
Omega-3 Supplementation and Its Effects on Osteoarthritis

Abstract
1. Introduction
2. Osteoarthritis
2.1. Prevalence
2.2. Genetics
2.3. Normal Cartilage
2.4. Osteoarthritic Cartilage
2.5. Joint Tissues Involved in Osteoarthritis
3. Inflammatory Process of Osteoarthritis
3.1. Pro-Inflammatory Cytokines
3.2. Pro-Inflammatory Chemokines
4. Anti-Inflammatory Process of Osteoarthritis
5. Fatty Acids
5.1. Omega-3 and Omega-6 Fatty Acids
5.2. Anti-Inflammatory Processes of Omega-3 PUFAs
6. Supplementation
6.1. Fatty Acid Composition: Ratio of Omega-6 to Omega-3
6.2. Omega-3 Supplementation and Synovial Fluid
6.3. Omega-3 Supplementation and Chondrocytes
6.4. Omega-3 Supplementation and Pain, Stiffness, and Physical Function
6.5. Omega-3 Supplementation for OA with Comorbidities
6.6. Omega-3 Supplementation and Its Indirect Effects on OA
6.7. Omega-3 Supplementation in Combination with Other Nutraceuticals in OA
6.8. Diet Rich in Omega-3 and Osteoarthritis
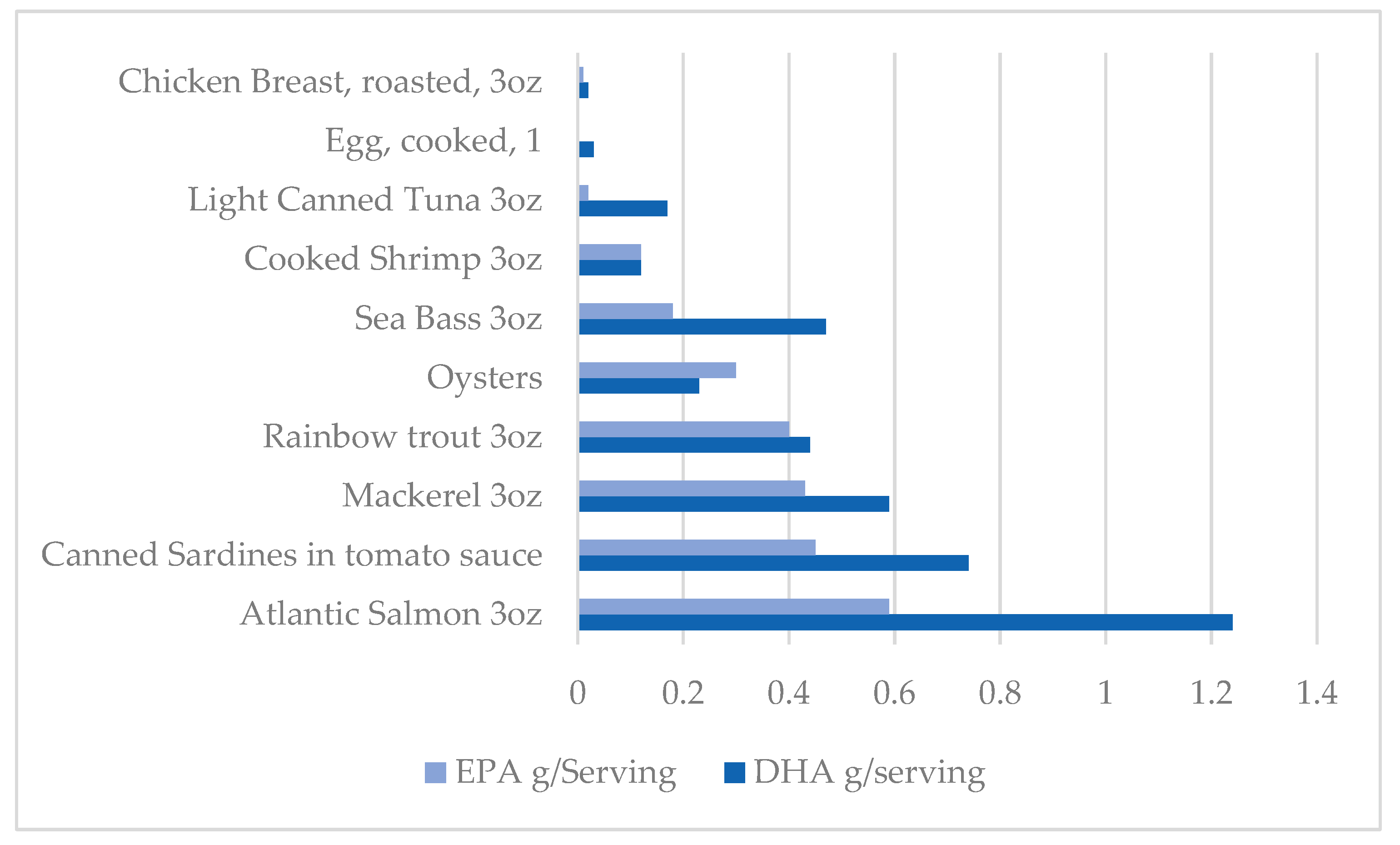
7. Dietary Sources of Omega-3
8. Limitations
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Martel-Pelletier, J.; Boileau, C.; Pelletier, J.; Roughley, P.J. Cartilage in normal and osteoarthritis conditions. Best. Pract. Res. Clin. Rheumatol. 2008, 22, 351–384. [Google Scholar] [CrossRef] [PubMed]
- Osteoarthritis. Available online: https://www.nia.nih.gov/health/osteoarthritis (accessed on 15 March 2024).
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef]
- Swain, S.; Sarmanova, A.; Coupland, C.; Doherty, M.; Zhang, W. Comorbidities in Osteoarthritis: A Systematic Review and Meta-Analysis of Observational Studies. Arthritis Care Res. 2020, 72, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Kluzek, S.; Newton, J.L.; Arden, N.K. Is osteoarthritis a metabolic disorder? Br. Med. Bull. 2015, 115, 111–121. [Google Scholar] [CrossRef]
- Colletti, A.; Cicero, A.F.G. Nutraceutical approach to chronic osteoarthritis: From molecular research to clinical evidence. Int. J. Mol. Sci. 2021, 22, 12920. [Google Scholar] [CrossRef]
- Cordingley, D.M.; Cornish, S.M. Omega-3 Fatty Acids for the Management of Osteoarthritis: A Narrative Review. Nutrients 2022, 14, 3362. [Google Scholar] [CrossRef]
- Quicke, J.G.; Conaghan, P.G.; Corp, N.; Peat, G. Osteoarthritis year in review 2021: Epidemiology & therapy. Osteoarthr. Cartil. 2022, 30, 196–206. [Google Scholar]
- Vina, E.; Kwoh, C. Epidemiology of osteoarthritis: Literature update. Curr. Opin. Rheumatol. 2018, 30, 160–167. [Google Scholar] [CrossRef]
- Felson, D.T. The epidemiology of knee osteoarthritis: Results from the framingham osteoarthritis study. Semin. Arthritis Rheum. 1990, 20, 42–50. [Google Scholar] [CrossRef]
- Dagenais, S.; Garbedian, S.; Wai, E.K. Systematic Review of the Prevalence of Radiographic Primary Hip Osteoarthritis. Clin. Orthop. Relat. Res. 2009, 467, 623–637. [Google Scholar] [CrossRef] [PubMed]
- Prieto-Alhambra, D.; Judge, A.; Javaid, M.K.; Cooper, C.; Diez-Perez, A.; Arden, N.K. Incidence and risk factors for clinically diagnosed knee, hip and hand osteoarthritis: Influences of age, gender and osteoarthritis affecting other joints. Ann. Rheum. Dis. 2014, 73, 1659–1664. [Google Scholar] [CrossRef] [PubMed]
- Boer, C.G.; Hatzikotoulas, K.; Southam, L.; Zhang, Y.; Coutinho de Almeida, R.; Wu, T.T.; Zheng, J.; Hartley, A.; Teder-Laving, M.; Terao, C.; et al. Deciphering osteoarthritis genetics across 826,690 individuals from 9 populations. Cell 2021, 184, 4784–4818. [Google Scholar] [CrossRef] [PubMed]
- Sacitharan; Korolchuk, V. I. Ageing and Osteoarthritis. Subcell. Biochem. 2019, 91, 123–159. [Google Scholar]
- Reynard, L.N.; Loughlin, J. Genetics and epigenetics of osteoarthritis. Maturitas 2012, 71, 200–204. [Google Scholar] [CrossRef]
- Yanagishita, M. Function of proteoglycans in the extracellular matrix. Acta Pathol. Jpn. 1993, 43, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Kohn, M.D.; Sassoon, A.A.; Fernando, N.D. Classifications in Brief: Kellgren-Lawrence Classification of Osteoarthritis. Clin. Orthop. Relat. Res. 2016, 474, 1886–1893. [Google Scholar] [CrossRef]
- Batushansky, A.; Lopes, E.B.P.; Zhu, S.; Humphries, K.M.; Griffin, T.M. GC–MS method for metabolic profiling of mouse femoral head articular cartilage reveals distinct effects of tissue culture and development. Osteoarthr. Cartil. 2019, 27, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, H.; Zhang, Z.; Yang, F.; Chen, J. Serum Metabolites as Potential Biomarkers for Diagnosis of Knee Osteoarthritis. Dis. Mak. 2015, 2015, 684794. [Google Scholar] [CrossRef]
- Zhang, W.; Likhodii, S.; Aref-Eshghi, E.; Zhang, Y.; Harper, P.E.; Randell, E.; Green, R.; Martin, G.; Furey, A.; Sun, G.; et al. Relationship between Blood Plasma and Synovial Fluid Metabolite Concentrations in Patients with Osteoarthritis. J. Rheumatol. 2015, 42, 859–865. [Google Scholar] [CrossRef]
- Batushansky, A.; Zhu, S.; Komaravolu, R.K.; South, S.; Mehta-D’souza, P.; Griffin, T.M. Fundamentals of OA. An initiative of Osteoarthritis and Cartilage. Obesity and metabolic factors in OA. Osteoarthr. Cartil. 2022, 30, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Cui, Z.; Urban, J.P.G. Factors influencing the oxygen concentration gradient from the synovial surface of articular cartilage to the cartilage–bone interface: A modeling study. Arthritis Rheum. 2004, 50, 3915–3924. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.; Svensson, C.I.; Etten, D.V.; Ghosh, S.S.; Murphy, A.N.; Powell, H.C.; Terkeltaub, R. Mediation of spontaneous knee osteoarthritis by progressive chondrocyte ATP depletion in Hartley guinea pigs. Arthritis Rheum. 2004, 50, 1216–1225. [Google Scholar] [CrossRef] [PubMed]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, Q.; Wang, X.; Kang, L.; Fu, Y.; Yin, Y.; Li, Z.; Liu, Y.; Xu, X.; Wang, Y. SOX9 is a regulator of ADAMTSs-induced cartilage degeneration at the early stage of human osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2259–2268. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T. The nuclear factor NF-kappaB pathway in inflammation. Cold Spring Harb. Perspect. Biol. 2009, 1, a001651. [Google Scholar] [CrossRef] [PubMed]
- Kalliolias, G.D.; Ivashkiv, L.B. TNF biology, pathogenic mechanisms and emerging therapeutic strategies. Nat. Rev. Rheumatol. 2016, 12, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Xu, X.; Zhang, Q.; Kang, L.; Xu, Y.; Zhang, K.; Li, L.; Liang, Y.; Hong, T.; Ye, Q.; et al. The IL-1β/AP-1/miR-30a/ADAMTS-5 axis regulates cartilage matrix degradation in human osteoarthritis. J. Mol. Med. 2016, 94, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α Signaling in Macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, Ł.A.; Szukiewicz, D. The Role of Inflammatory and Anti-Inflammatory Cytokines in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef]
- Scheller, J.; Chalaris, A.; Schmidt-Arras, D.; Rose-John, S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim. Biophys. Acta 2011, 1813, 878–888. [Google Scholar] [CrossRef]
- Li, L.; Li, Z.; Li, Y.; Hu, X.; Zhang, Y.; Fan, P. Profiling of inflammatory mediators in the synovial fluid related to pain in knee osteoarthritis. BMC Musculoskelet. Disord. 2020, 21, 99. [Google Scholar] [CrossRef]
- Raghu, H.; Lepus, C.M.; Wang, Q.; Wong, H.H.; Lingampalli, N.; Oliviero, F.; Punzi, L.; Giori, N.J.; Goodman, S.B.; Chu, C.R.; et al. CCL2/CCR2, but not CCL5/CCR5, mediates monocyte recruitment, inflammation and cartilage destruction in osteoarthritis. Ann. Rheum. Dis. 2017, 76, 914–922. [Google Scholar] [CrossRef]
- Alberts, B.; Johnson, A.; Lewis, J.; Morgan, D.; Raff, M.; Roberts, K.; Walter, P. Molecular Biology of the Cell, 6th ed.; International Student Edition; Garland Science, Taylor & Francis Group: New York, NY, USA, 2015. [Google Scholar]
- Scanzello, C.R. Chemokines and inflammation in osteoarthritis: Insights from patients and animal models. J. Orthop. Res. 2017, 35, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Monibi, F.; Roller, B.L.; Stoker, A.; Garner, B.; Bal, S.; Cook, J.L. Identification of Synovial Fluid Biomarkers for Knee Osteoarthritis and Correlation with Radiographic Assessment. J. Knee Surg. 2016, 29, 242–247. [Google Scholar] [CrossRef]
- Jordan, L.A.; Erlandsson, M.C.; Fenner, B.F.; Davies, R.; Harvey, A.K.; Choy, E.H.; Errington, R.; Bokarewa, M.I.; Williams, A.S. Inhibition of CCL3 abrogated precursor cell fusion and bone erosions in human osteoclast cultures and murine collagen-induced arthritis. Rheumatology 2018, 57, 2042–2052. [Google Scholar] [CrossRef] [PubMed]
- Koch, A.E.; Kunkel, S.L.; Shah, M.R.; Fu, R.; Mazarakis, D.D.; Haines, G.K.; Burdick, M.D.; Pope, R.M.; Strieter, R.M. Macrophage inflammatory protein-1β: A C-C chemokine in osteoarthritis. Clin. Immunol. Immunopathol. 1995, 77, 307–314. [Google Scholar] [CrossRef]
- Sandell, L.J.; Xing, X.; Franz, C.; Davies, S.; Chang, L.; Patra, D. Exuberant expression of chemokine genes by adult human articular chondrocytes in response to IL-1beta. Osteoarthr. Cartil. 2008, 16, 1560–1571. [Google Scholar] [CrossRef] [PubMed]
- García-Manrique, M.; Calvet, J.; Orellana, C.; Berenguer-Llergo, A.; Garcia-Cirera, S.; Llop, M.; Albiñana-Giménez, N.; Galisteo-Lencastre, C.; Gratacós, J. Synovial fluid but not plasma interleukin-8 is associated with clinical severity and inflammatory markers in knee osteoarthritis women with joint effusion. Sci. Rep. 2021, 11, 5258. [Google Scholar] [CrossRef]
- Chauffier, K.; Laiguillon, M.; Bougault, C.; Gosset, M.; Priam, S.; Salvat, C.; Mladenovic, Z.; Nourissat, G.; Jacques, C.; Houard, X.; et al. Induction of the chemokine IL-8/Kc by the articular cartilage: Possible influence on osteoarthritis. Jt. Bone Spine 2012, 79, 604–609. [Google Scholar] [CrossRef]
- Koh, S.M.; Chan, C.K.; Teo, S.H.; Singh, S.; Merican, A.; Ng, W.M.; Abbas, A.; Kamarul, T. Elevated plasma and synovial fluid interleukin-8 and interleukin-18 may be associated with the pathogenesis of knee osteoarthritis. Knee 2020, 27, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, W.; Bragg, R.; Elmansi, A.M.; McGee-Lawrence, M.E.; Isales, C.M.; Hamrick, M.W.; Hill, W.D.; Fulzele, S. Stromal cell-derived factor-1 (CXCL12) and its role in bone and muscle biology. Cytokine 2019, 123, 154783. [Google Scholar] [CrossRef]
- Guang, L.G.; Boskey, A.L.; Zhu, W. Regulatory role of stromal cell-derived factor-1 in bone morphogenetic protein-2-induced chondrogenic differentiation in vitro. Int. J. Biochem. Cell Biol. 2012, 44, 1825–1833. [Google Scholar] [CrossRef] [PubMed]
- Schlaak, J.F.; Pfers, I.; Meyer Zum Büschenfelde, K.; Märker-Hermann, E. Different cytokine profiles in the synovial fluid of patients with osteoarthritis, rheumatoid arthritis and seronegative spondylarthropathies. Clin. Exp. Rheumatol. 1996, 14, 155–162. [Google Scholar]
- Forster, T.; Chapman, K.; Loughlin, J. Common variants within the interleukin 4 receptor α gene (IL4R) are associated with susceptibility to osteoarthritis. Hum. Genet. 2004, 114, 391–395. [Google Scholar] [CrossRef] [PubMed]
- von Kaeppler, E.P.; Wang, Q.; Raghu, H.; Bloom, M.S.; Wong, H.; Robinson, W.H. Interleukin 4 promotes anti-inflammatory macrophages that clear cartilage debris and inhibits osteoclast development to protect against osteoarthritis. Clin. Immunol. 2021, 229, 108784. [Google Scholar] [CrossRef] [PubMed]
- Behrendt, P.; Häfelein, K.; Preusse-Prange, A.; Bayer, A.; Seekamp, A.; Kurz, B. IL-10 ameliorates TNF-α induced meniscus degeneration in mature meniscal tissue in vitro. BMC Musculoskelet. Disord. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.; Rogers, V.E.; Henriksen, V.T.; Trawick, R.H.; Momberger, N.G.; Lynn Rasmussen, G. Circulating IL-10 is compromised in patients predisposed to developing and in patients with severe knee osteoarthritis. Sci. Rep. 2021, 11, 1812. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Browne, H.; Mobasheri, A.; Rayman, M.P. What is the evidence for a role for diet and nutrition in osteoarthritis? Rheumatology 2018, 57, iv61–iv74. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Vauzour, D.; Tejera, N.; O’Neill, C.; Booz, V.; Jude, B.; Wolf, I.M.A.; Rigby, N.; Silvan, J.M.; Curtis, P.J.; Cassidy, A.; et al. Anthocyanins do not influence long-chain n-3 fatty acid status: Studies in cells, rodents and humans. J. Nutr. Biochem. 2015, 26, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Gibson, R.A.; Muhlhausler, B.; Makrides, M. Conversion of linoleic acid and alpha-linolenic acid to long-chain polyunsaturated fatty acids (LCPUFAs), with a focus on pregnancy, lactation and the first 2 years of life. Matern. Child. Nutr. 2011, 7, 17–26. [Google Scholar] [CrossRef]
- Mustonen, A.; Nieminen, P. Fatty Acids and Oxylipins in Osteoarthritis and Rheumatoid Arthritis—A Complex Field with Significant Potential for Future Treatments. Curr. Rheumatol. Rep. 2021, 23, 41. [Google Scholar] [CrossRef]
- Möller, I.; Rodas, G.; Villalón, J.M.; Rodas, J.A.; Angulo, F.; Martínez, N.; Vergés, J. Randomized, double-blind, placebo-controlled study to evaluate the effect of treatment with an SPMs-enriched oil on chronic pain and inflammation, functionality, and quality of life in patients with symptomatic knee osteoarthritis: GAUDI study. J. Transl. Med. 2023, 21, 423. [Google Scholar] [CrossRef]
- Liu, C.; Fan, D.; Lei, Q.; Lu, A.; He, X. Roles of Resolvins in Chronic Inflammatory Response. Int. J. Mol. Sci. 2022, 23, 14883. [Google Scholar] [CrossRef]
- Dravid, A.A.; Dhanabalan, K.; Agarwal, S.; Agarwal, R. Resolvin D1-loaded nanoliposomes promote M2 macrophage polarization and are effective in the treatment of osteoarthritis. Bioeng. Transl. Med. 2022, 7, e10281. [Google Scholar] [CrossRef]
- Wang, X.; Yang, M.; Yu, G.; Qi, J.; Jia, Q.; Liu, S.; Jiang, W.; Su, S.; Chi, Z.; Wang, R.; et al. Promoting the proliferation of osteoarthritis chondrocytes by resolvin D1 regulating the NLRP3/caspase-1 signaling pathway. Cell. Signal. 2024, 113, 110960. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Jiang, W.; Wang, X.; Du, S.; Qi, J.; Jia, Q.; Song, H. Resolvin D1 inhibits the proliferation of osteoarthritis fibroblast-like synoviocytes through the Hippo-YAP signaling pathway. BMC Musculoskelet. Disord. 2022, 23, 149. [Google Scholar] [CrossRef] [PubMed]
- Kariminezhad, Z.; Rahimi, M.; Fernandes, J.; Maltais, R.; Sancéau, J.; Poirier, D.; Fahmi, H.; Benderdour, M. Development of New Resolvin D1 Analogues for Osteoarthritis Therapy: Acellular and Computational Approaches to Study Their Antioxidant Activities. Antioxidants 2024, 13, 386. [Google Scholar] [CrossRef]
- Piao, S.; Du, W.; Wei, Y.; Yang, Y.; Feng, X.; Bai, L. Protectin DX attenuates IL-1β-induced inflammation via the AMPK/NF-κB pathway in chondrocytes and ameliorates osteoarthritis progression in a rat model. Int. Immunopharmacol. 2020, 78, 106043. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res. Rev. 2012, 11, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Feng, X.; Zhang, H.; Wei, Y.; Yang, Y.; Tian, Y.; Bai, L. Maresin-1 suppresses IL-1β-induced MMP-13 secretion by activating the PI3K/AKT pathway and inhibiting the NF-κB pathway in synovioblasts of an osteoarthritis rat model with treadmill exercise. Connect. Tissue Res. 2021, 62, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Oshimura, M.; Ito, H. PI3K-Akt pathway: Its functions and alterations in human cancer. Apoptosis 2004, 9, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Li, Y.; Tang, Y.; Ding, C.; Hunter, D.J. Pharmacotherapy for knee osteoarthritis: Current and emerging therapies. Expert. Opin. Pharmacother. 2020, 21, 797–809. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipids Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.; Driban, J.B.; Xu, C.; Lapane, K.L.; McAlindon, T.E.; Eaton, C.B. Dietary Fat Intake and Radiographic Progression of Knee Osteoarthritis: Data From the Osteoarthritis Initiative. Arthritis Care Res. 2017, 69, 368–375. [Google Scholar] [CrossRef] [PubMed]
- Jenniskens, Y.; Siawash, M.; van de Lest, C.; Verhaar, J.; Kloppenburg, M.; Zuurmond, A.M.; Stojanovic-Susulic, V.; van Osch, G.; Clockaerts, S. Monounsaturated and Saturated, but Not N-6 Polyunsaturated Fatty Acids Decrease Cartilage Destruction under Inflammatory Conditions: A Preliminary Study. Cartilage 2013, 4, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Dunn, D.M.; Henry, J.H.; Li, Y.; Watkins, B.A. Decreased production of inflammatory mediators in human osteoarthritic chondrocytes by conjugated linoleic acids. Lipids 2004, 39, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, Y.; Ma, L.; Meng, H.; Bai, X.; Fan, Z.; Yu, F.; Guo, A. A low ratio of n-6/n-3 polyunsaturated fatty acids suppresses matrix metalloproteinase 13 expression and reduces adjuvant-induced arthritis in rats. Nutr. Res. 2015, 35, 1113–1121. [Google Scholar] [CrossRef]
- Sibille, K.; King, C.; Garrett, T.; Glover, T.; Zhang, H.; Chen, H.; Reddy, D.; Goodin, B.; Sotolongo, A.; Petrov, M.; et al. Omega-6: Omega-3 PUFA Ratio, Pain, Functioning, and Distress in Adults With Knee Pain. Clin. J. Pain 2018, 34, 182–189. [Google Scholar] [CrossRef]
- Mustonen, A.; Käkelä, R.; Joukainen, A.; Lehenkari, P.; Jaroma, A.; Kääriäinen, T.; Kröger, H.; Paakkonen, T.; Sihvo, S.P.; Nieminen, P. Synovial Fluid Fatty Acid Profiles Are Differently Altered by Inflammatory Joint Pathologies in the Shoulder and Knee Joints. Biology 2021, 10, 401. [Google Scholar] [CrossRef] [PubMed]
- Baker, K.R.; Matthan, N.R.; Lichtenstein, A.H.; Niu, J.; Guermazi, A.; Roemer, F.; Grainger, A.; Nevitt, M.C.; Clancy, M.; Lewis, C.E.; et al. Association of plasma n-6 and n-3 polyunsaturated fatty acids with synovitis in the knee: The MOST study. Osteoarthr. Cartil. 2012, 20, 382–387. [Google Scholar] [CrossRef] [PubMed]
- Knott, L.; Avery, N.C.; Hollander, A.P.; Tarlton, J.F. Regulation of osteoarthritis by omega-3 (n-3) polyunsaturated fatty acids in a naturally occurring model of disease. Osteoarthr. Cart. 2011, 19, 1150–1157. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Zhang, L.; Yan, Z.; Li, Z.; Fu, M.; Xue, C.; Wang, J. A low proportion n-6/n-3 PUFA diet supplemented with Antarctic krill (Euphausia superba) oil protects against osteoarthritis by attenuating inflammation in ovariectomized mice. Food Funct. 2021, 12, 6766–6779. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Dai, Y.; Zhang, L.; Tian, Y.; Li, Z.; Wang, J. Effects of Edible Oils with Different n-6/n-3 PUFA Ratios on Articular Cartilage Degeneration via Regulating the NF-κB Signaling Pathway. J. Agric. Food Chem. 2020, 68, 12641–12650. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Gong, Z.; Wang, G.; Cao, R.; Yin, H.; Ma, L.; Guo, A. DHA attenuates cartilage degeneration by mediating apoptosis and autophagy in human chondrocytes and rat models of osteoarthritis. In Vitro Cell. Dev. Biol.-Anim. 2023, 59, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Hill, C.L.; March, L.M.; Aitken, D.; Lester, S.E.; Battersby, R.; Hynes, K.; Fedorova, T.; Proudman, S.M.; James, M.; Cleland, L.G.; et al. Fish oil in knee osteoarthritis: A randomised clinical trial of low dose versus high dose. Ann. Rheum. Dis. 2016, 75, 23–29. [Google Scholar] [CrossRef]
- Stonehouse, W.; Benassi-Evans, B.; Bednarz, J.; Vincent, A.D.; Hall, S.; Hill, C.L. Krill oil improved osteoarthritic knee pain in adults with mild to moderate knee osteoarthritis: A 6-month multicenter, randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2022, 116, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Brien, S.; Prescott, P.; Coghlan, B.; Bashir, N.; Lewith, G. Systematic review of the nutritional supplement Perna Canaliculus (green-lipped mussel) in the treatment of osteoarthritis. QJM 2008, 101, 167–179. [Google Scholar] [CrossRef]
- Stebbings, S.; Gray, A.; Schneiders, A.G.; Sansom, A. A randomized double-blind placebo-controlled trial to investigate the effectiveness and safety of a novel green-lipped mussel extract -BioLex® -for managing pain in moderate to severe osteoarthritis of the hip and knee. BMC Complement. Altern. Med. 2017, 17, 416. [Google Scholar] [CrossRef]
- Tsubosaka, M.; Kihara, S.; Hayashi, S.; Nagata, J.; Kuwahara, T.; Fujita, M.; Kikuchi, K.; Takashimi, Y.; Kamenaga, T.; Kuroda, Y.; et al. Gelatin hydrogels with eicosapentaenoic acid can prevent osteoarthritis progression in vivo in a mouse model. J. Orthop. Res. 2020, 38, 2157–2169. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Yokoyama, M.; Itakura, H.; Kita, T.; Kitabatake, A.; Nakaya, N.; Sakata, T.; Shimada, K.; Shirato, K.; Origasa, H.; et al. Effects of eicosapentaenoic acid on major coronary events in hypercholesterolaemic patients (JELIS): A randomised open-label, blinded endpoint analysis. Lancet 2007, 369, 1090–1098. [Google Scholar]
- Alfaddagh, A.; Elajami, T.K.; Saleh, M.; Elajami, M.; Bistrian, B.R.; Welty, F.K. The effect of eicosapentaenoic and docosahexaenoic acids on physical function, exercise, and joint replacement in patients with coronary artery disease: A secondary analysis of a randomized clinical trial. J. Clin. Lipidol. 2018, 12, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Moskowitz, R.W.; Nuki, G.; Abramson, S.; Altman, R.D.; Arden, N.; Bierma-Zeinstra, S.; Brandt, K.D.; Croft, P.; Doherty, M.; et al. OARSI recommendations for the management of hip and knee osteoarthritis, Part II: OARSI evidence-based, expert consensus guidelines. Osteoarthr. Cartil. 2008, 16, 137–162. [Google Scholar] [CrossRef]
- Smith, G.I.; Julliand, S.; Reeds, D.N.; Sinacore, D.R.; Klein, S.; Mittendorfer, B. Fish oil–derived n–3 PUFA therapy increases muscle mass and function in healthy older adults. Am. J. Clin. Nutr. 2015, 102, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.A.; de Oliveira, L.M.; Watanabe, S.H.; Jones, A.; Natour, J. Isokinetic assessment of the hip muscles in patients with osteoarthritis of the knee. Clinics 2010, 65, 1253–1259. [Google Scholar] [CrossRef]
- Tsuchiya, Y.; Yanagimoto, K.; Nakazato, K.; Hayamizu, K.; Ochi, E. Eicosapentaenoic and docosahexaenoic acids-rich fish oil supplementation attenuates strength loss and limited joint range of motion after eccentric contractions: A randomized, double-blind, placebo-controlled, parallel-group trial. Eur. J. Appl. Physiol. 2016, 116, 1179–1188. [Google Scholar] [CrossRef]
- Ochi, E.; Tsuchiya, Y.; Yanagimoto, K. Effect of eicosapentaenoic acids-rich fish oil supplementation on motor nerve function after eccentric contractions. Int. Soc. Sports Nutr. 2017, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, Y.; Ueda, H.; Yanagimoto, K.; Kato, A.; Ochi, E. 4-week eicosapentaenoic acid-rich fish oil supplementation partially protects muscular damage following eccentric contractions. J. Int. Soc. Sports Nutr. 2021, 18, 18. [Google Scholar] [CrossRef]
- Clayton, J.J. Nutraceuticals in the Management of Osteoarthritis. Orthopedics 2007, 30, 624–631. [Google Scholar]
- Jacquet, A.; Girodet, P.; Pariente, A.; Forest, K.; Mallet, L.; Moore, N. Phytalgic®, a food supplement, vs placebo in patients with osteoarthritis of the knee or hip: A randomised double-blind placebo-controlled clinical trial. Arthritis Res. Ther. 2009, 11, R192. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Ao, M.; Dong, B.; Jiang, Y.; Yu, L.; Chen, Z.; Hu, C.; Xu, R. Anti-Inflammatory Effects of Curcumin in the Inflammatory Diseases: Status, Limitations and Countermeasures. Drug Des. Devel Ther. 2021, 15, 4503–4525. [Google Scholar] [CrossRef] [PubMed]
- Kuszewski, J.C.; Wong, R.H.X.; Howe, P.R.C. Fish oil supplementation reduces osteoarthritis-specific pain in older adults with overweight/obesity. Rheumatol. Adv. Pract. 2020, 4, rkaa036. [Google Scholar] [CrossRef] [PubMed]
- Towheed, T.; Maxwell, L.; Anastassiades, T.P.; Shea, B.; Houpt, J.; Welch, V.; Hochberg, M.C.; Wells, G.A.; Towheed, T. Glucosamine therapy for treating osteoarthritis. Cochrane Database Syst. Rev. 2005, 2009, CD002946. [Google Scholar] [CrossRef] [PubMed]
- Gruenwald, J.; Petzold, E.; Busch, R.; Petzold, H.; Graubaum, H. Effect of glucosamine sulfate with or without omega-3 fatty acids in patients with osteoarthritis. Adv. Ther. 2009, 26, 858–871. [Google Scholar] [CrossRef] [PubMed]
- MacFarlane, L.A.; Cook, N.R.; Kim, E.; Lee, I.; Iversen, M.D.; Gordon, D.; Buring, J.E.; Katz, J.N.; Manson, J.E.; Costenbader, K.H. The Effects of Vitamin D and Marine Omega-3 Fatty Acid Supplementation on Chronic Knee Pain in Older US Adults: Results from a Randomized Trial. Arthritis Rheumatol. 2020, 72, 1836–1844. [Google Scholar] [CrossRef]
- Messier, S.P.; Beavers, D.P.; Queen, K.; Mihalko, S.L.; Miller, G.D.; Losina, E.; Katz, J.N.; Loeser, R.F.; DeVita, P.; Hunter, D.J.; et al. Effect of Diet and Exercise on Knee Pain in Patients with Osteoarthritis and Overweight or Obesity: A Randomized Clinical Trial. JAMA 2022, 328, 2242–2251. [Google Scholar] [CrossRef] [PubMed]
- Dolatkhah, N.; Toopchizadeh, V.; Barmaki, S.; Salekzamani, Y.; Najjari, A.; Farshbaf-Khalili, A.; Dolati, S. The effect of an anti-inflammatory in comparison with a low caloric diet on physical and mental health in overweight and obese women with knee osteoarthritis: A randomized clinical trial. Eur. J. Nutr. 2023, 62, 659–672. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, A.; Zarrinjooiee, G.; Mousavi, S.N.; Abdollahi Sabet, S.; Jalili, N. Effects of a Mediterranean Diet Compared with the Low-Fat Diet on Patients with Knee Osteoarthritis: A Randomized Feeding Trial. Int. J. Clin. Pract. 2022, 2022, 7275192. [Google Scholar] [CrossRef]
- Pitaraki, E.E. The role of Mediterranean diet and it’s components on the progress of osteoarthritis. J. Frailty Sarcopenia Falls 2017, 2, 45–52. [Google Scholar] [CrossRef]
- Dyer, J.; Davison, G.; Marcora, S.M.; Mauger, A.R. Effect of a Mediterranean type diet on inflammatory and cartilage degradation biomarkers in patients with osteoarthritis. J. Nutr. Health Aging 2017, 21, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Sharif, M.; Kirwan, J.; Charni, N.; Sandell, L.J.; Whittles, C.; Garnero, P. A 5-yr longitudinal study of type IIA collagen synthesis and total type II collagen degradation in patients with knee osteoarthritis-association with disease progression. Rheumatology 2007, 46, 938–943. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Office of Dietary Supplements—Omega-3 Fatty Acids. Internet. Available online: https://ods.od.nih.gov/factsheets/Omega3FattyAcids-HealthProfessional/ (accessed on 20 January 2024).
- Arterburn, L.M.; Oken, H.A.; Hall, B.E. Algal-oil capsules and cooked salmon: Nutritionally equivalent sources of docosahexaenoic acid. J. Am. Diet Assoc. 2008, 13, 260. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef]
- National Institute of Health—Office of Dietary Supplements. Omega-3 Fatty Acids Fact Sheet for Consumers. Internet. Available online: https://ods.od.nih.gov/pdf/factsheets/Omega3FattyAcids-Consumer.pdf (accessed on 15 March 2024).

{kind=link}
| Grade | Classification |
|---|---|
| 0 | No evidence of OA, normal radiographic findings |
| 1 | Doubtful narrowing of the joint space with possible osteophyte formation |
| 2 | Possible narrowing of joint space with definite osteophyte formation |
| 3 | Definite narrowing of joint space, moderate osteophyte formation, some sclerosis, and possible deformity of bony ends |
| 4 | Large osteophyte formation, severe narrowing of the joint space, with marked sclerosis and definite deformity of bone ends |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shawl, M.; Geetha, T.; Burnett, D.; Babu, J.R. Omega-3 Supplementation and Its Effects on Osteoarthritis. Nutrients 2024, 16, 1650. https://doi.org/10.3390/nu16111650
Shawl M, Geetha T, Burnett D, Babu JR. Omega-3 Supplementation and Its Effects on Osteoarthritis. Nutrients. 2024; 16(11):1650. https://doi.org/10.3390/nu16111650
Chicago/Turabian StyleShawl, Megan, Thangiah Geetha, Donna Burnett, and Jeganathan Ramesh Babu. 2024. "Omega-3 Supplementation and Its Effects on Osteoarthritis" Nutrients 16, no. 11: 1650. https://doi.org/10.3390/nu16111650
APA StyleShawl, M., Geetha, T., Burnett, D., & Babu, J. R. (2024). Omega-3 Supplementation and Its Effects on Osteoarthritis. Nutrients, 16(11), 1650. https://doi.org/10.3390/nu16111650