
Efficacy of Lactococcus lactis subsp. lactis LY-66 and Lactobacillus plantarum PL-02 in Enhancing Explosive Strength and Endurance: A Randomized, Double-Blinded Clinical Trial

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Test Samples
2.2. Participants
2.3. Experimental Design and Content
2.4. Body Composition Analysis
2.5. Exercise Program to Induce Muscle Fatigue and Soreness
2.6. Countermovement Jump Assessment (CMJ)
2.7. Isometric Mid-Thigh Pull (IMTP)
2.8. Wingate Anaerobic Test (WAnT)
2.9. Measuring VO2max
2.10. Physiological Observation and Serum Biochemical Analysis
2.11. Fecal DNA Extraction and Next Generation Sequencing (NGS) Analysis
2.12. Bioinformatics Analysis and Statistics
2.13. Statistics
3. Results
3.1. The Effects of Supplementing with PL-02, LY-66, or PL-02+LY-66 Probiotics Were Beneficial for Body Composition
3.2. Effects of Biochemical Characteristics of Subjects before and after 6-Week Probiotic Intervention with PL-02, LY-66, or PL-02+LY-66
3.3. Effect of PL-02, LY-66, or PL-02+LY-66 Probiotic Supplementation on Maximal Oxygen Consumption
3.4. Effect of PL-02, LY-66, or PL-02+LY-66 Probiotic Supplementation on Enhancing Strength Performance, Explosive Power, and Anaerobic Power
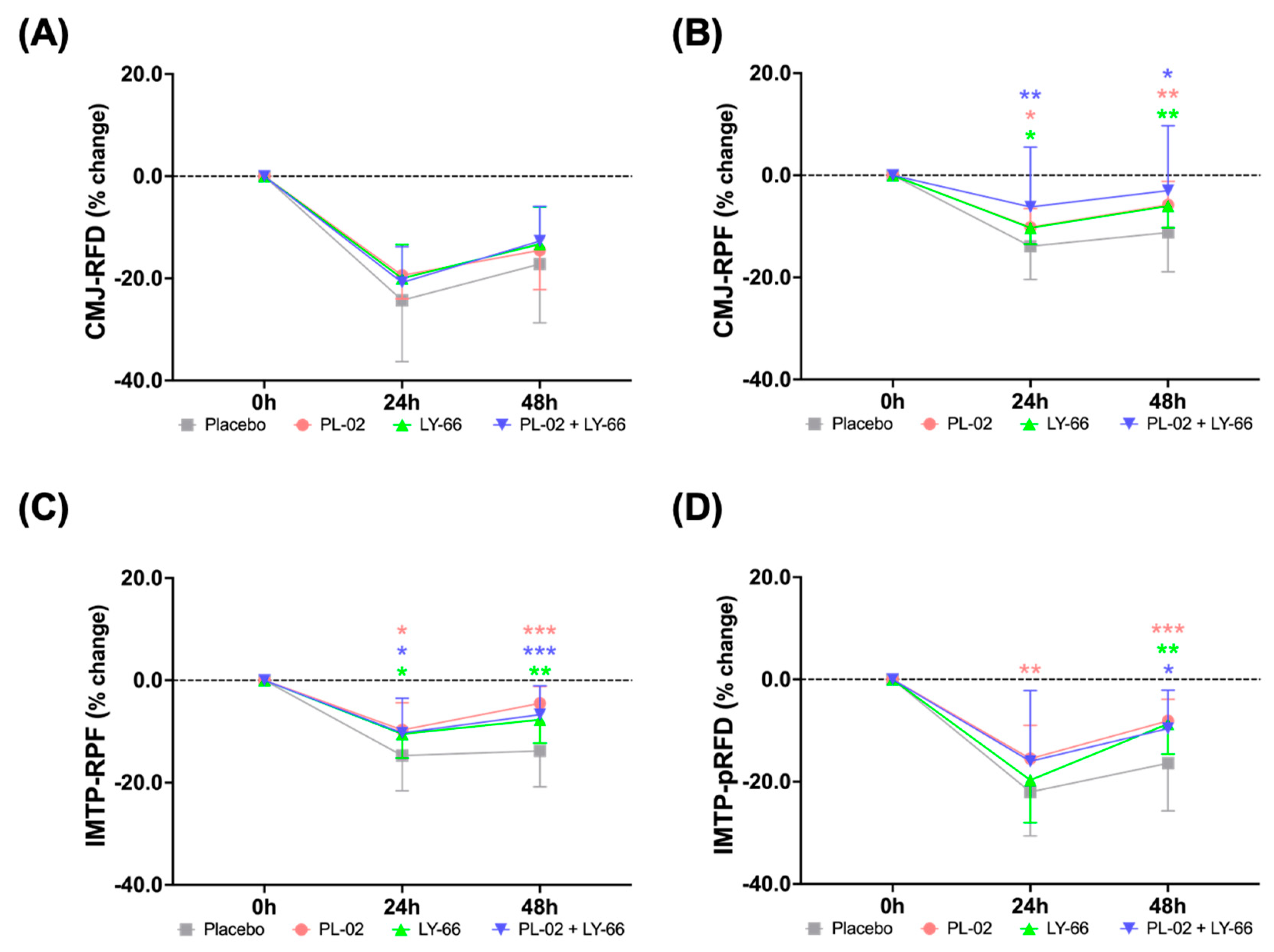
3.5. Effect of PL-02, LY-66, or PL-02+LY-66 Probiotic Supplementation on Restoring Muscle Performance and Explosive Strength after EIMD
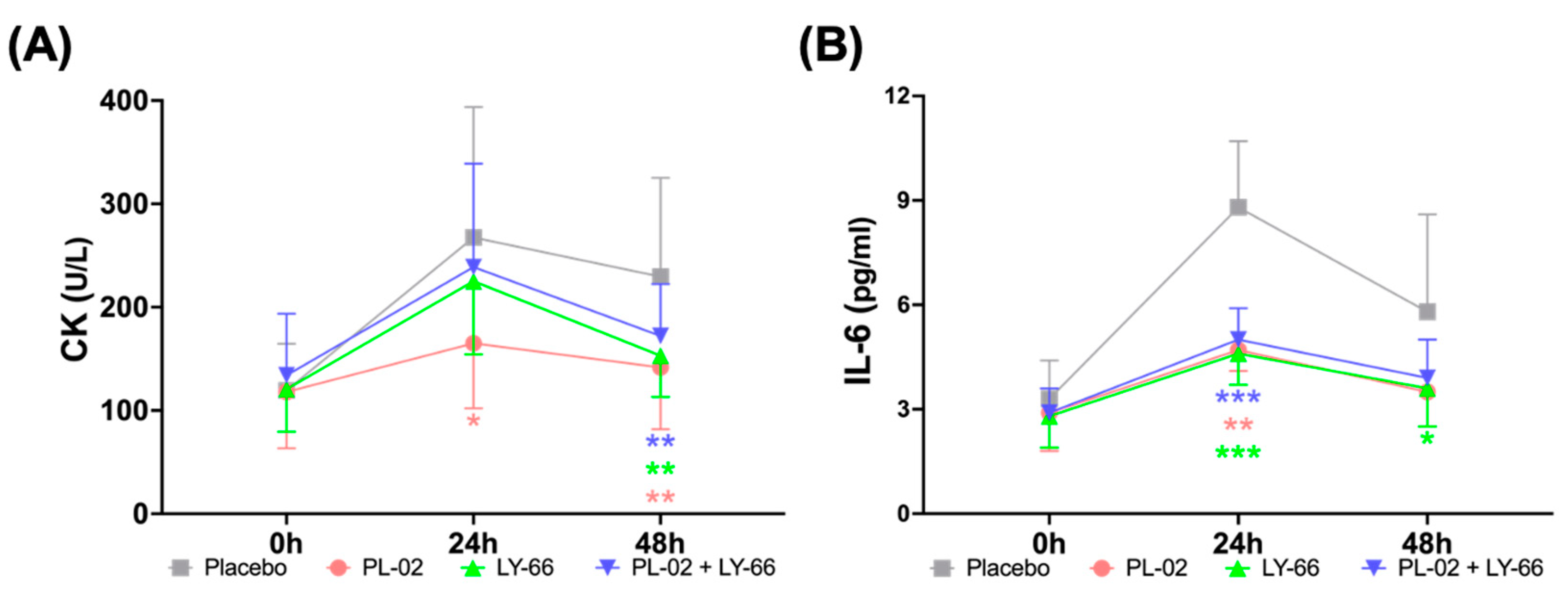
3.6. Effect of PL-02, LY-66, or PL-02+LY-66 Probiotic Supplementation on Blood Markers of Muscle Damage and Inflammation
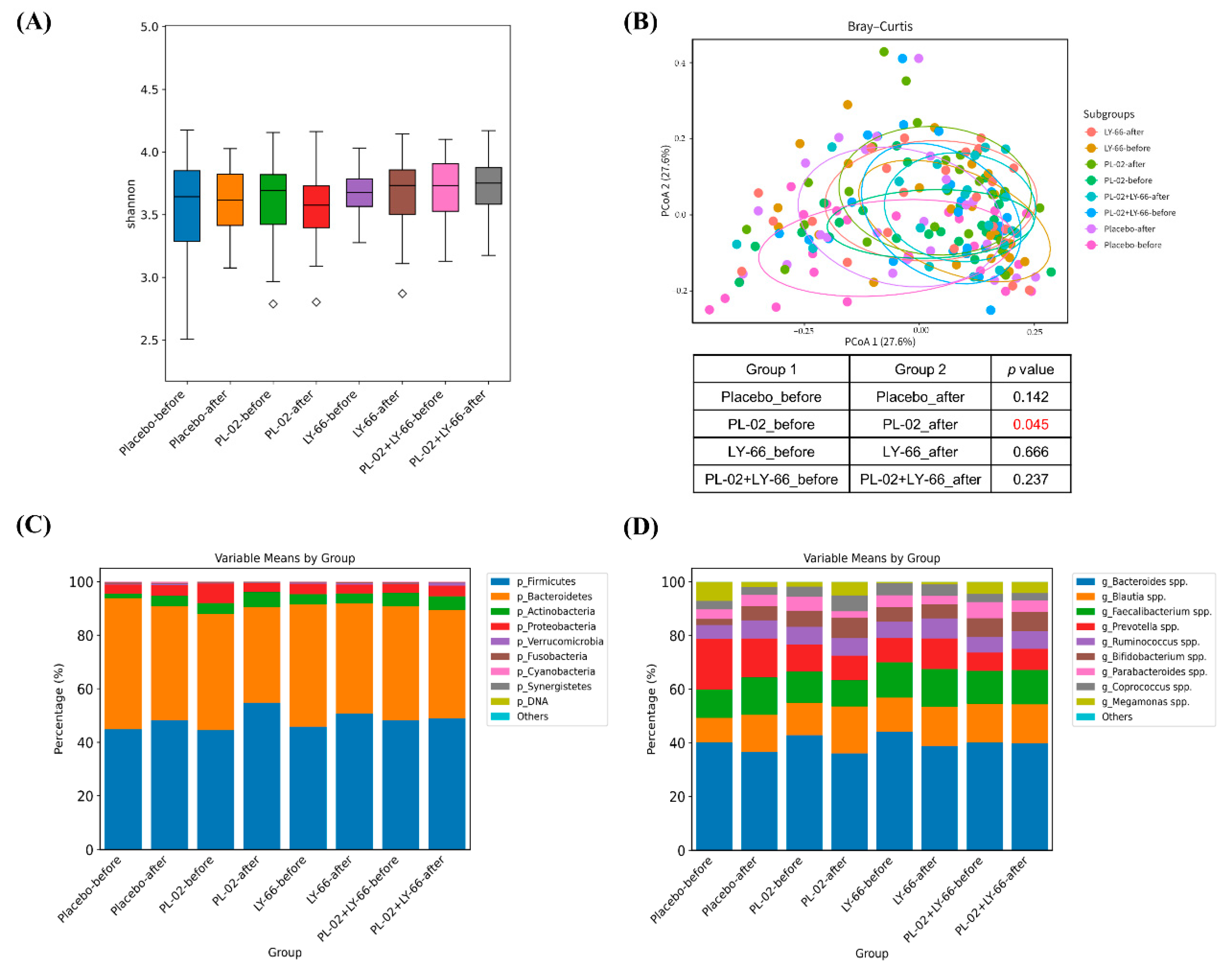
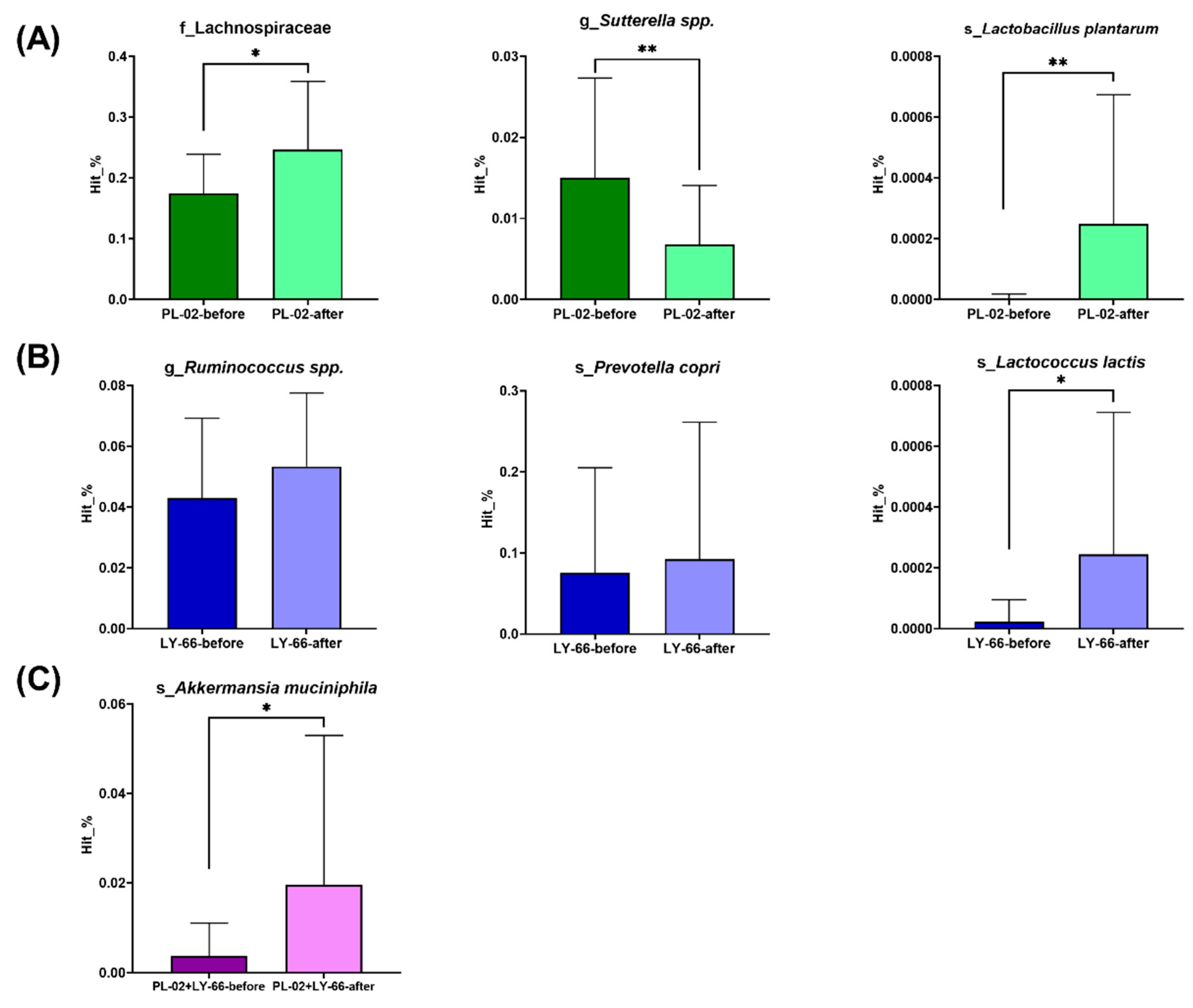
3.7. Effect of PL-02, LY-66, or PL-02+LY-66 Probiotic Supplementation on Modulation of Gut Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Penedo, F.J.; Dahn, J.R. Exercise and well-being: A review of mental and physical health benefits associated with physical activity. Curr. Opin. Psychiatry 2005, 18, 189–193. [Google Scholar] [CrossRef]
- Paulsen, G.; Ramer Mikkelsen, U.; Raastad, T.; Peake, J.M. Leucocytes, cytokines and satellite cells: What role do they play in muscle damage and regeneration following eccentric exercise? Exerc. Immunol. Rev. 2012, 18, 42–97. [Google Scholar]
- Sheridan, S.; McCarren, A.; Gray, C.; Murphy, R.P.; Harrison, M.; Wong, S.H.; Moyna, N.M. Maximal oxygen consumption and oxygen uptake efficiency in adolescent males. J. Exerc. Sci. Fit. 2021, 19, 75–80. [Google Scholar] [CrossRef]
- Denham, J.; Scott-Hamilton, J.; Hagstrom, A.D.; Gray, A.J. Cycling power outputs predict functional threshold power and maximum oxygen uptake. J. Strength. Cond. Res. 2020, 34, 3489–3497. [Google Scholar] [CrossRef]
- Daly, R.M. Independent and combined effects of exercise and vitamin D on muscle morphology, function and falls in the elderly. Nutrients 2010, 2, 1005–1017. [Google Scholar] [CrossRef]
- Kell, R.T.; Bell, G.; Quinney, A. Musculoskeletal fitness, health outcomes and quality of life. Sports Med. 2001, 31, 863–873. [Google Scholar] [CrossRef]
- Gremeaux, V.; Gayda, M.; Lepers, R.; Sosner, P.; Juneau, M.; Nigam, A. Exercise and longevity. Maturitas 2012, 73, 312–317. [Google Scholar] [CrossRef]
- Ekeland, E.; Heian, F.; Hagen, K.B.; Abbott, J.; Nordheim, L. Exercise to improve self-esteem in children and young people. Cochrane Database Syst. Rev. 2004, CD003683. [Google Scholar] [CrossRef]
- Larun, L.; Nordheim, L.V.; Ekeland, E.; Hagen, K.B.; Heian, F. Exercise in prevention and treatment of anxiety and depression among children and young people. Cochrane Database Syst. Rev. 2006, CD004691. [Google Scholar] [CrossRef]
- Fatouros, I.G.; Jamurtas, A.Z. Insights into the molecular etiology of exercise-induced inflammation: Opportunities for optimizing performance. J. Inflamm. Res. 2016, 9, 175–186. [Google Scholar] [CrossRef]
- Lee, M.-C.; Ho, C.-S.; Hsu, Y.-J.; Huang, C.-C. Live and heat-killed probiotic Lactobacillus paracasei PS23 accelerated the improvement and recovery of strength and damage biomarkers after exercise-induced muscle damage. Nutrients 2022, 14, 4563. [Google Scholar] [CrossRef]
- Fernández-Lázaro, D.; Fernandez-Lazaro, C.I.; Mielgo-Ayuso, J.; Navascués, L.J.; Córdova Martínez, A.; Seco-Calvo, J. The role of selenium mineral trace element in exercise: Antioxidant defense system, muscle performance, hormone response, and athletic performance. A systematic review. Nutrients 2020, 12, 1790. [Google Scholar] [CrossRef]
- Fernández-Lázaro, D.; Sanchez-Serrano, N.; Rabail, R.; Aadil, R.M.; Mielgo-Ayuso, J.; Radesca Fabiano, K.; Garrosa, E. Is Probiotics Supplementation an Appropriate Strategy to Modulate Inflammation in Physically Active Healthy Adults or Athletes? A Systematic Review. Appl. Sci. 2023, 13, 3448. [Google Scholar] [CrossRef]
- Ticinesi, A.; Lauretani, F.; Milani, C.; Nouvenne, A.; Tana, C.; Del Rio, D.; Maggio, M.; Ventura, M.; Meschi, T. Aging gut microbiota at the cross-road between nutrition, physical frailty, and sarcopenia: Is there a gut–muscle axis? Nutrients 2017, 9, 1303. [Google Scholar] [CrossRef]
- Scheiman, J.; Luber, J.M.; Chavkin, T.A.; MacDonald, T.; Tung, A.; Pham, L.-D.; Wibowo, M.C.; Wurth, R.C.; Punthambaker, S.; Tierney, B.T. Meta-omics analysis of elite athletes identifies a performance-enhancing microbe that functions via lactate metabolism. Nat. Med. 2019, 25, 1104–1109. [Google Scholar] [CrossRef]
- Przewłócka, K.; Folwarski, M.; Kaźmierczak-Siedlecka, K.; Skonieczna-Żydecka, K.; Kaczor, J.J. Gut-muscle axis exists and may affect skeletal muscle adaptation to training. Nutrients 2020, 12, 1451. [Google Scholar] [CrossRef]
- Jager, R.; Shields, K.A.; Lowery, R.P.; De Souza, E.O.; Partl, J.M.; Hollmer, C.; Purpura, M.; Wilson, J.M. Probiotic Bacillus coagulans GBI-30, 6086 reduces exercise-induced muscle damage and increases recovery. PeerJ 2016, 4, e2276. [Google Scholar] [CrossRef]
- Lin, C.-L.; Hsu, Y.-J.; Ho, H.-H.; Chang, Y.-C.; Kuo, Y.-W.; Yeh, Y.-T.; Tsai, S.-Y.; Chen, C.-W.; Chen, J.-F.; Huang, C.-C. Bifidobacterium longum subsp. longum OLP-01 supplementation during endurance running training improves exercise performance in middle-and long-distance runners: A double-blind controlled trial. Nutrients 2020, 12, 1972. [Google Scholar] [CrossRef]
- Huang, W.C.; Hsu, Y.J.; Huang, C.C.; Liu, H.C.; Lee, M.C. Exercise Training Combined with Bifidobacterium longum OLP-01 Supplementation Improves Exercise Physiological Adaption and Performance. Nutrients 2020, 12, 1145. [Google Scholar] [CrossRef]
- Lin, W.Y.; Kuo, Y.W.; Lin, J.H.; Lin, C.H.; Chen, J.F.; Tsai, S.Y.; Lee, M.C.; Hsu, Y.J.; Huang, C.C.; Tsou, Y.A.; et al. Probiotic Strains Isolated from an Olympic Woman’s Weightlifting Gold Medalist Increase Weight Loss and Exercise Performance in a Mouse Model. Nutrients 2022, 14, 1270. [Google Scholar] [CrossRef]
- Dos’Santos, T.; Thomas, C.; Comfort, P.; McMahon, J.J.; Jones, P.A.; Oakley, N.P.; Young, A.L. Between-Session Reliability of Isometric Midthigh Pull Kinetics and Maximal Power Clean Performance in Male Youth Soccer Players. J. Strength. Cond. Res. 2018, 32, 3364–3372. [Google Scholar] [CrossRef]
- Huang, C.C.; Lee, M.C.; Ho, C.S.; Hsu, Y.J.; Ho, C.C.; Kan, N.W. Protective and Recovery Effects of Resveratrol Supplementation on Exercise Performance and Muscle Damage following Acute Plyometric Exercise. Nutrients 2021, 13, 3217. [Google Scholar] [CrossRef]
- Bruce, R.A.; Kusumi, F.; Hosmer, D. Maximal oxygen intake and nomographic assessment of functional aerobic impairment in cardiovascular disease. Am. Heart J. 1973, 85, 546–562. [Google Scholar] [CrossRef]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Kramer, A. An Overview of the Beneficial Effects of Exercise on Health and Performance. Adv. Exp. Med. Biol. 2020, 1228, 3–22. [Google Scholar]
- Moreira, J.B.; Wohlwend, M.; Wisløff, U. Exercise and cardiac health: Physiological and molecular insights. Nat. Metab. 2020, 2, 829–839. [Google Scholar] [CrossRef]
- Vechetti, I.J., Jr.; Valentino, T.; Mobley, C.B.; McCarthy, J.J. The role of extracellular vesicles in skeletal muscle and systematic adaptation to exercise. J. Physiol. 2021, 599, 845–861. [Google Scholar] [CrossRef]
- Giron, M.; Thomas, M.; Dardevet, D.; Chassard, C.; Savary-Auzeloux, I. Gut microbes and muscle function: Can probiotics make our muscles stronger? J. Cachexia Sarcopenia Muscle 2022, 13, 1460–1476. [Google Scholar] [CrossRef]
- Chew, W.; Lim, Y.P.; Lim, W.S.; Chambers, E.S.; Frost, G.; Wong, S.H.; Ali, Y. Gut-muscle crosstalk. A perspective on influence of microbes on muscle function. Front. Med. 2023, 9, 1065365. [Google Scholar] [CrossRef]
- Li, G.; Jin, B.; Fan, Z. Mechanisms involved in gut microbiota regulation of skeletal muscle. Oxidative Med. Cell. Longev. 2022, 2022, 2151191. [Google Scholar] [CrossRef]
- Lahiri, S.; Kim, H.; Garcia-Perez, I.; Reza, M.M.; Martin, K.A.; Kundu, P.; Cox, L.M.; Selkrig, J.; Posma, J.M.; Zhang, H. The gut microbiota influences skeletal muscle mass and function in mice. Sci. Transl. Med. 2019, 11, eaan5662. [Google Scholar] [CrossRef]
- Qi, R.; Sun, J.; Qiu, X.; Zhang, Y.; Wang, J.; Wang, Q.; Huang, J.; Ge, L.; Liu, Z. The intestinal microbiota contributes to the growth and physiological state of muscle tissue in piglets. Sci. Rep. 2021, 11, 11237. [Google Scholar] [CrossRef]
- Frampton, J.; Murphy, K.G.; Frost, G.; Chambers, E.S. Short-chain fatty acids as potential regulators of skeletal muscle metabolism and function. Nat. Metab. 2020, 2, 840–848. [Google Scholar] [CrossRef]
- Yoshida, T.; Delafontaine, P. Mechanisms of IGF-1-mediated regulation of skeletal muscle hypertrophy and atrophy. Cells 2020, 9, 1970. [Google Scholar] [CrossRef]
- Bhardwaj, G.; Penniman, C.M.; Jena, J.; Beltran, P.A.S.; Foster, C.; Poro, K.; Junck, T.L.; Hinton, A.O.; Souvenir, R.; Fuqua, J.D. Insulin and IGF-1 receptors regulate complex I–dependent mitochondrial bioenergetics and supercomplexes via FoxOs in muscle. J. Clin. Investig. 2021, 131, e146415. [Google Scholar] [CrossRef]
- Lv, W.Q.; Lin, X.; Shen, H.; Liu, H.M.; Qiu, X.; Li, B.Y.; Shen, W.D.; Ge, C.L.; Lv, F.Y.; Shen, J. Human gut microbiome impacts skeletal muscle mass via gut microbial synthesis of the short-chain fatty acid butyrate among healthy menopausal women. J. Cachexia Sarcopenia Muscle 2021, 12, 1860–1870. [Google Scholar] [CrossRef]
- Chen, W.; Zhang, S.; Wu, J.; Ye, T.; Wang, S.; Wang, P.; Xing, D. Butyrate-producing bacteria and the gut-heart axis in atherosclerosis. Clin. Chim. Acta 2020, 507, 236–241. [Google Scholar] [CrossRef]
- Pant, K.; Venugopal, S.K.; Pisarello, M.J.L.; Gradilone, S.A. The role of gut microbiome-derived short chain fatty acid butyrate in hepatobiliary diseases. Am. J. Pathol. 2023, 193, 1455–1467. [Google Scholar] [CrossRef]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef]
- Liu, H.; Xi, Q.; Tan, S.; Qu, Y.; Meng, Q.; Zhang, Y.; Cheng, Y.; Wu, G. The metabolite butyrate produced by gut microbiota inhibits cachexia-associated skeletal muscle atrophy by regulating intestinal barrier function and macrophage polarization. Int. Immunopharmacol. 2023, 124, 111001. [Google Scholar] [CrossRef] [PubMed]
- Derqaoui, S.; Oukessou, M.; Attrassi, K.; Elftouhy, F.Z.; Nassik, S. Detection of Sutterella spp. in Broiler Liver and Breast. Front. Vet. Sci. 2022, 9, 859902. [Google Scholar] [CrossRef] [PubMed]
- Salameh, M.; Burney, Z.; Mhaimeed, N.; Laswi, I.; Yousri, N.A.; Bendriss, G.; Zakaria, D. The role of gut microbiota in atopic asthma and allergy, implications in the understanding of disease pathogenesis. Scand. J. Immunol. 2020, 91, e12855. [Google Scholar] [CrossRef]
- Mabalirajan, U.; Ahmad, T.; Rehman, R.; Leishangthem, G.D.; Dinda, A.K.; Agrawal, A.; Ghosh, B.; Sharma, S.K. Baicalein reduces airway injury in allergen and IL-13 induced airway inflammation. PLoS ONE 2013, 8, e62916. [Google Scholar] [CrossRef]
- Ma, J.-X.; Xiao, X.; Zhou, K.-F.; Huang, G.; Ao, B.; Zhang, Y.; Gao, W.-J.; Lei, T.; Yang, L.; Fan, X.-C. Herb pair of Ephedrae Herba-Armeniacae Semen Amarum alleviates airway injury in asthmatic rats. J. Ethnopharmacol. 2021, 269, 113745. [Google Scholar] [CrossRef]
- Chai, L.; Luo, Q.; Cai, K.; Wang, K.; Xu, B. Reduced fecal short-chain fatty acids levels and the relationship with gut microbiota in IgA nephropathy. BMC Nephrol. 2021, 22, 209. [Google Scholar] [CrossRef]
- Lakshmanan, A.P.; Al Zaidan, S.; Bangarusamy, D.K.; Al-Shamari, S.; Elhag, W.; Terranegra, A. Increased relative abundance of ruminoccocus is associated with reduced cardiovascular risk in an obese population. Front. Nutr. 2022, 9, 849005. [Google Scholar] [CrossRef]
- Asnicar, F.; Berry, S.E.; Valdes, A.M.; Nguyen, L.H.; Piccinno, G.; Drew, D.A.; Leeming, E.; Gibson, R.; Le Roy, C.; Khatib, H.A. Microbiome connections with host metabolism and habitual diet from 1098 deeply phenotyped individuals. Nat. Med. 2021, 27, 321–332. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Sun, Y.; Ip, L.Y.T.; Wang, L.; Chan, F.K.; Miao, Y.; Ng, S.C. Prevotella species in the human gut is primarily comprised of Prevotella copri, Prevotella stercorea and related lineages. Sci. Rep. 2022, 12, 9055. [Google Scholar] [CrossRef]
- Everard, A.; Lazarevic, V.; Derrien, M.; Girard, M.; Muccioli, G.G.; Neyrinck, A.M.; Possemiers, S.; Van Holle, A.; François, P.; de Vos, W.M. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 2011, 60, 2775–2786. [Google Scholar] [CrossRef]
- Cani, P.D.; Depommier, C.; Derrien, M.; Everard, A.; de Vos, W.M. Akkermansia muciniphila: Paradigm for next-generation beneficial microorganisms. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.-C.; Hsu, Y.-J.; Ho, H.H.; Kuo, Y.W.; Lin, W.-Y.; Tsai, S.-Y.; Chen, W.-L.; Lin, C.-L.; Huang, C.-C. Effectiveness of human-origin Lactobacillus plantarum PL-02 in improving muscle mass, exercise performance and anti-fatigue. Sci. Rep. 2021, 11, 19469. [Google Scholar] [CrossRef]
- Prokopidis, K.; Giannos, P.; Kirwan, R.; Ispoglou, T.; Galli, F.; Witard, O.C.; Triantafyllidis, K.K.; Kechagias, K.S.; Morwani-Mangnani, J.; Ticinesi, A. Impact of probiotics on muscle mass, muscle strength and lean mass: A systematic review and meta-analysis of randomized controlled trials. J. Cachexia Sarcopenia Muscle 2023, 14, 30–44. [Google Scholar] [CrossRef]
- Lim, S.; Moon, J.H.; Shin, C.M.; Jeong, D.; Kim, B. Effect of Lactobacillus sakei, a probiotic derived from kimchi, on body fat in Koreans with obesity: A randomized controlled study. Endocrinol. Metab. 2020, 35, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.G.; Yang, Y.J.; Yoon, Y.S.; Lee, E.S.; Lee, J.H.; Jeong, Y.; Kang, C.H. Effect of MED-02 Containing Two Probiotic Strains, Limosilactobacillus fermentum MG4231 and MG4244, on Body Fat Reduction in Overweight or Obese Subjects: A Randomized, Multicenter, Double-Blind, Placebo-Controlled Study. Nutrients 2022, 14, 3583. [Google Scholar] [CrossRef] [PubMed]
- Noakes, T.D. Testing for maximum oxygen consumption has produced a brainless model of human exercise performance. Br. J. Sports Med. 2008, 42, 551–555. [Google Scholar]
- Astorino, T.A.; Allen, R.P.; Roberson, D.W.; Jurancich, M. Effect of high-intensity interval training on cardiovascular function, VO2max, and muscular force. J. Strength Cond. Res. 2012, 26, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Gist, N.H.; Freese, E.C.; Cureton, K.J. Comparison of responses to two high-intensity intermittent exercise protocols. J. Strength Cond. Res. 2014, 28, 3033–3040. [Google Scholar] [CrossRef]
- Menz, V.; Marterer, N.; Amin, S.B.; Faulhaber, M.; Hansen, A.B.; Lawley, J.S. Functional vs. Running low-volume high-intensity interval training: Effects on vo2max and muscular endurance. J. Sports Sci. Med. 2019, 18, 497. [Google Scholar]
- Soares, A.D.N.; Wanner, S.P.; Morais, E.S.S.; Hudson, A.S.R.; Martins, F.S.; Cardoso, V.N. Supplementation with Saccharomyces boulardii increases the maximal oxygen consumption and maximal aerobic speed attained by rats subjected to an incremental-speed exercise. Nutrients 2019, 11, 2352. [Google Scholar] [CrossRef]
- Smarkusz-Zarzecka, J.; Ostrowska, L.; Leszczyńska, J.; Orywal, K.; Cwalina, U.; Pogodziński, D. Analysis of the impact of a multi-strain probiotic on body composition and cardiorespiratory fitness in long-distance runners. Nutrients 2020, 12, 3758. [Google Scholar] [CrossRef]
- Marttinen, M.; Ala-Jaakkola, R.; Laitila, A.; Lehtinen, M.J. Gut microbiota, probiotics and physical performance in athletes and physically active individuals. Nutrients 2020, 12, 2936. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Hermans, E.; Saeterbakken, A.H.; Vereide, V.; Nord, I.S.; Stien, N.; Andersen, V. The Effects of 10 Weeks Hangboard Training on Climbing Specific Maximal Strength, Explosive Strength, and Finger Endurance. Front. Sports Act. Living 2022, 4, 888158. [Google Scholar] [CrossRef] [PubMed]
- Stien, N.; Saeterbakken, A.H.; Hermans, E.; Vereide, V.A.; Olsen, E.; Andersen, V. Comparison of climbing-specific strength and endurance between lead and boulder climbers. PLoS ONE 2019, 14, e0222529. [Google Scholar] [CrossRef]
- Hoffman, J.R.; Hoffman, M.W.; Zelicha, H.; Gepner, Y.; Willoughby, D.S.; Feinstein, U.; Ostfeld, I. The effect of 2 weeks of inactivated probiotic Bacillus coagulans on endocrine, inflammatory, and performance responses during self-defense training in soldiers. J. Strength Cond. Res. 2019, 33, 2330–2337. [Google Scholar] [CrossRef] [PubMed]
- Tarik, M.; Ramakrishnan, L.; Bhatia, N.; Goswami, R.; Kandasamy, D.; Roy, A.; Chandran, D.S.; Singh, A.; Upadhyay, A.D.; Kalaivani, M. The effect of Bacillus coagulans Unique IS-2 supplementation on plasma amino acid levels and muscle strength in resistance trained males consuming whey protein: A double-blind, placebo-controlled study. Eur. J. Nutr. 2022, 61, 2673–2685. [Google Scholar] [CrossRef]
- Gomaa, M.A.; Allam, M.G.; Haridi, A.A.; Eliwa, A.-E.M.; Darwish, A.M. High-protein concentrated pro-yogurt (Pro-WPI) enriched with whey protein isolate improved athletic anemia and performance in a placebo-controlled study. Front. Nutr. 2022, 8, 788446. [Google Scholar] [CrossRef]
- Naderi, A.; Rezvani, M.H.; Degens, H. Foam rolling and muscle and joint proprioception after exercise-induced muscle damage. J. Athl. Train. 2020, 55, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Fakhro, M.A.; AlAmeen, F.; Fayad, R. Comparison of total cold-water immersion’s effects to ice massage on recovery from exercise-induced muscle damage. J. Exp. Orthop. 2022, 9, 59. [Google Scholar] [CrossRef]
- Harnett, J.E.; Pyne, D.B.; McKune, A.J.; Penm, J.; Pumpa, K.L. Probiotic supplementation elicits favourable changes in muscle soreness and sleep quality in rugby players. J. Sci. Med. Sport 2021, 24, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Zaragoza, J.; Purpura, M.; Iametti, S.; Marengo, M.; Tinsley, G.M.; Anzalone, A.J.; Oliver, J.M.; Fiore, W.; Biffi, A. Probiotic administration increases amino acid absorption from plant protein: A placebo-controlled, randomized, double-blind, multicenter, crossover study. Probiotics Antimicrob. Proteins 2020, 12, 1330–1339. [Google Scholar] [CrossRef] [PubMed]
- Walden, K.E.; Hagele, A.M.; Orr, L.S.; Gross, K.N.; Krieger, J.M.; Jäger, R.; Kerksick, C.M. Probiotic BC30 Improves Amino Acid Absorption from Plant Protein Concentrate in Older Women. Probiotics Antimicrob. Proteins 2022, 16, 125–137. [Google Scholar] [CrossRef]
- Chen, L.H.; Chang, S.S.; Chang, H.Y.; Wu, C.H.; Pan, C.H.; Chang, C.C.; Chan, C.H.; Huang, H.Y. Probiotic supplementation attenuates age-related sarcopenia via the gut–muscle axis in SAMP8 mice. J. Cachexia Sarcopenia Muscle 2022, 13, 515–531. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Wei, Z.; Wang, X.; Gao, Y.; Wang, D.; Liu, X.; Bai, C.; Su, G.; Yang, L.; Li, G. Myostatin Knockout Affects Mitochondrial Function by Inhibiting the AMPK/SIRT1/PGC1α Pathway in Skeletal Muscle. Int. J. Mol. Sci. 2022, 23, 13703. [Google Scholar] [CrossRef]
- Koh, J.-H.; Pataky, M.W.; Dasari, S.; Klaus, K.A.; Vuckovic, I.; Ruegsegger, G.N.; Kumar, A.P.; Robinson, M.M.; Nair, K.S. Enhancement of anaerobic glycolysis–a role of PGC-1α4 in resistance exercise. Nat. Commun. 2022, 13, 2324. [Google Scholar] [CrossRef]
- Fischer, C.P. Interleukin-6 in acute exercise and training: What is the biological relevance. Exerc. Immunol. Rev. 2006, 12, 41. [Google Scholar]
- Isaacs, A.W.; Macaluso, F.; Smith, C.; Myburgh, K.H. C-reactive protein is elevated only in high creatine kinase responders to muscle damaging exercise. Front. Physiol. 2019, 10, 86. [Google Scholar] [CrossRef]
- Walzik, D.; Joisten, N.; Zacher, J.; Zimmer, P. Transferring clinically established immune inflammation markers into exercise physiology: Focus on neutrophil-to-lymphocyte ratio, platelet-to-lymphocyte ratio and systemic immune-inflammation index. Eur. J. Appl. Physiol. 2021, 121, 1803–1814. [Google Scholar] [CrossRef]
- Cristofori, F.; Dargenio, V.N.; Dargenio, C.; Miniello, V.L.; Barone, M.; Francavilla, R. Anti-inflammatory and immunomodulatory effects of probiotics in gut inflammation: A door to the body. Front. Immunol. 2021, 12, 578386. [Google Scholar] [CrossRef]
- Yeh, W.-L.; Hsu, Y.-J.; Ho, C.-S.; Ho, H.-H.; Kuo, Y.-W.; Tsai, S.-Y.; Huang, C.-C.; Lee, M.-C. Lactobacillus plantarum Pl-02 supplementation combined with resistance training improved muscle mass, force, and exercise performance in mice. Front. Nutr. 2022, 9, 896503. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-C.; Wei, C.-C.; Huang, C.-C.; Chen, W.-L.; Huang, H.-Y. The beneficial effects of Lactobacillus plantarum PS128 on high-intensity, exercise-induced oxidative stress, inflammation, and performance in triathletes. Nutrients 2019, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Pugh, J.N.; Wagenmakers, A.J.; Doran, D.A.; Fleming, S.C.; Fielding, B.A.; Morton, J.P.; Close, G.L. Probiotic supplementation increases carbohydrate metabolism in trained male cyclists: A randomized, double-blind, placebo-controlled crossover trial. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E504–E513. [Google Scholar] [CrossRef] [PubMed]
- Baird, M.F.; Graham, S.M.; Baker, J.S.; Bickerstaff, G.F. Creatine-kinase-and exercise-related muscle damage implications for muscle performance and recovery. J. Nutr. Metab. 2012, 2012, 960363. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placebo | PL-02 | LY-66 | PL-02+LY-66 | |||||
|---|---|---|---|---|---|---|---|---|
| Age (years) | 21.9 ± 1.4 | 21.3 ± 1.5 | 21.1 ± 2.1 | 20.9 ± 2.0 | ||||
| Body composition | Baseline | Endpoint | Baseline | Endpoint | Baseline | Endpoint | Baseline | Endpoint |
| Body weight (kg) | 68.2 ± 12.7 | 68.1 ± 13.1 | 64.1 ± 13.6 | 64.0 ± 13.9 | 62.6 ± 12.4 | 62.5 ± 15.7 | 68.1 ± 16.0 | 67.7 ± 15.7 |
| BMI (kg/m2) | 23.7 ± 2.8 | 23.6 ± 3.1 | 22.5 ± 3.1 | 22.5 ± 3.2 | 22.0 ± 3.2 | 21.9 ± 3.3 | 23.2 ± 3.3 | 23.0 ± 3.3 |
| Muscle mass (kg) | 30.1 ± 6.5 | 29.9 ± 6.0 | 28.3 ± 7.1 | 28.7 ± 7.0 * | 27.7 ± 6.3 | 27.9 ± 6.2 | 30.0 ± 7.9 | 30.0 ± 7.8 |
| FBM (%) | 21.8 ± 6.6 | 21.7 ± 7.0 | 21.2 ± 8.0 | 20.4 ± 7.9 | 20.8 ± 8.1 | 19.8 ± 8.0 * | 21.9 ± 6.5 | 21.2 ± 6.2 ** |
| Parameters | Week | Placebo | PL-02 | LY-66 | PL-02+LY-66 |
|---|---|---|---|---|---|
| AST (U/L) | Week 0 | 22 ± 4 | 23 ± 4 | 22 ± 4 | 21 ± 4 |
| Week 6 | 21 ± 4 | 20 ± 5 | 19 ± 3 | 19 ± 3 | |
| ALT (U/L) | Week 0 | 20 ± 3 | 20 ± 4 | 21 ± 5 | 22 ± 4 |
| Week 6 | 18 ± 3 | 18 ± 4 | 18 ± 5 | 19 ± 4 | |
| TC (mg/dL) | Week 0 | 179 ± 26 | 173 ± 20 | 179 ± 25 | 174 ± 22 |
| Week 6 | 165 ± 21 | 169 ± 26 | 168 ± 23 | 170 ± 25 | |
| TG (mg/dL) | Week 0 | 67 ± 19 | 69 ± 19 | 68 ± 20 | 66 ± 18 |
| Week 6 | 63 ± 17 | 62 ± 18 | 65 ± 17 | 62 ± 14 | |
| HDL (mg/dL) | Week 0 | 66.6 ± 11.7 | 66.6 ± 12.6 | 67.3 ± 14.8 | 63.8 ± 10.7 |
| Week 6 | 60.7 ± 10.7 | 63.0 ± 9.3 | 65.0 ± 13.8 | 65.0 ± 13.8 | |
| LDL (mg/dL) | Week 0 | 94.5 ± 16.4 | 93.4 ± 19.6 | 91.4 ± 18.6 | 90.0 ± 12.0 |
| Week 6 | 88.3 ± 16.1 | 86.2 ± 17.6 | 85.4 ± 17.9 | 86.0 ± 10.6 | |
| BUN (mg/dL) | Week 0 | 14.9 ± 2.9 | 14.7 ± 3.2 | 14.7 ± 4.0 | 14.7 ± 3.3 |
| Week 6 | 14.3 ± 3.4 | 14.0 ± 2.7 | 14.3 ± 3.7 | 14.2 ± 3.3 | |
| CREA (mg/dL) | Week 0 | 0.98 ± 0.24 | 0.94 ± 0.22 | 0.97 ± 0.17 | 0.98 ± 0.19 |
| Week 6 | 0.96 ± 0.19 | 0.99 ± 0.16 | 0.98 ± 0.10 | 0.98 ± 0.12 | |
| TP (g/dL) | Week 0 | 7.24 ± 1.15 | 7.05 ± 1.01 | 7.10 ± 1.21 | 7.04 ± 1.17 |
| Week 6 | 7.62 ± 1.05 | 7.66 ± 0.84 | 7.65 ± 0.76 | 7.45 ± 0.71 | |
| UA (mg/dL) | Week 0 | 4.60 ± 0.96 | 4.25 ± 1.31 | 4.46 ± 1.41 | 4.70 ± 1.33 |
| Week 6 | 4.60 ± 0.76 | 4.54 ± 1.00 | 4.80 ± 1.31 | 4.77 ± 1.08 | |
| Glucose (mg/dL) | Week 0 | 82 ± 12 | 81 ± 9 | 86 ± 15 | 82 ± 11 |
| Week 6 | 84 ± 9 | 84 ± 7 | 84 ± 13 | 82 ± 6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-C.; Hsu, Y.-J.; Chen, M.-T.; Kuo, Y.-W.; Lin, J.-H.; Hsu, Y.-C.; Huang, Y.-Y.; Li, C.-M.; Tsai, S.-Y.; Hsia, K.-C.; et al. Efficacy of Lactococcus lactis subsp. lactis LY-66 and Lactobacillus plantarum PL-02 in Enhancing Explosive Strength and Endurance: A Randomized, Double-Blinded Clinical Trial. Nutrients 2024, 16, 1921. https://doi.org/10.3390/nu16121921
Lee M-C, Hsu Y-J, Chen M-T, Kuo Y-W, Lin J-H, Hsu Y-C, Huang Y-Y, Li C-M, Tsai S-Y, Hsia K-C, et al. Efficacy of Lactococcus lactis subsp. lactis LY-66 and Lactobacillus plantarum PL-02 in Enhancing Explosive Strength and Endurance: A Randomized, Double-Blinded Clinical Trial. Nutrients. 2024; 16(12):1921. https://doi.org/10.3390/nu16121921
Chicago/Turabian StyleLee, Mon-Chien, Yi-Ju Hsu, Mu-Tsung Chen, Yi-Wei Kuo, Jia-Hung Lin, Yu-Chieh Hsu, Yen-Yu Huang, Ching-Min Li, Shin-Yu Tsai, Ko-Chiang Hsia, and et al. 2024. "Efficacy of Lactococcus lactis subsp. lactis LY-66 and Lactobacillus plantarum PL-02 in Enhancing Explosive Strength and Endurance: A Randomized, Double-Blinded Clinical Trial" Nutrients 16, no. 12: 1921. https://doi.org/10.3390/nu16121921
APA StyleLee, M.-C., Hsu, Y.-J., Chen, M.-T., Kuo, Y.-W., Lin, J.-H., Hsu, Y.-C., Huang, Y.-Y., Li, C.-M., Tsai, S.-Y., Hsia, K.-C., Ho, H.-H., & Huang, C.-C. (2024). Efficacy of Lactococcus lactis subsp. lactis LY-66 and Lactobacillus plantarum PL-02 in Enhancing Explosive Strength and Endurance: A Randomized, Double-Blinded Clinical Trial. Nutrients, 16(12), 1921. https://doi.org/10.3390/nu16121921