant reduction in theta wave activity. This implies reduced attention and greater distraction [
59]. Additionally, the sight of chocolate generated greater activation in chocolate cravers than in non-cravers in the medial orbitofrontal cortex and ventral striatum [
60]. However, none of this evidence explains our results for the HC chocolate effect in the regional cognitive task-related brain activity-saving.
This study had several limitations. We emphasized the intake of high amounts of cacao polyphenols in chocolate formulations and designed a comparative study based on the dosage. In a systematic review, the effective dose of cacao polyphenols for a variety of health promotions was reported with cacao flavanol contents at approximately 500–900 mg/day [
18]. However, the Japanese Ministry of Health, Labor, and Welfare recommends <200 kcal for snacks per day, and we could not recommend that consumers take more than 25 g of chocolate daily as a snack. Consequently, we did not design such a high dose of cacao flavanols in this study (approximately 160 mg/25 g of chocolate measured by our study group). Moreover, we could not use white chocolate, which does not contain cacao polyphenols, for the control condition because this would have been a cause of concern in terms of blinding, and the taste and components of white chocolate could have affected the evaluation, especially since it is difficult to reproduce the unique taste (bitterness and astringency) of dark chocolate with white chocolate. Thus, while it could have been possible to visually control the appearance of the chocolate, the taste would still have significantly compromised blinding. Additionally, milk-derived ingredients of white chocolate, such as milk protein, are known to affect cognitive function and mood states [
61,
62]. Another limitation is that the contributions of other HC chocolate ingredients such as caffeine and theobromine remain unclear in isolation from those of higher cacao polyphenols levels. Nonetheless, this study provides further evidence for the benefits of practical amounts of HC chocolate for brain activity, at least when compared to those of ordinary chocolate, as a daily supplement or snack. In the near future, more robust effects may be demonstrated when an increased intake of flavanols or other effective pure polyphenol(s) is tested.


 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}