
A Symbiotic Meal Containing Extruded Sorghum and Probiotic (Bifidobacterium longum) Ameliorated Intestinal Health Markers in Individuals with Chronic Kidney Disease: A Secondary Analysis of a Subsample from a Previous Randomized and Controlled Clinical Trial
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
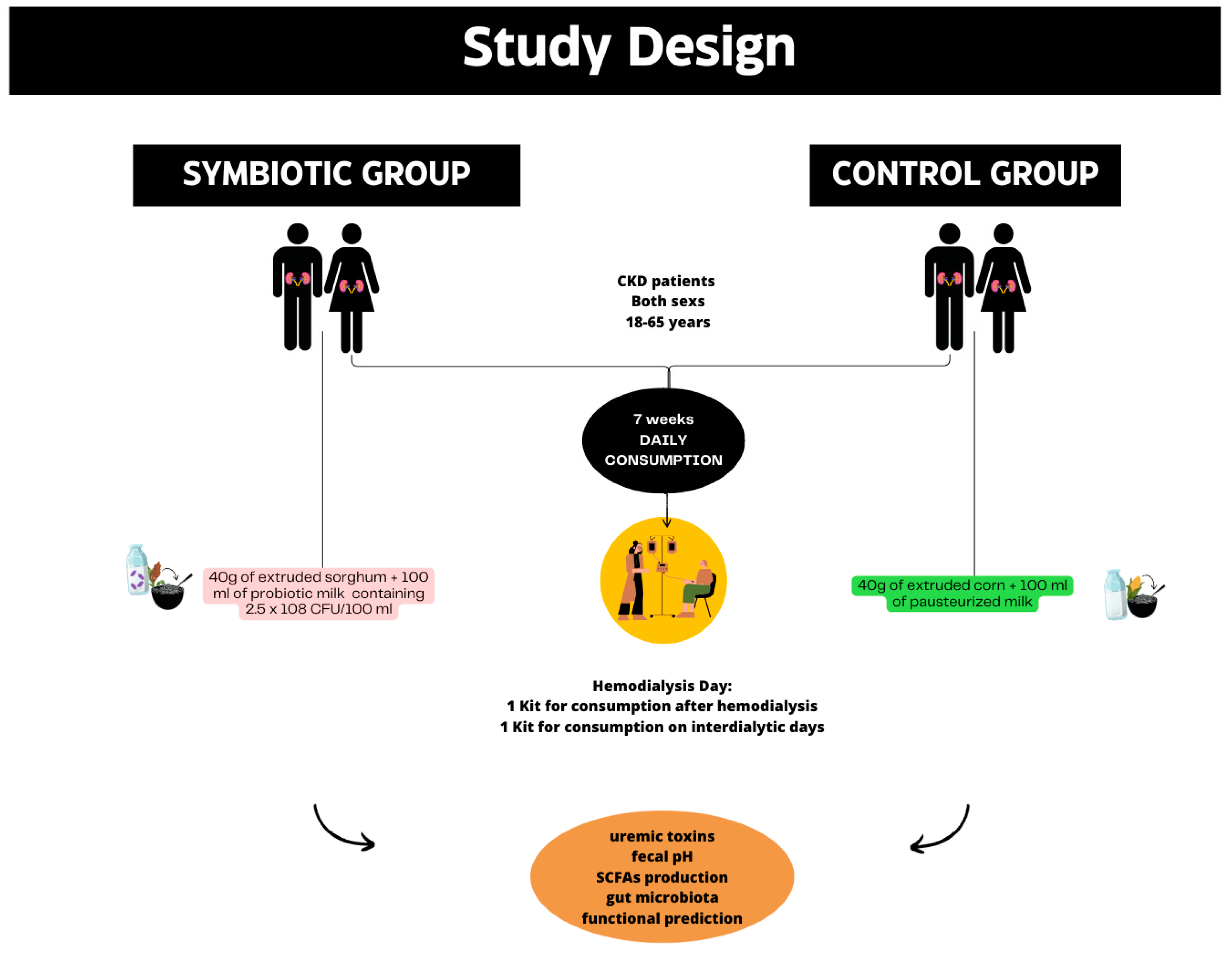
2.1. Study Design
2.2. Participants
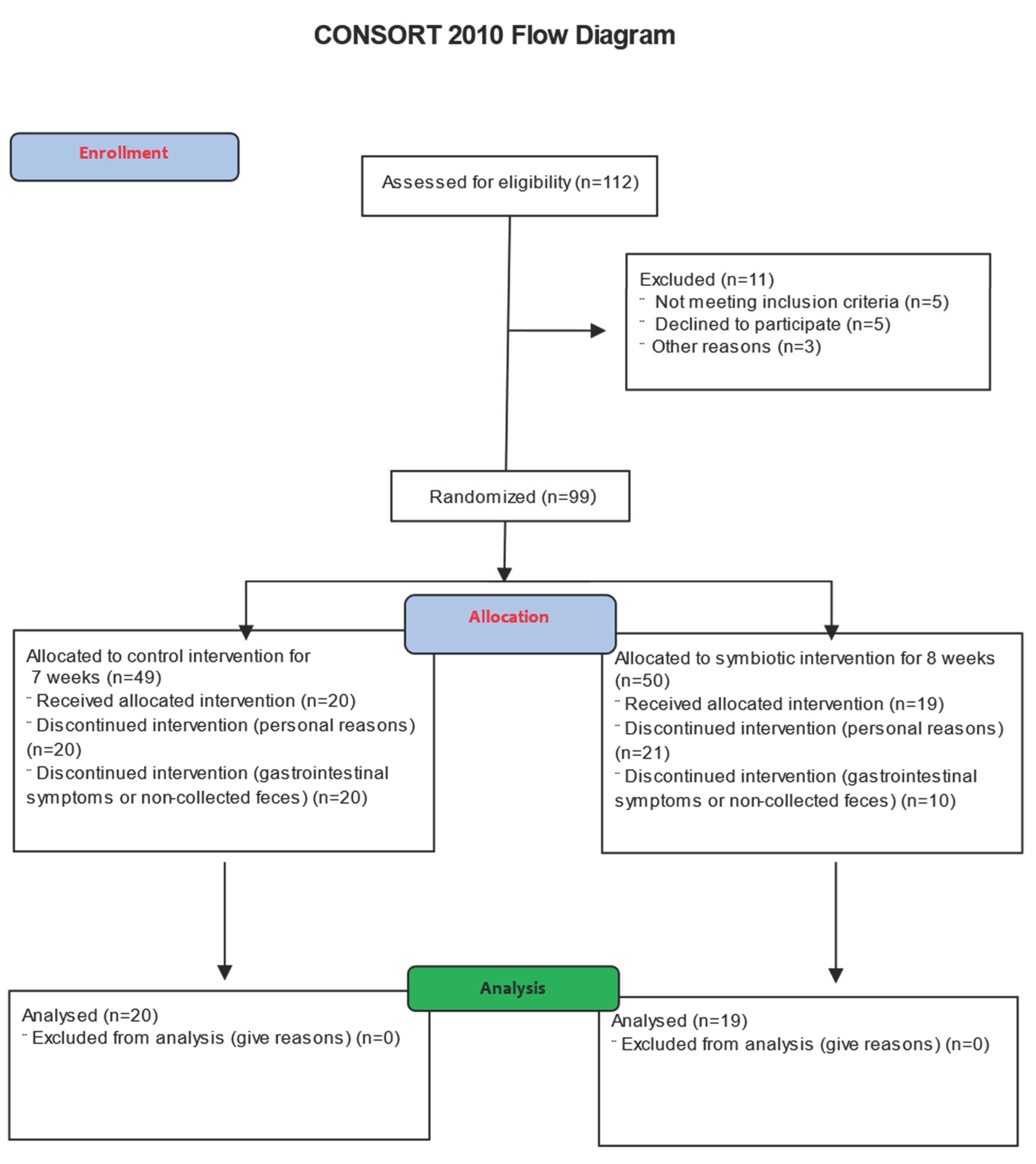
2.3. Randomization, Allocation, and Sample Power
2.4. Raw Material and Meal Preparation
2.5. Interventions
2.6. Outcomes
2.7. Anthropometric Measures
2.8. Analysis of the Consumption of Macronutrients
2.9. Uremic Markers
2.10. Gastrointestinal Symptoms
2.11. Fecal SCFA Concentrations
2.12. Analysis of Intestinal Microbiota
2.13. Statistical Analysis
3. Results
3.1. Baseline Characteristics of Treatment Groups
3.2. Consumption of Macronutrients and Body Mass Index
3.3. Uremic Markers
3.4. Gastrointestinal Symptoms
3.5. Fecal SCFA Concentrations
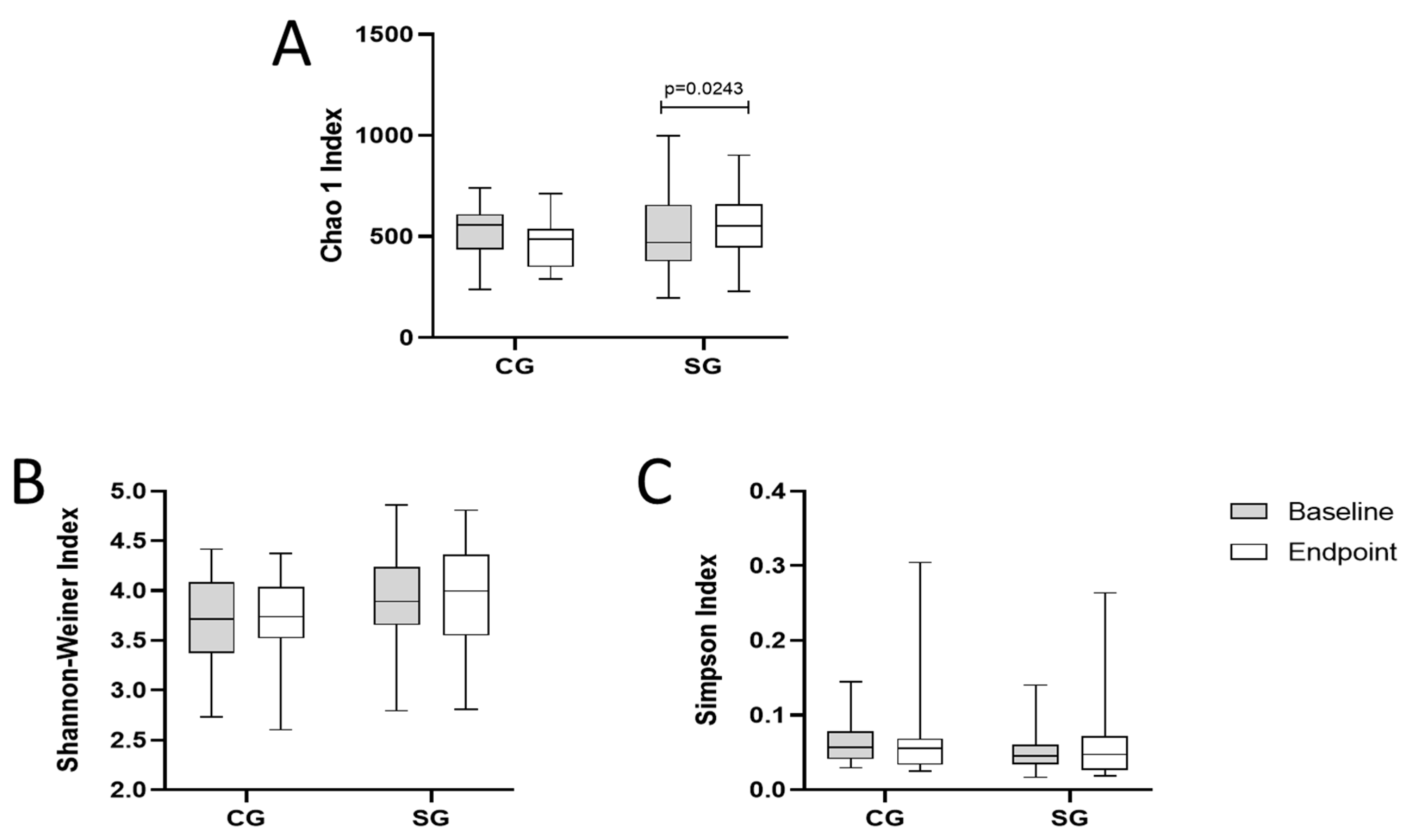
3.6. Analysis of Intestinal Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Charles, C.; Ferris, A.H. Chronic Kidney Disease. Prim. Care Clin. Off. Pract. 2020, 47, 585–595. [Google Scholar] [CrossRef]
- Bikbov, B.; Purcell, C.A.; Levey, A.S.; Smith, M.; Abdoli, A.; Abebe, M.; Adebayo, O.M.; Afarideh, M.; Agarwal, S.K.; Agudelo-Botero, M.; et al. Global, regional, and national burden of chronic kidney disease, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2020, 395, 709–733. [Google Scholar] [CrossRef] [PubMed]
- Poesen, R.; Viaene, L.; Verbeke, K.; Augustijns, P.; Bammens, B.; Claes, K.; Kuypers, D.; Evenepoel, P.; Meijers, B. Cardiovascular disease relates to intestinal uptake of p-cresol in patients with chronic kidney disease. BMC Nephrol. 2014, 15, 87. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of Urease- and Uricase-Containing, Indole- and p-Cresol-Forming and Contraction of Short-Chain Fatty Acid-Producing Intestinal Microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Chen, D.-Q.; Chen, L.; Liu, J.-R.; Vaziri, N.D.; Guo, Y.; Zhao, Y.-Y. Microbiome–metabolome reveals the contribution of gut–kidney axis on kidney disease 11 Medical and Health Sciences 1103 Clinical Sciences. J. Transl. Med. 2019, 17, 5. [Google Scholar] [CrossRef]
- Kanbay, M.; Onal, E.M.; Afsar, B.; Dagel, T.; Yerlikaya, A.; Covic, A.; Vaziri, N.D. The crosstalk of gut microbiota and chronic kidney disease: Role of inflammation, proteinuria, hypertension, and diabetes mellitus. Int. Urol. Nephrol. 2018, 50, 1453–1466. [Google Scholar] [CrossRef]
- Guldris, S.C.; Parra, E.G.; Amenós, A.C. Microbiota intestinal en la enfermedad renal crónica. Nefrologia 2017, 37, 9–19. [Google Scholar] [CrossRef]
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.d.C.S.O.; de Lima, S.L.S.; da Silva, B.P.; Toledo, R.C.L.; Moreira, M.E.d.C.; Anunciação, P.C.; Walter, E.H.M.; Carvalho, C.W.P.; Queiroz, V.A.V.; Ribeiro, A.Q.; et al. Evaluation of the health benefits of consumption of extruded tannin sorghum with unfermented probiotic milk in individuals with chronic kidney disease. Food Res. Int. 2018, 107, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Li, Y.; Marion, T.; Tong, Y.; Zaiss, M.M.; Tang, Z.; Zhang, Q.; Liu, Y.; Luo, Y. Resistant starch intake alleviates collagen-induced arthritis in mice by modulating gut microbiota and promoting concomitant propionate production. J. Autoimmun. 2020, 116, 102564. [Google Scholar] [CrossRef]
- Martinez, O.D.M.; Gomes, M.J.C.; Grancieri, M.; José, V.P.B.d.S.; Toledo, R.C.L.; Queiroz, V.A.V.; da Silva, B.P.; Martino, H.S.D. Sorghum flour BRS 305 hybrid has the potential to modulate the intestinal microbiota of rats fed with a high-fat high-fructose diet. Eur. J. Nutr. 2022, 62, 647–657. [Google Scholar] [CrossRef] [PubMed]
- Lopes, R.d.C.S.O.; Theodoro, J.M.V.; da Silva, B.P.; Queiroz, V.A.V.; Moreira, M.E.d.C.; Mantovani, H.C.; Hermsdorff, H.H.; Martino, H.S.D. Synbiotic meal decreases uremic toxins in hemodialysis individuals: A placebo-controlled trial. Food Res. Int. 2019, 116, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Toscano, M.; De Grandi, R.; Stronati, L.; De Vecchi, E.; Drago, L. Effect of Lactobacillus rhamnosus HN001 and Bifidobacterium longum BB536 on the healthy gut microbiota composition at phyla and species level: A preliminary study. World J. Gastroenterol. 2017, 23, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Vitellio, P.; Celano, G.; Bonfrate, L.; Gobbetti, M.; Portincasa, P.; De Angelis, M. Effects of Bifidobacterium longum and Lactobacillus rhamnosus on Gut Microbiota in Patients with Lactose Intolerance and Persisting Functional Gastrointestinal Symptoms: A Randomised, Double-Blind, Cross-Over Study. Nutrients 2019, 11, 886. [Google Scholar] [CrossRef] [PubMed]
- Lydia, A.; Indra, T.A.; Rizka, A.; Abdullah, M. The effects of synbiotics on indoxyl sulphate level, constipation, and quality of life associated with constipation in chronic haemodialysis patients: A randomized controlled trial. BMC Nephrol. 2022, 23, 259. [Google Scholar] [CrossRef] [PubMed]
- De Mauri, A.; Carrera, D.; Bagnati, M.; Rolla, R.; Vidali, M.; Chiarinotti, D.; Pane, M.; Amoruso, A.; Del Piano, M. Probiotics-Supplemented Low-Protein Diet for Microbiota Modulation in Patients with Advanced Chronic Kidney Disease (ProLowCKD): Results from a Placebo-Controlled Randomized Trial. Nutrients 2022, 14, 1637. [Google Scholar] [CrossRef] [PubMed]
- Borges, N.A.; Carmo, F.L.; Stockler-Pinto, M.B.; de Brito, J.S.; Dolenga, C.J.; Ferreira, D.C.; Nakao, L.S.; Rosado, A.; Fouque, D.; Mafra, D. Probiotic Supplementation in Chronic Kidney Disease: A Double-blind, Randomized, Placebo-controlled Trial. J. Ren. Nutr. 2018, 28, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ensminger, L.G. The Association of Official Analytical Chemists. Clin. Toxicol. 1976, 9, 471. [Google Scholar] [CrossRef]
- Dixon, N.M. Common knowledge (book excerpt). Ubiquity 2000, 2000, 3. [Google Scholar] [CrossRef]
- dos Santos, L.A.; Lemaire, D.C.; Coutinho-Lima, C.R.O. Tempo de trânsito intestinal de indivíduos portadores de Síndrome Metabólica pela Escala de Bristol. Revista de Ciências Médicas e Biológicas 2017, 16, 338–343. [Google Scholar] [CrossRef]
- Anunciação, P.C.; Cardoso, L.d.M.; Gomes, J.V.P.; Della Lucia, C.M.; Carvalho, C.W.P.; Galdeano, M.C.; Queiroz, V.A.V.; Alfenas, R.d.C.G.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Comparing sorghum and wheat whole grain breakfast cereals: Sensorial acceptance and bioactive compound content. Food Chem. 2017, 221, 984–989. [Google Scholar] [CrossRef] [PubMed]
- WHO Technical Report Series OBESITY: Preventing and Managing the Global Epidemic; WHO: Geneva, Switzerland, 2000.
- de Loor, H.; Meijers, B.K.; Meyer, T.W.; Bammens, B.; Verbeke, K.; Dehaen, W.; Evenepoel, P. Sodium octanoate to reverse indoxyl sulfate and p-cresyl sulfate albumin binding in uremic and normal serum during sample preparation followed by fluorescence liquid chromatography. J. Chromatogr. A 2009, 1216, 4684–4688. [Google Scholar] [CrossRef] [PubMed]
- Ussar, S.; Griffin, N.W.; Bezy, O.; Fujisaka, S.; Vienberg, S.; Softic, S.; Deng, L.; Bry, L.; Gordon, J.I.; Kahn, C.R. Interactions between Gut Microbiota, Host Genetics and Diet Modulate the Predisposition to Obesity and Metabolic Syndrome. Cell Metab. 2015, 22, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, Colonic Fermentation, and Gastrointestinal Health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Ramezani, A.; Massy, Z.A.; Meijers, B.; Evenepoel, P.; Vanholder, R.; Raj, D.S. Role of the Gut Microbiome in Uremia: A Potential Therapeutic Target. Am. J. Kidney Dis. 2016, 67, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Le Chatelier, E.; Nielsen, T.; Qin, J.; Prifti, E.; Hildebrand, F.; Falony, G.; Almeida, M.; Arumugam, M.; Batto, J.-M.; Kennedy, S.; et al. Richness of human gut microbiome correlates with metabolic markers. Nature 2013, 500, 541–546. [Google Scholar] [CrossRef] [PubMed]
- Plata, C.; Cruz, C.; Cervantes, L.G.; Ramírez, V. The gut microbiota and its relationship with chronic kidney disease. Int. Urol. Nephrol. 2019, 51, 2209–2226. [Google Scholar] [CrossRef] [PubMed]
- Esgalhado, M.; Kemp, J.A.; de Paiva, B.R.; Brito, J.S.; Cardozo, L.F.M.F.; Azevedo, R.; Cunha, D.B.; Nakao, L.S.; Mafra, D. Resistant starch type-2 enriched cookies modulate uremic toxins and inflammation in hemodialysis patients: A randomized, double-blind, crossover and placebo-controlled trial. Food Funct. 2020, 11, 2617–2625. [Google Scholar] [CrossRef] [PubMed]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Topping, D.L.; Clifton, P.M. Short-Chain Fatty Acids and Human Colonic Function: Roles of Resistant Starch and Nonstarch Polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Biruete, A.; Cross, T.-W.L.; Allen, J.M.; Kistler, B.M.; de Loor, H.; Evenepoel, P.; Fahey, G.C.; Bauer, L.; Swanson, K.S.; Wilund, K.R. Effect of Dietary Inulin Supplementation on the Gut Microbiota Composition and Derived Metabolites of Individuals Undergoing Hemodialysis: A Pilot Study. J. Ren. Nutr. 2021, 31, 512–522. [Google Scholar] [CrossRef] [PubMed]
- Barros, F.; Awika, J.M.; Rooney, L.W. Interaction of Tannins and Other Sorghum Phenolic Compounds with Starch and Effects on in Vitro Starch Digestibility. J. Agric. Food Chem. 2012, 60, 11609–11617. [Google Scholar] [CrossRef] [PubMed]
- Ben Othman, R.; Ben Amor, N.; Mahjoub, F.; Berriche, O.; El Ghali, C.; Gamoudi, A.; Jamoussi, H. A clinical trial about effects of prebiotic and probiotic supplementation on weight loss, psychological profile and metabolic parameters in obese subjects. Endocrinol. Diabetes Metab. 2023, 6, e402. [Google Scholar] [CrossRef]
- Yildirim, G.K.; Dinleyici, M.; Vandenplas, Y.; Dinleyici, E.C. Effects of Multispecies Synbiotic Supplementation on Anthropometric Measurements, Glucose and Lipid Parameters in Children with Exogenous Obesity: A Randomized, Double Blind, Placebo-Controlled Clinical Trial (Probesity-2 Trial). Front. Nutr. 2022, 9, 898037. [Google Scholar] [CrossRef]
- Su, G.; Qin, X.; Yang, C.; Sabatino, A.; Kelly, J.T.; Avesani, C.M.; Carrero, J.J. Fiber intake and health in people with chronic kidney disease. Clin. Kidney J. 2022, 15, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Mora, J.; Martínez-Hernández, N.E.; Martindel Campo-Lopez, F.; Viramontes-Hörner, D.; Vizmanos-Lamotte, B.; Muñoz-Valle, J.F.; García-García, G.; Parra-Rojas, I.; Castro-Alarcón, N. Effects of a Symbiotic on Gut Microbiota in Mexican Patients with End-Stage Renal Disease. J. Ren. Nutr. 2014, 24, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Hobby, G.P.; Karaduta, O.; Dusio, G.F.; Singh, M.; Zybailov, B.L.; John Arthur, X.M. REVIEW Translational Physiology Chronic kidney disease and the gut microbiome. Am. J. Physiol. Renal. Physiol. 2019, 316, 1211–1217. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Elderman, M.E.; El Aidy, S.; Burgerhof, J.G.M.; Visser, H.; Vaughan, E.E.; Faas, M.M.; de Vos, P. Flexibility of Gut Microbiota in Ageing Individuals during Dietary Fiber Long-Chain Inulin Intake. Mol. Nutr. Food Res. 2021, 65, e2000390. [Google Scholar] [CrossRef] [PubMed]
- Ballini, A.; Scacco, S.; Boccellino, M.; Santacroce, L.; Arrigoni, R. Microbiota and Obesity: Where Are We Now? Biology 2020, 9, 415. [Google Scholar] [CrossRef] [PubMed]
- Barber, C.; Mego, M.; Sabater, C.; Vallejo, F.; Bendezu, R.A.; Masihy, M.; Guarner, F.; Espín, J.C.; Margolles, A.; Azpiroz, F. Differential Effects of Western and Mediterranean-Type Diets on Gut Microbiota: A Metagenomics and Metabolomics Approach. Nutrients 2021, 13, 2638. [Google Scholar] [CrossRef] [PubMed]
- Sato, J.; O’Brien, T.; Katusic, Z.S.; Fu, A.; Nygren, J.; Singh, R.; Nair, K. Dietary antioxidants preserve endothelium dependent vasorelaxation in overfed rats. Atherosclerosis 2002, 161, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Luo, S.; Ye, Y.; Yin, S.; Fan, J.; Xia, M. Intermittent Fasting Improves Cardiometabolic Risk Factors and Alters Gut Microbiota in Metabolic Syndrome Patients. J. Clin. Endocrinol. Metab. 2021, 106, 64–79. [Google Scholar] [CrossRef]
- Ampatzoglou, A.; Atwal, K.K.; Maidens, C.M.; Williams, C.L.; Ross, A.B.; Thielecke, F.; Jonnalagadda, S.S.; Kennedy, O.B.; Yaqoob, P. Increased Whole Grain Consumption Does Not Affect Blood Biochemistry, Body Composition, or Gut Microbiology in Healthy, Low-Habitual Whole Grain Consumers. J. Nutr. 2015, 145, 215–221. [Google Scholar] [CrossRef]
- Kopf, J.C.; Suhr, M.J.; Clarke, J.; Eyun, S.-I.; Riethoven, J.-J.M.; Ramer-Tait, A.E.; Rose, D.J. Role of whole grains versus fruits and vegetables in reducing subclinical inflammation and promoting gastrointestinal health in individuals affected by overweight and obesity: A randomized controlled trial. Nutr. J. 2018, 17, 72. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-R.; Shin, J.; Guevarra, R.B.; Lee, J.H.; Kim, D.W.; Seol, K.-H.; Kim, H.B.; Isaacson, R.E. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.M.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Tan, S.W.; Tan, S.G. The Promising Future of Chia, Salvia hispanica L. J. Biomed. Biotechnol. 2012, 2012, 171956. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.-Y.; Guo, X.-Y.; Zhang, B.; Jiang, Y.; Ye, B.-C. Recent Advances of L-ornithine Biosynthesis in Metabolically Engineered Corynebacterium glutamicum. Front. Bioeng. Biotechnol. 2020, 7, 440. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.C.; Gillitt, N.; Jin, F.; Henson, D.A.; Kennerly, K.; Shanely, R.A.; Ore, B.; Su, M.; Schwartz, S. Chia seed supplementation and disease risk factors in overweight women: A metabolomics investigation. J. Altern. Complement. Med. 2012, 18, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Itakura, M.; Sabina, R.L.; Heald, P.W.; Holmes, E.W. Basis for the control of purine biosynthesis by purine ribonucleotides. J. Clin. Investig. 1981, 67, 994–1002. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Wu, G.; Zhou, Z.; Dai, Z.; Sun, Y.; Ji, Y.; Li, W.; Wang, W.; Liu, C.; Han, F.; et al. Glutamine and intestinal barrier function. Amino Acids 2014, 47, 2143–2154. [Google Scholar] [CrossRef]
- Cupisti, A.; D’Alessandro, C.; Gesualdo, L.; Cosola, C.; Gallieni, M.; Egidi, M.F.; Fusaro, M. Non-Traditional Aspects of Renal Diets: Focus on Fiber, Alkali and Vitamin K1 Intake. Nutrients 2017, 9, 444. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbiotic Group | Control Group | p Value |
|---|---|---|---|
| Subjects (n = 39) | 19 | 20 | - |
| Sex | Man: 12 Woman: 7 | Man: 15 Woman: 5 | - |
| Age (years) | 62.85 ± 11.74 | 64.22 ± 9.68 | 0.68 |
| Body weight (kg) | 66.06 ± 10.79 | 59.20 ± 9.89 | 0.05 |
| BMI (kg/m²) | 25.96 ± 4.68 | 23.08 ± 3.09 | 0.05 |
| HD time (months) | 59.60 ± 72.79 | 50.83 ± 55.05 | >0.99 |
| Variables | Symbiotic Group | p¹ Value | Control Group | p¹ Value | Delta p Value | ||
|---|---|---|---|---|---|---|---|
| Baseline | Endpoint | Baseline | Endpoint | ||||
| IS (mg/dL) | 140.46 ± 70.85 | 115.95 ± 55.65 | 0.1297 | 151.94 ± 61.13 | 147.74 ± 51.36 | 0.7075 | 0.1758 |
| IAA (µg/L) | 24.21 ± 13.73 | 18.19 ± 10.67 | 0.0030 | 18.62 ± 13.44 | 15.62 ± 4.91 | 0.9764 | 0.3891 |
| p-CS (mg/L) | 386.47 ± 197.99 | 241.13 ± 99.79 | 0.0001 | 289.21 ± 245.62 | 295.02 ± 127.18 | 0.065 | 0.3524 |
| Urea Creatinine | 37.15 ± 16.70 8.31 ± 3.23 | 37.80 ± 12.42 8.67 ± 2.60 | 0.8835 0.1011 | 43.70 ± 32.88 8.44 ± 3.30 | 33.09 ± 16.35 9.01 ± 3.93 | 0.1482 0.1530 | 0.2175 0.7016 |
| Variables | Symbiotic Group | p¹ Value | Control Group | p¹ Value | Delta p Value | ||
|---|---|---|---|---|---|---|---|
| Baseline | Endpoint | Baseline | Endpoint | ||||
| Acetic acid | 3.71 ± 1.76 | 7.00 ± 2.60 | <0.0001 | 4.91 ± 2.07 | 7.95 ± 3.98 | 0.0007 | 0.1758 |
| Propionic acid | 2.41 ± 2.04 | 6.38 ± 3.73 | <0.0001 | 3.05 ± 2.91 | 5.95 ± 3.71 | 0.0050 | 0.3891 |
| Butyric acid | 2.26 ± 1.66 | 3.87 ± 2.38 | 0.040 | 2.57 ± 1.91 | 4.42 ± 4.10 | 0.0636 | 0.3524 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lúcio, H.G.; Lopes, R.d.C.S.O.; Gomes, M.J.C.; da Silva, A.; Grancieri, M.; Della Lucia, C.M.; Queiroz, V.A.V.; da Silva, B.P.; Martino, H.S.D. A Symbiotic Meal Containing Extruded Sorghum and Probiotic (Bifidobacterium longum) Ameliorated Intestinal Health Markers in Individuals with Chronic Kidney Disease: A Secondary Analysis of a Subsample from a Previous Randomized and Controlled Clinical Trial. Nutrients 2024, 16, 1852. https://doi.org/10.3390/nu16121852
Lúcio HG, Lopes RdCSO, Gomes MJC, da Silva A, Grancieri M, Della Lucia CM, Queiroz VAV, da Silva BP, Martino HSD. A Symbiotic Meal Containing Extruded Sorghum and Probiotic (Bifidobacterium longum) Ameliorated Intestinal Health Markers in Individuals with Chronic Kidney Disease: A Secondary Analysis of a Subsample from a Previous Randomized and Controlled Clinical Trial. Nutrients. 2024; 16(12):1852. https://doi.org/10.3390/nu16121852
Chicago/Turabian StyleLúcio, Haira Guedes, Rita de Cassia Stampini Oliveira Lopes, Mariana Juste Contin Gomes, Alessandra da Silva, Mariana Grancieri, Ceres Mattos Della Lucia, Valéria Aparecida Vieira Queiroz, Bárbara Pereira da Silva, and Hercia Stampini Duarte Martino. 2024. "A Symbiotic Meal Containing Extruded Sorghum and Probiotic (Bifidobacterium longum) Ameliorated Intestinal Health Markers in Individuals with Chronic Kidney Disease: A Secondary Analysis of a Subsample from a Previous Randomized and Controlled Clinical Trial" Nutrients 16, no. 12: 1852. https://doi.org/10.3390/nu16121852