Oleuropein-Rich Jasminum Grandiflorum Flower Extract Regulates the LKB1-PGC-1α Axis Related to the Attenuation of Hepatocellular Lipid Dysmetabolism

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of OLE-JGF
2.2. Animals and Experimental Design
2.3. Cell and Treatment
2.4. Biochemical Analysis
2.5. Determination of TG in Liver Tissue and Cells
2.6. Histological Analysis
2.7. Western Blot
2.8. Quantitative RT-PCR
2.9. Determination of Mitochondrial DNA (mtDNA) Copy Numbers
2.10. Chromatin Immunoprecipitation (ChIP)
2.11. Statistical Analysis
3. Results
3.1. OLE-JGF Preparation
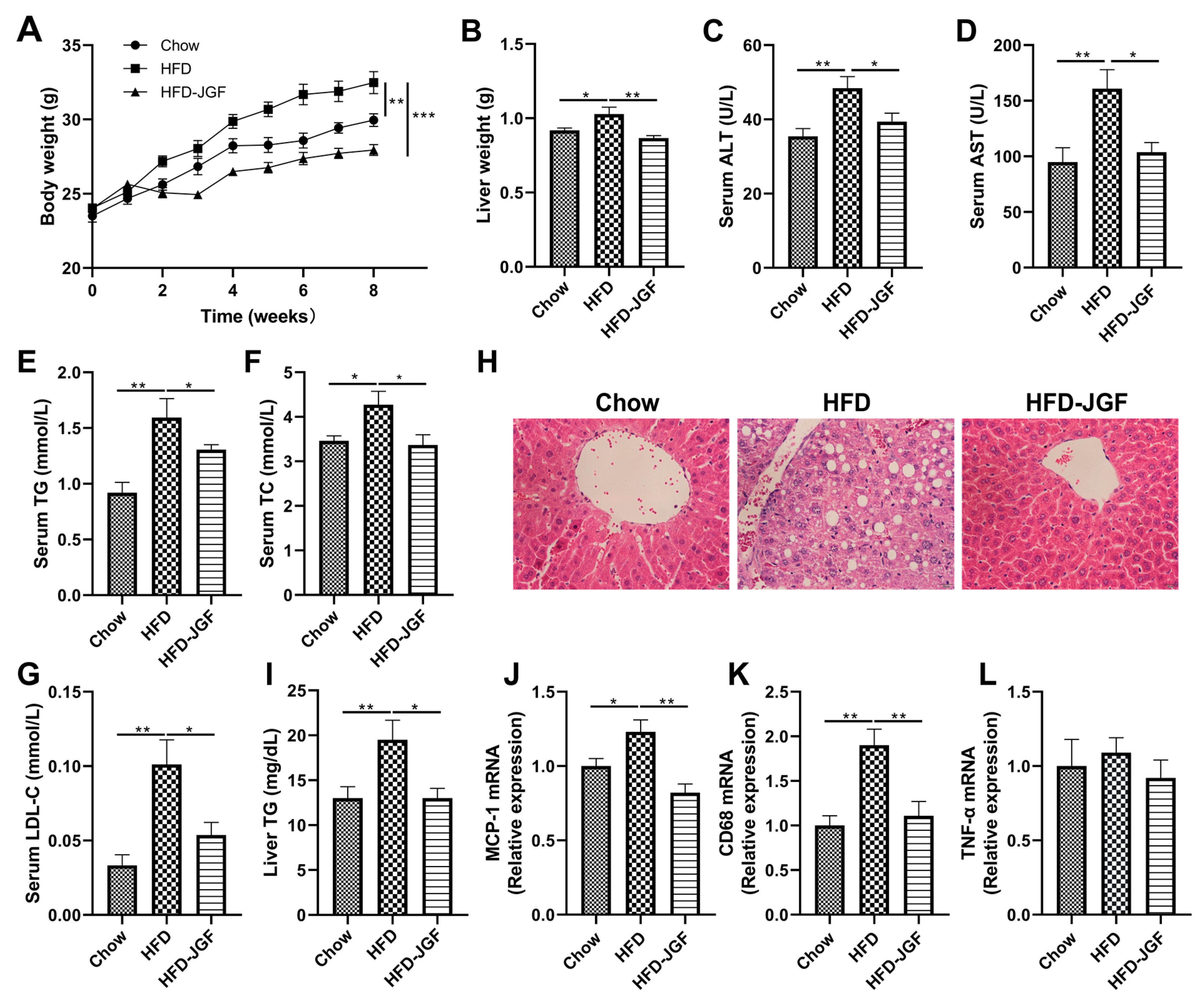
3.2. OLE-JGF Attenuated Liver Inflammation and Steatosis in HFD-Fed Mice
3.3. OLE-JGF Inhibited Hepatic Lipogenesis and Activated Mitochondrial Biogenesis in HFD-Fed Mice
3.4. OLE-JGF Activated Hepatic LKB1/PGC-1α Signaling Pathway in HFD-Fed Mice
3.5. OLE-JGF Inhibited OA-Induced Lipid Production in Liver Cells
3.6. OLE Stimulated PGC-1α Involved in Mitochondrial Biogenesis in OA-Induced Liver Cells
3.7. The Activation of LKB1-PGC-1α Pathway Participated in the Lipid-Lowering Effect of OLE in OA-Induced Liver Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Sun, X.; Yan, A.F.; Shi, Z.; Zhao, B.; Yan, N.; Li, K. Health consequences of obesity and projected future obesity health burden in China. Obesity 2022, 30, 1724–1751. [Google Scholar] [CrossRef] [PubMed]
- Grabner, G.F.; Xie, H.; Schweiger, M.; Zechner, R. Lipolysis: Cellular mechanisms for lipid mobilization from fat stores. Nat. Metab. 2021, 3, 1445–1465. [Google Scholar] [CrossRef] [PubMed]
- Ipsen, D.H.; Lykkesfeldt, J.; Tveden-Nyborg, P. Molecular mechanisms of hepatic lipid accumulation in non-alcoholic fatty liver disease. Cell Mol. Life Sci. 2018, 75, 3313–3327. [Google Scholar] [CrossRef] [PubMed]
- Yanovski, S.Z.; Yanovski, J.A. Long-term drug treatment for obesity: A systematic and clinical review. JAMA 2014, 311, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.L.; Zhang, Y.Z.; Wen, S.; Li, Q.H.; Chen, R.H.; Lai, X.F.; Zhang, Z.; Zhou, Z.; Xie, Y.; Zheng, X.; et al. Extract of Jasminum grandiflorum L. alleviates CCl4-induced liver injury by decreasing inflammation 2022, oxidative stress and hepatic CYP2E1 expression in mice. Biomed. Pharmacother. 2022, 152, 113255. [Google Scholar] [CrossRef] [PubMed]
- El-Shiekh, R.A.; Hussein, D.; Atta, A.H.; Mounier, S.M.; Mousa Shiekh, M.R.; Abdel-Sattar, E. Anti-inflammatory activity of Jasminum grandiflorum L. subsp. floribundum (Oleaceae) in inflammatory bowel disease and arthritis models. Biomed. Pharmacother. 2021, 140, 111770. [Google Scholar] [CrossRef] [PubMed]
- Arun, M.; Satish, S.; Anima, P. Phytopharmacological profile of Jasminum grandiflorum Linn. (Oleaceae). Chin. J. Integr. Med. 2016, 22, 311–320. [Google Scholar] [CrossRef]
- Li, D.L.; Tang, X.D.; Liu, C.; Li, H.F.; Li, S.Z.; Sun, S.L.; Zheng, X.; Wu, P.; Xu, X.; Zhang, K.; et al. Jasmine (Jasminum grandiflorum) flower extracts ameliorate tetradecanoylphorbol acetate induced ear edema in mice. Nat. Prod. Commun. 2020, 15, 1–7. [Google Scholar] [CrossRef]
- Zhang, Y.Z.; Sun, L.L.; Lai, X.F.; Peng, X.J.; Wen, S.; Zhang, Z.B.; Xie, Y.; Li, Q.; Chen, R.; Zheng, X.; et al. Gastroprotective effects of extract of Jasminum grandiflorum L. flower in HCl/EtOH-induced gastric mucosal ulceration mice. Biomed. Pharmacother. 2021, 144, 112268. [Google Scholar] [CrossRef]
- Tan, M.J.; Ye, J.M.; Turner, N.; Hohnen-Behrens, C.; Ke, C.Q.; Tang, C.P.; Chen, T.; Weiss, H.-C.; Gesing, E.-R.; Rowland, A.; et al. Antidiabetic activities of triterpenoids isolated from bitter melon associated with activation of the AMPK pathway. Chem. Biol. 2008, 15, 263–273. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, Y.Y.; Hu, C.M.; Wu, X.Z.; Lin, J.; Xiong, Z.; Zhang, K.; Xu, X.-T. Synthesis and biological evaluation of coumarin derivatives containing oxime ester as α-glucosidase inhibitors. Arab. J. Chem. 2022, 15, 104072. [Google Scholar] [CrossRef]
- Park, S.; Choi, Y.; Um, S.J.; Yoon, S.K.; Park, T. Oleuropein attenuates hepatic steatosis induced by high-fat diet in mice. J. Hepatol. 2011, 54, 984–993. [Google Scholar] [CrossRef] [PubMed]
- Hur, W.; Kim, S.W.; Lee, Y.K.; Choi, J.E.; Hong, S.W.; Song, M.J.; Bae, S.H.; Park, T.; Um, S.-J.; Yoon, S.K. Oleuropein reduces free fatty acid-induced lipogenesis via lowered extracellular signal-regulated kinase activation in hepatocytes. Nutr. Res. 2012, 32, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Hur, W.; Li, T.Z.; Lee, Y.K.; Choi, J.E.; Hong, S.W.; Lyoo, K.-S.; You, C.R.; Jung, E.S.; Jung, C.K.; et al. Oleuropein prevents the progression of steatohepatitis to hepatic fibrosis induced by a high-fat diet in mice. Exp. Mol. Med. 2014, 46, e92. [Google Scholar] [CrossRef] [PubMed]
- Ahamad, J.; Toufeeq, I.; Khan, M.A.; Ameen, M.S.M.; Anwer, E.T.; Uthirapathy, S.; Mir, S.R. Oleuropein: A natural antioxidant molecule in the treatment of metabolic syndrome. Phytother. Res. 2019, 33, 3112–3128. [Google Scholar] [CrossRef] [PubMed]
- Vezza, T.; Rodriguez-Nogales, A.; Algieri, F.; Garrido-Mesa, J.; Romero, M.; Sanchez, M.; Toral, M.; Martín-García, B.; Gómez-Caravaca, A.M.; Arráez-Román, D.; et al. The metabolic and vascular protective effects of olive (Olea europaea L.) leaf extract in diet-induced obesity in mice are related to the amelioration of gut microbiota dysbiosis and to its immunomodulatory properties. Pharmacol. Res. 2019, 150, 104487. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.M.; Liang, B.W.; Sun, J.P.; Li, J.Y.; Xiong, Z.; Wang, S.H.; Xuetao, X. Synthesis and biological evaluation of indole derivatives containing thiazolidine-2,4-dione as α-glucosidase inhibitors with antidiabetic activity. Eur. J. Med. Chem. 2023, 264, 115957. [Google Scholar] [CrossRef]
- Rao, Y.; Kuang, Z.Q.; Li, C.; Guo, S.Y.; Xu, Y.H.; Zhao, D.D.; Hu, Y.; Song, B.; Jiang, Z.; Ge, Z.; et al. Gut Akkermansia muciniphila ameliorates metabolic dysfunction-associated fatty liver disease by regulating the metabolism of L-aspartate via gut-liver axis. Gut Microbes 2021, 13, 1927633. [Google Scholar] [CrossRef]
- Zhang, S.P.; Zhu, P.; Yuan, J.N.; Cheng, K.M.; Xu, Q.X.; Chen, W.; Pan, Z.; Zheng, Y.Q. Non-alcoholic fatty liver disease combined with rheumatoid arthritis exacerbates liver fibrosis by stimulating co-localization of PTRF and TLR4 in rats. Front. Pharmacol. 2023, 14, 1149665. [Google Scholar] [CrossRef]
- Shuai, C.; Xia, G.Q.; Yuan, F.; Wang, S.; Lv, X.W. CD39-mediated ATP-adenosine signalling promotes hepatic stellate cell activation and alcoholic liver disease. Eur. J. Pharmacol. 2021, 905, 174198. [Google Scholar] [CrossRef]
- Qi, X.Y.; Zheng, S.G.; Ma, M.Y.; Lian, N.Q.; Wang, H.T.; Chen, L.R.; Song, A.P.; Lu, C.F.; Zheng, S.Z.; Jin, H.J. Curcumol suppresses CCF-mediated hepatocyte senescence through blocking LC3B–lamin B1 interaction in alcoholic fatty liver disease. Front. Pharmacol. 2022, 13, 912825. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.; Lu, Y.T.; Li, C.; Song, Q.Q.; Xu, Y.H.; Xu, Z.; Hu, Y.; Yu, H.; Gao, L.; Gu, L.; et al. Bouchardatine analogue alleviates non-alcoholic hepatic fatty liver disease/non-alcoholic steatohepatitis in high-fat fed mice by inhibiting ATP synthase activity. Br. J. Pharmacol. 2019, 176, 2877–2893. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.Z.; Liu, X.X.; Peng, C.; Tan, C.; Sun, H.L.; Liu, H.; Zhang, Y.; Wu, P.; Cui, C.; Liu, C.; et al. The phytochemical hyperforin triggers thermogenesis in adipose tissue via a Dlat-AMPK signaling axis to curb obesity. Cell Metab. 2021, 33, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Jeon, S.M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Li, Y.; Dai, Y.; Peng, J. Natural products for the treatment of type 2 diabetes mellitus: Pharmacology and mechanisms. Pharmacol. Res. 2018, 130, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zheng, Y.Q.; Tian, Y.Q.; Xu, Q.X.; Liu, S.C.; Li, H.H.; Cheng, K.M.; Yuan, J.N.; Liu, H.M.; Zhu, P. Astragalus polysaccharide alleviates alcoholic-induced hepatic fibrosis by inhibiting polymerase I and transcript release factor and the TLR4/JNK/NF-κB/MyD88 pathway. J. Ethnopharmacol. 2023, 314, 116662. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.Y.; Liu, L.N.; Xiang, R.X.; Bu, X.Y.; Qin, G.Z.; Dai, J.J.; Zhao, Z.G.; Fang, X.; Yang, S.; Han, J.; et al. Arctigenin mitigates insulin resistance by modulating the IRS2/GLUT4 pathway via TLR4 in type 2 diabetes mellitus mice. Int. Immunopharmacol. 2023, 114, 109529. [Google Scholar] [CrossRef]
- Hassen, I.; Casabianca, H.; Hosni, K. Biological activities of the natural antioxidant oleuropein: Exceeding the expectation—A mini-review. J. Funct. Foods 2015, 926–940. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Sayed Khan, A.; Khan, N.A.; Hichami, A. Antioxidant and anti-inflammatory potential of polyphenols contained in mediterranean diet in obesity: Molecular mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Domitrovic, R.; Jakovac, H.; Marchesi, V.V.; Sain, I.; Romic, Z.; Rahelic, D. Preventive and therapeutic effects of oleuropein against carbon tetrachloride-induced liver damage in mice. Pharmacol. Res. 2012, 65, 451–464. [Google Scholar] [CrossRef]
- Rocchetti, G.; Callegari, M.L.; Senizza, A.; Giuberti, G.; Ruzzolini, J.; Romani, A.; Urciuoli, S.; Nediani, C.; Lucini, L. Oleuropein from olive leaf extracts and extra-virgin olive oil provides distinctive phenolic profiles and modulation of microbiota in the large intestine. Food Chem. 2022, 380, 132187. [Google Scholar] [CrossRef] [PubMed]
- Viollet, B.; Foretz, M. LKB1/AMPK pathway in the control of hepatic energy metabolism. Signal Path Liver Dis. 2015, 23, 326–338. [Google Scholar] [CrossRef]
- Viollet, B.; Guigas, B.; Leclerc, J.; Hebrard, S.; Lantier, L.; Mounier, R.; Andreelli, F.; Foretz, M. AMP-activated protein kinase in the regulation of hepatic energy metabolism: From physiology to therapeutic perspectives. Acta Physiol. 2009, 196, 81–98. [Google Scholar] [CrossRef] [PubMed]
- Hadrich, F.; Mahmoudi, A.; Bouallagui, Z.; Feki, I.; Isoda, H.; Feve, B.; Sayadi, S. Evaluation of hypocholesterolemic effect of oleuropein in cholesterol-fed rats. Chem. Biol. Interact. 2016, 252, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Porcu, C.; Sideri, S.; Martini, M.; Cocomazzi, A.; Galli, A.; Tarantino, G. Oleuropein induces AMPK-dependent autophagy in NAFLD mice, regardless of the gender. Int. J. Mol. Sci. 2018, 19, 3948. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhao, H.; Wang, A. Oleuropein alleviates gestational diabetes mellitus by activating AMPK signaling. Endocr. Connect. 2021, 10, 45–53. [Google Scholar] [CrossRef]
- Hadrich, F.; Garcia, M.; Maalej, A.; Moldes, M.; Isoda, H.; Feve, B.; Sayadi, S. Oleuropein activated AMPK and induced insulin sensitivity in C2C12 muscle cells. Life Sci. 2016, 151, 167–173. [Google Scholar] [CrossRef]
- Patel, K.; Foretz, M.; Marion, A.; Campbell, D.G.; Gourlay, R.; Boudaba, N.; Tournier, E.; Titchenell, P.; Peggie, M.; Deak, M.; et al. The LKB1-salt-inducible kinase pathway functions as a key gluconeogenic suppressor in the liver. Nat. Commun. 2014, 5, 4535. [Google Scholar] [CrossRef]
- Shaw, R.J.; Lamia, K.A.; Vasquez, D.; Koo, S.H.; Bardeesy, N.; Depinho, R.A.; Montminy, M.; Cantley, L.C. The kinase LKB1 mediates glucose homeostasis in liver and therapeutic effects of metformin. Science 2005, 310, 1642–1646. [Google Scholar] [CrossRef]
- Wu, Z.X.; Xi, P.J.; Zhang, Y.; Wang, H.M.; Xue, J.; Sun, X.G.; Tian, D. LKB1 up-regulation inhibits hypothalamic inflammation and attenuates diet-induced obesity in mice. Metabolism 2021, 116, 154694. [Google Scholar] [CrossRef]
- Besse-Patin, A.; Leveille, M.; Oropeza, D.; Nguyen, B.N.; Prat, A.; Estall, J.L. Estrogen signals through peroxisome proliferator-activated receptor-gamma coactivator 1alpha to reduce oxidative damage associated with diet-induced fatty liver disease. Gastroenterology 2017, 152, 243–256. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.S.; Liu, J.Y.; Deng, H.; Ma, R.Y.; Liao, J.Y.; Liang, H.X.; Hu, J.; Li, J.; Guo, Z.; Cai, J.; et al. Targeting mitochondria-located circRNA SCAR alleviates NASH via reducing mROS output. Cell 2020, 183, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Jornayvaz, F.R.; Shulman, G.I. Regulation of mitochondrial biogenesis. Essays Biochem. 2010, 47, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Gureev, A.P.; Shaforostova, E.A.; Popov, V.N. Regulation of mitochondrial biogenesis as a way for active longevity: Interaction between the Nrf2 and PGC-1alpha signaling pathways. Front. Genet. 2019, 10, 435. [Google Scholar] [CrossRef]
- Yang, X.X.; Wang, X.; Shi, T.T.; Dong, J.C.; Li, F.J.; Zeng, L.X.; Yang, M.; Gu, W.; Li, J.-P.; Yu, J. Mitochondrial dysfunction in high-fat diet-induced nonalcoholic fatty liver disease: The alleviating effect and its mechanism of Polygonatum kingianum. Biomed. Pharmacother. 2019, 117, 109083. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hou, Y.; Zhao, X.; Wang, Y.; Li, Y.; Chen, C.; Zhou, X.; Jin, J.; Ye, J.; Li, D.; Gan, L.; et al. Oleuropein-Rich Jasminum Grandiflorum Flower Extract Regulates the LKB1-PGC-1α Axis Related to the Attenuation of Hepatocellular Lipid Dysmetabolism. Nutrients 2024, 16, 58. https://doi.org/10.3390/nu16010058
Hou Y, Zhao X, Wang Y, Li Y, Chen C, Zhou X, Jin J, Ye J, Li D, Gan L, et al. Oleuropein-Rich Jasminum Grandiflorum Flower Extract Regulates the LKB1-PGC-1α Axis Related to the Attenuation of Hepatocellular Lipid Dysmetabolism. Nutrients. 2024; 16(1):58. https://doi.org/10.3390/nu16010058
Chicago/Turabian StyleHou, Yajun, Xuan Zhao, Yalin Wang, Yapeng Li, Caihong Chen, Xiu Zhou, Jingwei Jin, Jiming Ye, Dongli Li, Lishe Gan, and et al. 2024. "Oleuropein-Rich Jasminum Grandiflorum Flower Extract Regulates the LKB1-PGC-1α Axis Related to the Attenuation of Hepatocellular Lipid Dysmetabolism" Nutrients 16, no. 1: 58. https://doi.org/10.3390/nu16010058
APA StyleHou, Y., Zhao, X., Wang, Y., Li, Y., Chen, C., Zhou, X., Jin, J., Ye, J., Li, D., Gan, L., & Wu, R. (2024). Oleuropein-Rich Jasminum Grandiflorum Flower Extract Regulates the LKB1-PGC-1α Axis Related to the Attenuation of Hepatocellular Lipid Dysmetabolism. Nutrients, 16(1), 58. https://doi.org/10.3390/nu16010058