

The Anticancer Effects and Therapeutic Potential of Kaempferol in Triple-Negative Breast Cancer

Abstract
:1. Introduction
2. Breast Cancer
2.1. Breast Cancer Types
2.2. Treatment Approaches for TNBC
2.3. Natural Compounds Used in Breast Cancer Therapy
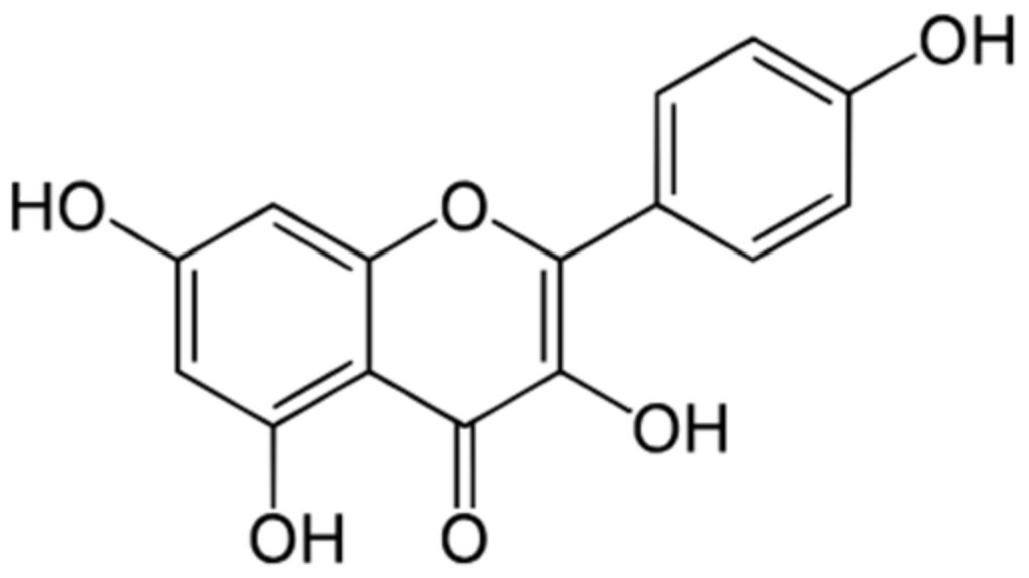
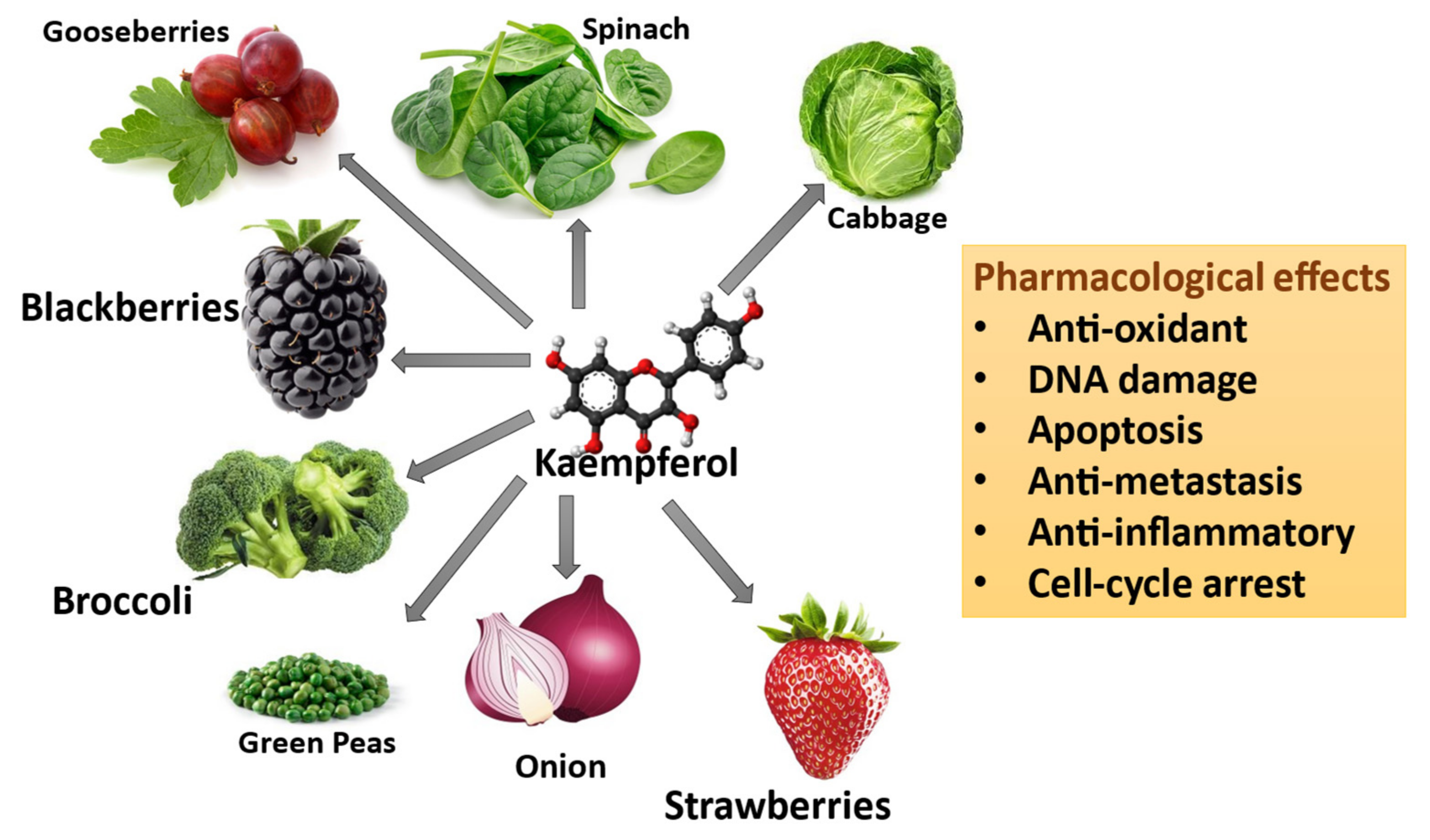
3. Kaempferol
3.1. Kaempferol Absorption and Metabolism
3.2. Kaempferol Pharmacokinetics
3.3. Pharmacological and Toxicological Properties of Kaempferol
4. Anticancer Effects of Kaempferol in Human Cancers
4.1. Kaempferol in Combinatorial Drug Therapy
4.2. Kaempferol Reversal of the Chemoresistance of Chemotherapeutic Agents
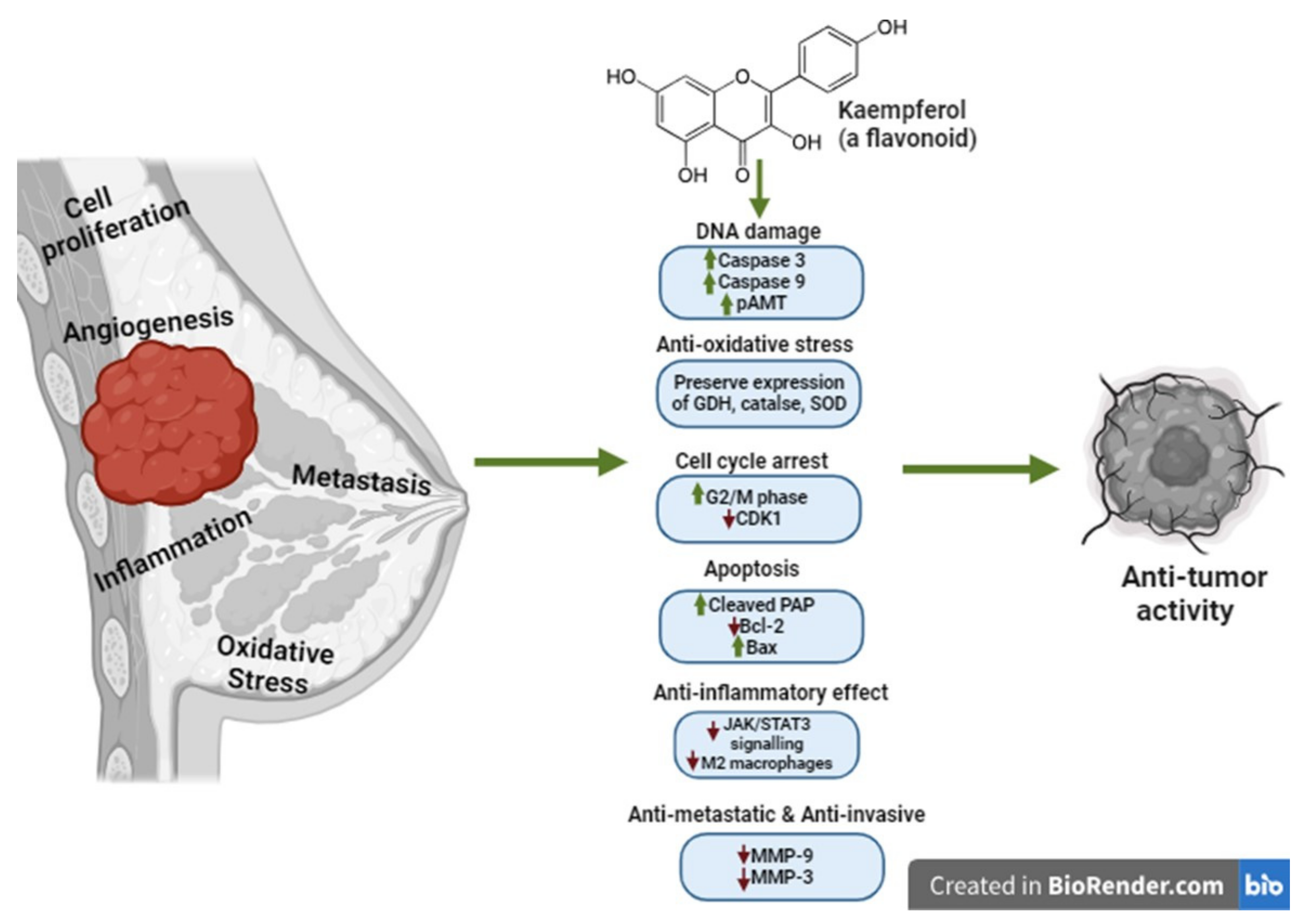
5. Kaempferol Anticancer Mechanisms of Action in Breast Cancer
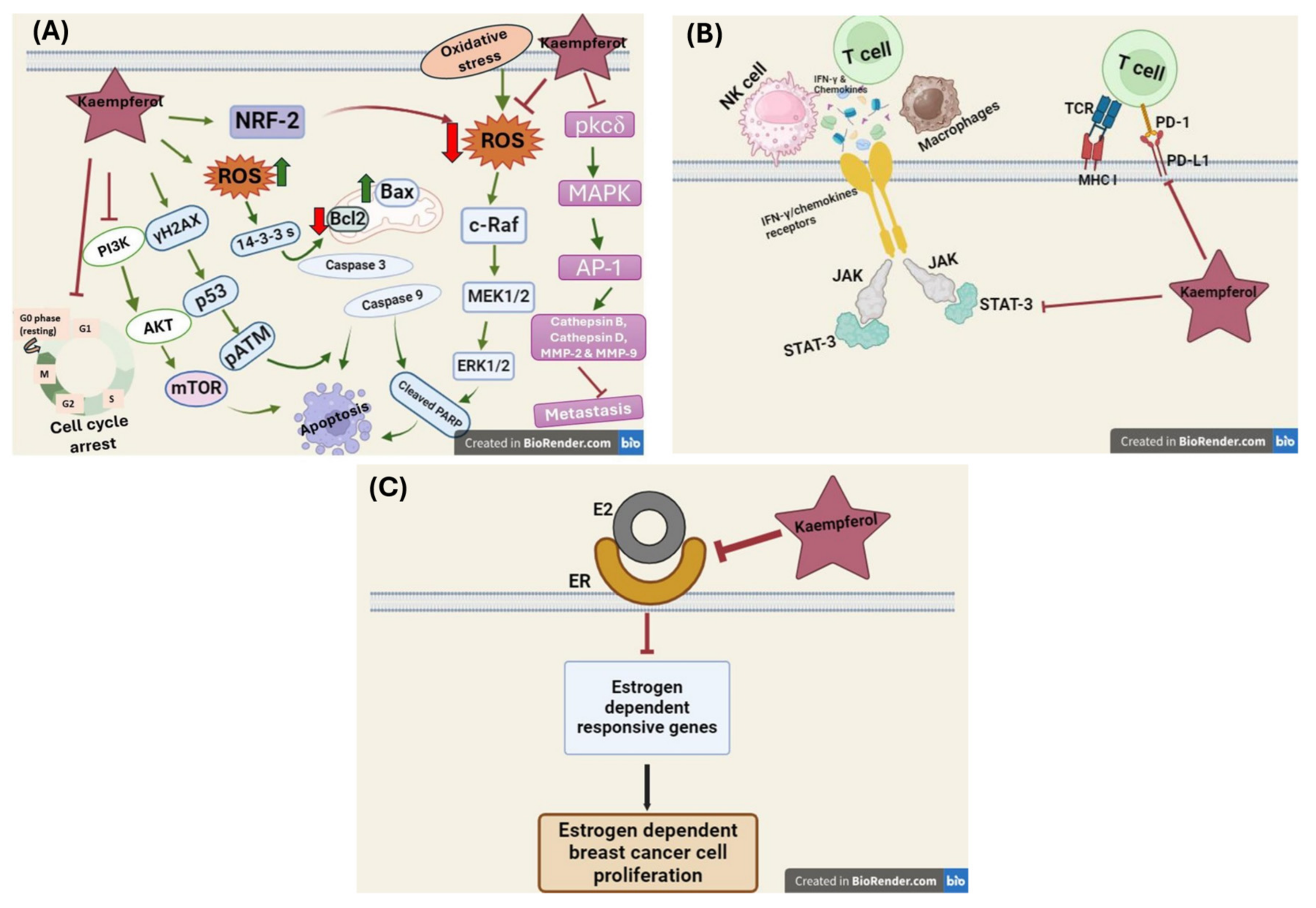
5.1. Effect of Kaempferol on DNA Synthesis Inhibition
5.2. Effect of Kaempferol on ROS Production
5.3. Effect of Kaempferol on Nrf2 Activation
5.4. Effect of Kaempferol on Cell Cycle Arrest
5.5. Effect of Kaempferol on Apoptosis Induction
5.6. Effect of Kaempferol on Cell Invasion and Metastasis Inhibition
5.7. Kaempferol Epigenetic Modulation
5.8. Effect of Kaempferol on the Tumor Microenvironment and Immune Response
5.8.1. Effect of Kaempferol on Tumor-Associated Macrophages (TAMs)
5.8.2. Effect of Kaempferol on the Expression of CCL2
6. Addressing Kaempferol’s Poor Solubility: Pharmaceutical Formulations of Kaempferol
7. Clinical Translation of Kaempferol as an Anticancer Agent
8. Critical Areas for Future Research
8.1. Extrachromosomal Circular DNA (ecDNA) as a Target in Cancer Therapy
8.2. The Regulatory Role of MicroRNAs (miRNAs) in Breast Cancer
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Breast Cancer Facts & Statistics 2023. Available online: https://www.breastcancer.org/facts-statistics (accessed on 13 June 2023).
- Breast Cancer Facts & Figures | American Cancer Society. Available online: https://www.cancer.org/research/cancer-facts-statistics/breast-cancer-facts-figures.html (accessed on 13 June 2023).
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Moulder, S.; Hortobagyi, G. Advances in the Treatment of Breast Cancer. Clin. Pharmacol. Ther. 2008, 83, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Barzaman, K.; Karami, J.; Zarei, Z.; Hosseinzadeh, A.; Kazemi, M.H.; Moradi-Kalbolandi, S.; Safari, E.; Farahmand, L. Breast cancer: Biology, biomarkers, and treatments. Int. Immunopharmacol. 2020, 84, 106535. [Google Scholar] [CrossRef] [PubMed]
- Fisusi, F.A.; Akala, E.O. Drug Combinations in Breast Cancer Therapy. Pharm. Nanotechnol. 2019, 7, 3–23. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, R.; Ordóñez-Morán, P.; Allegrucci, C. Challenges for Triple Negative Breast Cancer Treatment: Defeating Heterogeneity and Cancer Stemness. Cancers 2022, 14, 4280. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, L.N.; Wilkinson, K.H.; Kong, A. Triple-Negative Breast Cancer: Who Should Receive Neoadjuvant Chemotherapy? Surg. Oncol. Clin. N. Am. 2018, 27, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.R.; Chang, C.H.; Hsu, C.F.; Tsai, M.J.; Cheng, H.; Leong, M.K.; Sung, P.J.; Chen, J.C.; Weng, C.F. Natural compounds as potential adjuvants to cancer therapy: Preclinical evidence. Br. J. Pharmacol. 2020, 177, 1409–1423. [Google Scholar] [CrossRef]
- Nobili, S.; Lippi, D.; Witort, E.; Donnini, M.; Bausi, L.; Mini, E.; Capaccioli, S. Natural compounds for cancer treatment and prevention. Pharmacol. Res. 2009, 59, 365–378. [Google Scholar] [CrossRef]
- Liang, Z.; Xie, H.; Shen, W.; Shao, L.; Zeng, L.; Huang, X. The Synergism of Natural Compounds and Conventional Therapeutics against Colorectal Cancer Progression and Metastasis. Front. Biosci. Landmark Ed. 2022, 27, 263. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Gondal, T.A.; Saeed, F.; Imran, A.; Shahbaz, M.; Fokou, P.V.T.; Arshad, M.U.; Khan, H.; et al. Kaempferol: A Key Emphasis to Its Anticancer Potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomed. Pharmacother. 2019, 117, 109086. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Alarifi, A.; Karami, A.M.; Ayub, R.; Abduh, N.A.Y.; Saeed, W.S.; Muddassir, M. Antiproliferative Mechanisms of a Polyphenolic Combination of Kaempferol and Fisetin in Triple-Negative Breast Cancer Cells. Int. J. Mol. Sci. 2023, 24, 6393. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Chen, J.; Zheng, X.; Song, J.; Yin, L.; Guo, H.; Chen, Q.; Liu, Y.; Ma, Q.; Zhang, H.; et al. Kaempferol attenuates doxorubicin-induced renal tubular injury by inhibiting ROS/ASK1-mediated activation of the MAPK signaling pathway. Biomed. Pharmacother. 2023, 157, 114087. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Xing, J.; Aikemu, B.; Sun, J.; Zheng, M. Kaempferol exhibits a synergistic effect with doxorubicin to inhibit proliferation, migration, and invasion of liver cancer. Oncol. Rep. 2021, 45, 32. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Yang, B.; Qiao, Y.; Zhou, Q.; He, H.; He, M. Kaempferol protects mitochondria and alleviates damages against endotheliotoxicity induced by doxorubicin. Biomed. Pharmacother. 2020, 126, 110040. [Google Scholar] [CrossRef]
- Naeem, A.; Hu, P.; Yang, M.; Zhang, J.; Liu, Y.; Zhu, W.; Zheng, Q. Natural Products as Anticancer Agents: Current Status and Future Perspectives. Molecules 2022, 27, 8367. [Google Scholar] [CrossRef]
- Siegel, R.L.; Giaquinto, A.N.; Jemal, A. Cancer statistics, 2024. CA Cancer J. Clin. 2024, 74, 12–49. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Lehmann, B.D.; Bauer, J.A.; Chen, X.; Sanders, M.E.; Chakravarthy, A.B.; Shyr, Y.; Pietenpol, J.A. Identification of human triple-negative breast cancer subtypes and preclinical models for selection of targeted therapies. J. Clin. Investig. 2011, 121, 2750–2767. [Google Scholar] [CrossRef] [PubMed]
- Masuda, H.; Baggerly, K.A.; Wang, Y.; Zhang, Y.; Gonzalez-Angulo, A.M.; Meric-Bernstam, F.; Valero, V.; Lehmann, B.D.; Pietenpol, J.A.; Hortobagyi, G.N.; et al. Differential Response to Neoadjuvant Chemotherapy among 7 Triple-Negative Breast Cancer Molecular Subtypes. Clin. Cancer Res. 2013, 19, 5533–5540. [Google Scholar] [CrossRef] [PubMed]
- Kudelova, E.; Smolar, M.; Holubekova, V.; Hornakova, A.; Dvorska, D.; Lucansky, V.; Koklesova, L.; Kudela, E.; Kubatka, P. Genetic Heterogeneity, Tumor Microenvironment and Immunotherapy in Triple-Negative Breast Cancer. Int. J. Mol. Sci. 2022, 23, 14937. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Z.; Liu, S.; Sun, J.; Chen, Z.; Jiang, M.; Zhang, Q.; Wei, Y.; Wang, X.; Huang, Y.-Y.; et al. Potential therapeutic effects of dipyridamole in the severely ill patients with COVID-19. Acta Pharm. Sin. B 2020, 10, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Sauter, E.R. Cancer prevention and treatment using combination therapy with natural compounds. Expert Rev. Clin. Pharmacol. 2020, 13, 265–285. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, Y. Teaching an old dog new tricks: Drug discovery by repositioning natural products and their derivatives. Drug Discov. Today 2022, 27, 1936–1944. [Google Scholar] [CrossRef]
- Egbuna, C.; Kumar, S.; Ifemeje, J.C.; Ezzat, S.M.; Kaliyaperumal, S. Phytochemicals as Lead Compounds for New Drug Discovery; Elsevier: London, UK, 2019. [Google Scholar]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef]
- Periferakis, A.; Periferakis, K.; Badarau, I.A.; Petran, E.M.; Popa, D.C.; Caruntu, A.; Costache, R.S.; Scheau, C.; Caruntu, C.; Costache, D.O. Kaempferol: Antimicrobial Properties, Sources, Clinical, and Traditional Applications. Int. J. Mol. Sci. 2022, 23, 15054. [Google Scholar] [CrossRef]
- Alam, W.; Khan, H.; Shah, M.A.; Cauli, O.; Saso, L. Kaempferol as a Dietary Anti-Inflammatory Agent: Current Therapeutic Standing. Molecules 2020, 25, 4073. [Google Scholar] [CrossRef]
- Lee, H.S.; Cho, H.J.; Yu, R.; Lee, K.W.; Chun, H.S.; Park, J.H.Y. Mechanisms Underlying Apoptosis-Inducing Effects of Kaempferol in HT-29 Human Colon Cancer Cells. Int. J. Mol. Sci. 2014, 15, 2722–2737. [Google Scholar] [CrossRef]
- Rho, H.S.; Ghimeray, A.K.; Yoo, D.S.; Ahn, S.M.; Kwon, S.S.; Lee, K.H.; Cho, D.H.; Cho, J.Y. Kaempferol and Kaempferol Rhamnosides with Depigmenting and Anti-Inflammatory Properties. Molecules 2011, 16, 3338–3344. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhong, H.; Huang, Z.; Chen, X.; You, J.; Zou, T. A Critical Review of Kaempferol in Intestinal Health and Diseases. Antioxidants 2023, 12, 1642. [Google Scholar] [CrossRef]
- Fernández-Del-Río, L.; Soubeyrand, E.; Basset, G.J.; Clarke, C.F. Metabolism of the Flavonol Kaempferol in Kidney Cells Liberates the B-ring to Enter Coenzyme Q Biosynthesis. Molecules 2020, 25, 2955. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.Y.; Chen, Y.C. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013, 138, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Calderon-Montaño, J.M.; Burgos-Morón, E.; Perez-Guerrero, C.; Lopez-Lazaro, M. A Review on the Dietary Flavonoid Kaempferol. Mini-Reviews Med. Chem. 2011, 11, 298–344. [Google Scholar] [CrossRef] [PubMed]
- Ashrafizadeh, M.; Tavakol, S.; Ahmadi, Z.; Roomiani, S.; Mohammadinejad, R.; Samarghandian, S. Therapeutic effects of kaempferol affecting autophagy and endoplasmic reticulum stress. Phytotherapy Res. 2020, 34, 911–923. [Google Scholar] [CrossRef] [PubMed]
- Devi, K.P.; Malar, D.S.; Nabavi, S.F.; Sureda, A.; Xiao, J.; Nabavi, S.M.; Daglia, M. Kaempferol and inflammation: From chemistry to medicine. Pharmacol. Res. 2015, 99, 1–10. [Google Scholar] [CrossRef]
- Duan, L.; Rao, X.; Sigdel, K.R. Regulation of Inflammation in Autoimmune Disease. J. Immunol. Res. 2019, 2019, 7403796. [Google Scholar] [CrossRef]
- Lemos, C.; Peters, G.J.; Jansen, G.; Martel, F.; Calhau, C. Modulation of folate uptake in cultured human colon adenocarcinoma Caco-2 cells by dietary compounds. Eur. J. Nutr. 2007, 46, 329–336. [Google Scholar] [CrossRef]
- Balabhadrapathruni, S.; Thomas, T.J.; Yurkow, E.J.; Amenta, P.S.; Thomas, T. Effects of genistein and structurally related phytoestrogens on cell cycle kinetics and apoptosis in MDA-MB-468 human breast cancer cells. Oncol. Rep. 2000, 7, 3–12. [Google Scholar] [CrossRef]
- Lee, K.-H.; Tagahara, K.; Suzuki, H.; Wu, R.-Y.; Haruna, M.; Hall, I.H.; Huang, H.-C.; Ito, K.; Iida, T.; Lai, J.-S. Antitumor Agents. 49. Tricin,Kaempferol-3-0-β-D-Glucopyranoside and (+)-Nortrachelogenin, Antileukemic Principles from Wikstroemia indica. J. Nat. Prod. 1981, 44, 530–535. [Google Scholar] [CrossRef]
- Lin, J.K.; Chen, Y.C.; Huang, Y.T.; Lin-Shiau, S.Y. Suppression of protein kinase C and nuclear oncogene expression as possible molecular mechanisms of cancer chemoprevention by apigenin and curcumin. J. Cell. Biochem. Suppl. 1997, 28–29, 39–48. [Google Scholar] [CrossRef]
- Stoner, G.D.; Chen, T.; Kresty, L.A.; Aziz, R.M.; Reinemann, T.; Nines, R. Protection Against Esophageal Cancer in Rodents with Lyophilized Berries: Potential Mechanisms. Nutr. Cancer 2006, 54, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, A.; Brandis, D. Shock Resuscitation. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK534830/ (accessed on 11 December 2023).
- Niering, P.; Michels, G.; Wätjen, W.; Ohler, S.; Steffan, B.; Chovolou, Y.; Kampkötter, A.; Proksch, P.; Kahl, R. Protective and detrimental effects of kaempferol in rat H4IIE cells: Implication of oxidative stress and apoptosis. Toxicol. Appl. Pharmacol. 2005, 209, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, P.; Ramanathan, M. Effect of kaempferol on lipid peroxidation and antioxidant status in 1,2-dimethyl hydrazine induced colorectal carcinoma in rats. Eur. J. Pharmacol. 2011, 654, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H. A Protocol for Flavonols, Kaempferol and Quercetin, Staining in Plant Root Tips. Bio-Protocol 2020, 10, e3781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, A.Y.; Li, M.; Chen, C.; Yao, Q. Ginkgo biloba Extract Kaempferol Inhibits Cell Proliferation and Induces Apoptosis in Pancreatic Cancer Cells. J. Surg. Res. 2008, 148, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.W.; Kim, J.H.; Song, K.; Kim, S.H.; Yoon, J.; Kim, K. Kaempferol and quercetin, components of Ginkgo biloba extract (EGb 761), induce caspase-3-dependent apoptosis in oral cavity cancer cells. Phytotherapy Res. 2010, 24, S77–S82. [Google Scholar] [CrossRef]
- Wang, F.; Wang, L.; Qu, C.; Chen, L.; Geng, Y.; Cheng, C.; Yu, S.; Wang, D.; Yang, L.; Meng, Z.; et al. Kaempferol induces ROS-dependent apoptosis in pancreatic cancer cells via TGM2-mediated Akt/mTOR signaling. BMC Cancer 2021, 21, 396. [Google Scholar] [CrossRef]
- Hung, T.-W.; Chen, P.-N.; Wu, H.-C.; Wu, S.-W.; Tsai, P.-Y.; Hsieh, Y.-S.; Chang, H.-R. Kaempferol Inhibits the Invasion and Migration of Renal Cancer Cells through the Downregulation of AKT and FAK Pathways. Int. J. Med Sci. 2017, 14, 984–993. [Google Scholar] [CrossRef]
- Kashafi, E.; Moradzadeh, M.; Mohamadkhani, A.; Erfanian, S. Kaempferol increases apoptosis in human cervical cancer HeLa cells via PI3K/AKT and telomerase pathways. Biomed. Pharmacother. 2017, 89, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.-B.; Kim, J.-H.; Lee, H.; Pak, J.-N.; Shim, B.S.; Kim, S.-H. Reactive Oxygen Species and p53 Mediated Activation of p38 and Caspases is Critically Involved in Kaempferol Induced Apoptosis in Colorectal Cancer Cells. J. Agric. Food Chem. 2018, 66, 9960–9967. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Wang, X.; Li, C.; Zhao, T.; Jin, H.; Fang, W. Kaempferol inhibits cell proliferation and glycolysis in esophagus squamous cell carcinoma via targeting EGFR signaling pathway. Tumor Biol. 2016, 37, 10247–10256. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Dang, Q.; Xu, D.; Chen, Y.; Zhu, G.; Wu, K.; Zeng, J.; Long, Q.; Wang, X.; He, D.; et al. Kaempferol induces cell cycle arrest and apoptosis in renal cell carcinoma through EGFR/p38 signaling. Oncol. Rep. 2014, 31, 1350–1356. [Google Scholar] [CrossRef]
- Gao, Y.; Yin, J.; Rankin, G.O.; Chen, Y.C. Kaempferol Induces G2/M Cell Cycle Arrest via Checkpoint Kinase 2 and Promotes Apoptosis via Death Receptors in Human Ovarian Carcinoma A2780/CP70 Cells. Molecules 2018, 23, 1095. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Jiang, B.; Li, B.; Li, Z.; Jiang, B.-H.; Chen, Y.C. Kaempferol nanoparticles achieve strong and selective inhibition of ovarian cancer cell viability. Int. J. Nanomed. 2012, 7, 3951–3959. [Google Scholar] [CrossRef]
- Chin, H.; Horng, C.; Liu, Y.; Lu, C.; Su, C.; Chen, P.; Chiu, H.; Tsai, F.; Shieh, P.; Yang, J. Kaempferol inhibits angiogenic ability by targeting VEGF receptor-2 and downregulating the PI3K/AKT, MEK and ERK pathways in VEGF-stimulated human umbilical vein endothelial cells. Oncol. Rep. 2018, 39, 2351–2357. [Google Scholar] [CrossRef]
- Ju, P.; Ho, Y.; Chen, P.; Lee, H.; Lai, S.; Yang, S.; Yeh, C. Kaempferol inhibits the cell migration of human hepatocellular carcinoma cells by suppressing MMP-9 and Akt signaling. Environ. Toxicol. 2021, 36, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Čipák, L.; Novotný, L.; Čipáková, I.; Rauko, P. Differential modulation of cisplatin and doxorubicin efficacies in leukemia cells by flavonoids. Nutr. Res. 2003, 23, 1045–1057. [Google Scholar] [CrossRef]
- Luo, H.; Daddysman, M.K.; O Rankin, G.; Jiang, B.-H.; Chen, Y.C. Kaempferol enhances cisplatin’s effect on ovarian cancer cells through promoting apoptosis caused by down regulation of cMyc. Cancer Cell Int. 2010, 10, 16. [Google Scholar] [CrossRef]
- Zhou, Q.; Fang, G.; Pang, Y.; Wang, X. Combination of Kaempferol and Docetaxel Induces Autophagy in Prostate Cancer Cells In Vitro and In Vivo. Int. J. Mol. Sci. 2023, 24, 14519. [Google Scholar] [CrossRef] [PubMed]
- Al-Nour, M.Y.; Ibrahim, M.M.; Elsaman, T. Ellagic Acid, Kaempferol, and Quercetin from Acacia nilotica: Promising Combined Drug with Multiple Mechanisms of Action. Curr. Pharmacol. Rep. 2019, 5, 255–280. [Google Scholar] [CrossRef] [PubMed]
- To, K.K.; Cho, W.C. Flavonoids Overcome Drug Resistance to Cancer Chemotherapy by Epigenetically Modulating Multiple Mechanisms. Curr. Cancer Drug Targets 2021, 21, 289–305. [Google Scholar] [CrossRef]
- Riahi-Chebbi, I.; Souid, S.; Othman, H.; Haoues, M.; Karoui, H.; Morel, A.; Srairi-Abid, N.; Essafi, M.; Essafi-Benkhadir, K. The Phenolic compound Kaempferol overcomes 5-fluorouracil resistance in human resistant LS174 colon cancer cells. Sci. Rep. 2019, 9, 195. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Du, J.; Li, C.; Li, H.; Guo, H.; Li, Z. Kaempferol Can Reverse the 5-Fu Resistance of Colorectal Cancer Cells by Inhibiting PKM2-Mediated Glycolysis. Int. J. Mol. Sci. 2022, 23, 3544. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Lee, G.-E.; An, H.-J.; Lee, C.-J.; Cho, E.S.; Kang, H.C.; Lee, J.Y.; Lee, H.S.; Choi, J.-S.; Kim, D.J.; et al. Kaempferol sensitizes cell proliferation inhibition in oxaliplatin-resistant colon cancer cells. Arch. Pharmacal Res. 2021, 44, 1091–1108. [Google Scholar] [CrossRef] [PubMed]
- Nandi, S.K.; Pradhan, A.; Das, B.; Das, B.; Basu, S.; Mallick, B.; Dutta, A.; Sarkar, D.K.; Mukhopadhyay, A.; Mukhopadhyay, S.; et al. Kaempferol attenuates viability of ex-vivo cultured post-NACT breast tumor explants through downregulation of p53 induced stemness, inflammation and apoptosis evasion pathways. Pathol.-Res. Pr. 2022, 237, 154029. [Google Scholar] [CrossRef] [PubMed]
- Adebamowo, C.A.; Cho, E.; Sampson, L.; Katan, M.B.; Spiegelman, D.; Willett, W.C.; Holmes, M.D. Dietary flavonols and flavonol-rich foods intake and the risk of breast cancer. Int. J. Cancer 2005, 114, 628–633. [Google Scholar] [CrossRef]
- Touillaud, M.S.; Pillow, P.C.; Jakovljevic, J.; Bondy, M.L.; Singletary, S.E.; Li, D.; Chang, S. Effect of Dietary Intake of Phytoestrogens on Estrogen Receptor Status in Premenopausal Women with Breast Cancer. Nutr. Cancer 2005, 51, 162–169. [Google Scholar] [CrossRef]
- Oh, S.M.; Kim, Y.P.; Chung, K.H. Biphasic effects of kaempferol on the estrogenicity in human breast cancer cells. Arch. Pharmacal Res. 2006, 29, 354–362. [Google Scholar] [CrossRef]
- Zhu, L.; Xue, L. Kaempferol Suppresses Proliferation and Induces Cell Cycle Arrest, Apoptosis, and DNA Damage in Breast Cancer Cells. Oncol. Res. Featur. Preclin. Clin. Cancer Ther. 2019, 27, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Wang, J.; Li, Y.; Fan, P.; Liu, G.; Zhang, N.; Conaway, M.; Wang, H.; Korach, K.S.; Bocchinfuso, W.; et al. Effects of estrogen on breast cancer development: Role of estrogen receptor independent mechanisms. Int. J. Cancer 2010, 127, 1748–1757. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kurzer, M.S. Phytoestrogen concentration determines effects on DNA synthesis in human breast cancer cells. Nutr. Cancer 1997, 28, 236–247. [Google Scholar] [CrossRef] [PubMed]
- Zava, D.T.; Duwe, G. Estrogenic and antiproliferative properties of genistein and other flavonoids in human breast cancer cells in vitro. Nutr. Cancer 1997, 27, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Hung, H. Inhibition of estrogen receptor alpha expression and function in MCF-7 cells by kaempferol. J. Cell. Physiol. 2004, 198, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Hwang, K.-A.; Choi, K.-C. Treatment with kaempferol suppresses breast cancer cell growth caused by estrogen and triclosan in cellular and xenograft breast cancer models. J. Nutr. Biochem. 2016, 28, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.-A.; Choi, K.-C.; Hwang, K.-A. Kaempferol, a phytoestrogen, suppressed triclosan-induced epithelial-mesenchymal transition and metastatic-related behaviors of MCF-7 breast cancer cells. Environ. Toxicol. Pharmacol. 2017, 49, 48–57. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-Salinas, F.L.; Delgado-Magallón, A.; Montes-Alvarado, J.B.; Ramírez-Ramírez, D.; Flores-Alonso, J.C.; Cortés-Hernández, P.; Reyes-Leyva, J.; Herrera-Camacho, I.; Anaya-Ruiz, M.; Pelayo, R.; et al. Breast Cancer Subtypes Present a Differential Production of Reactive Oxygen Species (ROS) and Susceptibility to Antioxidant Treatment. Front. Oncol. 2019, 9, 480. [Google Scholar] [CrossRef] [PubMed]
- Oshi, M.; Gandhi, S.; Yan, L.; Tokumaru, Y.; Wu, R.; Yamada, A.; Matsuyama, R.; Endo, I.; Takabe, K. Abundance of reactive oxygen species (ROS) is associated with tumor aggressiveness, immune response, and worse survival in breast cancer. Breast Cancer Res. Treat. 2022, 194, 231–241. [Google Scholar] [CrossRef]
- Raza, M.H.; Siraj, S.; Arshad, A.; Waheed, U.; Aldakheel, F.; Alduraywish, S.; Arshad, M. ROS-modulated therapeutic approaches in cancer treatment. J. Cancer Res. Clin. Oncol. 2017, 143, 1789–1809. [Google Scholar] [CrossRef]
- Okoh, V.; Deoraj, A.; Roy, D. Estrogen-induced reactive oxygen species-mediated signalings contribute to breast cancer. Biochim. Biophys. Acta (BBA)—Rev. Cancer 2011, 1815, 115–133. [Google Scholar] [CrossRef] [PubMed]
- Felty, Q.; Xiong, W.-C.; Sun, D.; Sarkar, S.; Singh, K.P.; Parkash, J.; Roy, D. Estrogen-Induced Mitochondrial Reactive Oxygen Species as Signal-Transducing Messengers. Biochemistry 2005, 44, 6900–6909. [Google Scholar] [CrossRef]
- Liao, W.; Chen, L.; Ma, X.; Jiao, R.; Li, X.; Wang, Y. Protective effects of kaempferol against reactive oxygen species-induced hemolysis and its antiproliferative activity on human cancer cells. Eur. J. Med. Chem. 2016, 114, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Xu, H.; Fan, P.-Z.; Xie, J.; He, J.; Yu, J.; Gu, X.; Zhang, C.-J. Kaempferol blocks neutrophil extracellular traps formation and reduces tumour metastasis by inhibiting ROS-PAD4 pathway. J. Cell. Mol. Med. 2020, 24, 7590–7599. [Google Scholar] [CrossRef]
- Kim, B.-W.; Lee, E.-R.; Min, H.-M.; Jeong, H.-S.; Ahn, J.-Y.; Kim, J.-H.; Choi, H.-Y.; Choi, H.; Kim, E.Y.; Park, S.P.; et al. Sustained ERK activation is involved in the kaempferol-induced apoptosis of breast cancer cells and is more evident under 3-D culture condition. Cancer Biol. Ther. 2008, 7, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Menegon, S.; Columbano, A.; Giordano, S. The Dual Roles of NRF2 in Cancer. Trends Mol. Med. 2016, 22, 578–593. [Google Scholar] [CrossRef]
- Sajadimajd, S.; Khazaei, M. Oxidative Stress and Cancer: The Role of Nrf2. Curr. Cancer Drug Targets 2018, 18, 538–557. [Google Scholar] [CrossRef]
- Telkoparan-Akillilar, P.; Panieri, E.; Cevik, D.; Suzen, S.; Saso, L. Therapeutic Targeting of the NRF2 Signaling Pathway in Cancer. Molecules 2021, 26, 1417. [Google Scholar] [CrossRef] [PubMed]
- De La Vega, M.R.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef]
- Fouzder, C.; Mukhuty, A.; Kundu, R. Kaempferol inhibits Nrf2 signalling pathway via downregulation of Nrf2 mRNA and induces apoptosis in NSCLC cells. Arch. Biochem. Biophys. 2021, 697, 108700. [Google Scholar] [CrossRef]
- Li, S.; Yan, T.; Deng, R.; Jiang, X.; Xiong, H.; Wang, Y.; Yu, Q.; Wang, X.; Chen, C.; Zhu, Y. Low dose of kaempferol suppresses the migration and invasion of triple-negative breast cancer cells by downregulating the activities of RhoA and Rac1. OncoTargets Ther. 2017, 10, 4809–4819. [Google Scholar] [CrossRef] [PubMed]
- Hall, R.; Birrell, S.; Tilley, W.; Sutherland, R. MDA-MB-453, an androgen-responsive human breast carcinoma cell line with high level androgen receptor expression. Eur. J. Cancer 1994, 30, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.J.; Ahn, W.S. Kaempferol induced the apoptosis via cell cycle arrest in human breast cancer MDA-MB-453 cells. Nutr. Res. Pr. 2008, 2, 322–325. [Google Scholar] [CrossRef] [PubMed]
- Kaur, V.; Kumar, M.; Kumar, A.; Kaur, S. Butea monosperma (Lam.) Taub. Bark fractions protect against free radicals and induce apoptosis in MCF-7 breast cancer cells via cell-cycle arrest and ROS-mediated pathway. Drug Chem. Toxicol. 2020, 43, 398–408. [Google Scholar] [CrossRef]
- Brusselmans, K.; Vrolix, R.; Verhoeven, G.; Swinnen, J.V. Induction of Cancer Cell Apoptosis by Flavonoids Is Associated with Their Ability to Inhibit Fatty Acid Synthase Activity. J. Biol. Chem. 2005, 280, 5636–5645. [Google Scholar] [CrossRef] [PubMed]
- Diantini, A.; Subarnas, A.; Lestari, K.; Halimah, E.; Susilawati, Y.; Supriyatna; Julaeha, E.; Achmad, T.H.; Suradji, E.W.; Yamazaki, C.; et al. Kaempferol-3-O-rhamnoside isolated from the leaves of Schima wallichii Korth. inhibits MCF-7 breast cancer cell proliferation through activation of the caspase cascade pathway. Oncol. Lett. 2012, 3, 1069–1072. [Google Scholar] [CrossRef] [PubMed]
- Radhika, M.; Ghoshal, N.; Chatterjee, A. Comparison of effectiveness in antitumor activity between flavonoids and polyphenols of the methanolic extract of roots of Potentilla fulgens in breast cancer cells. J. Complement. Integr. Med. 2012, 9, Article 24. [Google Scholar] [CrossRef] [PubMed]
- Tor, Y.S.; Yazan, L.S.; Foo, J.B.; Wibowo, A.; Ismail, N.; Cheah, Y.K.; Abdullah, R.; Ismail, M.; Ismail, I.S.; Yeap, S.K. Induction of Apoptosis in MCF-7 Cells via Oxidative Stress Generation, Mitochondria-Dependent and Caspase-Independent Pathway by Ethyl Acetate Extract of Dillenia suffruticosa and Its Chemical Profile. PLOS ONE 2015, 10, e0127441. [Google Scholar] [CrossRef]
- Shoja, M.; Reddy, N.D.; Nayak, P.G.; Srinivasan, K.; Rao, C.M. Glycosmis pentaphylla (Retz.) DC arrests cell cycle and induces apoptosis via caspase-3/7 activation in breast cancer cells. J. Ethnopharmacol. 2015, 168, 50–60. [Google Scholar] [CrossRef]
- Yi, X.; Zuo, J.; Tan, C.; Xian, S.; Luo, C.; Chen, S. Kaempferol, a flavonoid compound from GYNURA MEDICA induced apoptosis and growth inhibition in mcf-7 breast cancer cell. Afr. J. Tradit. Complement. Altern. Med. AJTCAM 2016, 13, 210–215. [Google Scholar] [CrossRef]
- Kang, G.-Y.; Lee, E.-R.; Kim, J.-H.; Jung, J.W.; Lim, J.; Kim, S.K.; Cho, S.-G.; Kim, K.P. Downregulation of PLK-1 expression in kaempferol-induced apoptosis of MCF-7 cells. Eur. J. Pharmacol. 2009, 611, 17–21. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, F.; Chandrashekar, D.S.; Varambally, S.; Creighton, C.J. Proteogenomic characterization of 2002 human cancers reveals pan-cancer molecular subtypes and associated pathways. Nat. Commun. 2022, 13, 2669. [Google Scholar] [CrossRef] [PubMed]
- Phromnoi, K.; Yodkeeree, S.; Anuchapreeda, S.; Limtrakul, P. Inhibition of MMP-3 activity and invasion of the MDA-MB-231 human invasive breast carcinoma cell line by bioflavonoids. Acta Pharmacol. Sin. 2009, 30, 1169–1176. [Google Scholar] [CrossRef] [PubMed]
- Astin, J.W.; Jamieson, S.M.; Eng, T.C.; Flores, M.V.; Misa, J.P.; Chien, A.; Crosier, K.E.; Crosier, P.S. An In Vivo Antilymphatic Screen in Zebrafish Identifies Novel Inhibitors of Mammalian Lymphangiogenesis and Lymphatic-Mediated Metastasis. Mol. Cancer Ther. 2014, 13, 2450–2462. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Zhao, Y.; Yang, D.; Yu, Y.; Guo, H.; Zhao, Z.; Zhang, B.; Yin, X. Inhibitory effects of kaempferol on the invasion of human breast carcinoma cells by downregulating the expression and activity of matrix metalloproteinase-9. Biochem. Cell Biol. 2015, 93, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Izzo, S.; Naponelli, V.; Bettuzzi, S. Flavonoids as Epigenetic Modulators for Prostate Cancer Prevention. Nutrients 2020, 12, 1010. [Google Scholar] [CrossRef] [PubMed]
- Bouyahya, A.; Mechchate, H.; Oumeslakht, L.; Zeouk, I.; Aboulaghras, S.; Balahbib, A.; Zengin, G.; Kamal, M.A.; Gallo, M.; Montesano, D.; et al. The Role of Epigenetic Modifications in Human Cancers and the Use of Natural Compounds as Epidrugs: Mechanistic Pathways and Pharmacodynamic Actions. Biomolecules 2022, 12, 367. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Venturelli, S.; Kallnischkies, M.; Böcker, A.; Busch, C.; Weiland, T.; Noor, S.; Leischner, C.; Weiss, T.S.; Lauer, U.M.; et al. Kaempferol, a new nutrition-derived pan-inhibitor of human histone deacetylases. J. Nutr. Biochem. 2013, 24, 977–985. [Google Scholar] [CrossRef]
- Sharma, A.; Sinha, S.; Keswani, H.; Shrivastava, N. Kaempferol and Apigenin suppresses the stemness properties of TNBC cells by modulating Sirtuins. Mol. Divers. 2022, 26, 3225–3240. [Google Scholar] [CrossRef]
- Zamarron, B.F.; Chen, W. Dual Roles of Immune Cells and Their Factors in Cancer Development and Progression. Int. J. Biol. Sci. 2011, 7, 651–658. [Google Scholar] [CrossRef]
- Tiwari, A.; Trivedi, R.; Lin, S.-Y. Tumor microenvironment: Barrier or opportunity towards effective cancer therapy. J. Biomed. Sci. 2022, 29, 83. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, Y.; Du, W.; Gong, L.; Chang, H.; Zou, Z. Tumor-associated macrophages: An accomplice in solid tumor progression. J. Biomed. Sci. 2019, 26, 78. [Google Scholar] [CrossRef] [PubMed]
- Qiu, X.; Zhao, T.; Luo, R.; Qiu, R.; Li, Z. Tumor-Associated Macrophages: Key Players in Triple-Negative Breast Cancer. Front. Oncol. 2022, 12, 772615. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Shi, W.; Xu, Y.; Xu, C.; Zhao, T.; Geng, B.; Yang, J.; Pan, J.; Hu, S.; Zhang, C.; et al. Tumor-derived lactate induces M2 macrophage polarization via the activation of the ERK/STAT3 signaling pathway in breast cancer. Cell Cycle 2018, 17, 428–438. [Google Scholar] [CrossRef]
- Yoshimur, T.; Li, C.; Wang, Y.; Matsukawa, A. The Chemokine Monocyte Chemoattractant Protein 1/CCL2 Is a Promoter of Breast Cancer Metastasis | Cellular & Molecular Immunology. Available online: https://www.nature.com/articles/s41423-023-01013-0 (accessed on 10 June 2024).
- Lafta, H.A.; Hussein, A.H.A.; Al-Shalah, S.A.J.; Alnassar, Y.S.; Mohammed, N.M.; Akram, S.M.; Qasim, M.T.; Najafi, M. Tumor-associated Macrophages (TAMs) in Cancer Resistance; Modulation by Natural Products—PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/36722486/ (accessed on 10 June 2024).
- Malla, R.; Padmaraju, V.; Kundrapu, D.B. Tumor-associated macrophages: Potential target of natural compounds for management of breast cancer. Life Sci. 2022, 301, 120572. [Google Scholar] [CrossRef] [PubMed]
- Sonoki, H.; Tanimae, A.; Endo, S.; Matsunaga, T.; Furuta, T.; Ichihara, K.; Ikari, A. Kaempherol and Luteolin Decrease Claudin-2 Expression Mediated by Inhibition of STAT3 in Lung Adenocarcinoma A549 Cells. Nutrients 2017, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Wonganan, O.; He, Y.-J.; Shen, X.-F.; Wongkrajang, K.; Suksamrarn, A.; Zhang, G.-L.; Wang, F. 6-Hydroxy-3-O-methyl-kaempferol 6-O-glucopyranoside potentiates the anti-proliferative effect of interferon α/β by promoting activation of the JAK/STAT signaling by inhibiting SOCS3 in hepatocellular carcinoma cells. Toxicol. Appl. Pharmacol. 2017, 336, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guo, Y.; Chen, M.; Chen, F.; Liu, B.; Shen, C. Kaempferol potentiates the sensitivity of pancreatic cancer cells to erlotinib via inhibition of the PI3K/AKT signaling pathway and epidermal growth factor receptor. Inflammopharmacology 2021, 29, 1587–1601. [Google Scholar] [CrossRef]
- Yu, Q.; Zeng, K.; Ma, X.; Song, F.; Jiang, Y.; Tu, P.; Wang, X. Resokaempferol-mediated anti-inflammatory effects on activated macrophages via the inhibition of JAK2/STAT3, NF-κB and JNK/p38 MAPK signaling pathways. Int. Immunopharmacol. 2016, 38, 104–114. [Google Scholar] [CrossRef]
- Fang, W.B.; Yao, M.; Brummer, G.; Acevedo, D.; Alhakamy, N.; Berkland, C.; Cheng, N. Targeted gene silencing of CCL2 inhibits triple negative breast cancer progression by blocking cancer stem cell renewal and M2 macrophage recruitment. Oncotarget 2016, 7, 49349–49367. [Google Scholar] [CrossRef]
- Mohamed, H.T.; El-Ghonaimy, E.A.; El-Shinawi, M.; Hosney, M.; Götte, M.; Woodward, W.A.; El-Mamlouk, T.; Mohamed, M.M. IL-8 and MCP-1/CCL2 regulate proteolytic activity in triple negative inflammatory breast cancer a mechanism that might be modulated by Src and Erk1/2. Toxicol. Appl. Pharmacol. 2020, 401, 115092. [Google Scholar] [CrossRef] [PubMed]
- Hao, Q.; Vadgama, J.V.; Wang, P. CCL2/CCR2 signaling in cancer pathogenesis. Cell Commun. Signal. 2020, 18, 82. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Yang, M.; Yin, J.; Li, P.; Zeng, S.; Zheng, G.; He, Z.; Liu, H.; Wang, Q.; Zhang, F.; et al. Tumor-associated macrophages promote epithelial–mesenchymal transition and the cancer stem cell properties in triple-negative breast cancer through CCL2/AKT/β-catenin signaling. Cell Commun. Signal. 2022, 20, 92. [Google Scholar] [CrossRef] [PubMed]
- Fridlender, Z.G.; Buchlis, G.; Kapoor, V.; Cheng, G.; Sun, J.; Singhal, S.; Crisanti, M.C.; Wang, L.-C.S.; Heitjan, D.; Snyder, L.A.; et al. CCL2 Blockade Augments Cancer Immunotherapy. Cancer Res. 2010, 70, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, H.J.; Yoon, S.; Ryu, H.M.; Lee, E.; Jo, Y. Blockade of CCL2 expression overcomes intrinsic PD-1/PD-L1 inhibitor-resistance in transglutaminase 2-induced PD-L1 positive triple negative breast cancer. Am. J. Cancer Res. 2020, 10, 2878–2894. [Google Scholar] [PubMed]
- Mendonca, P.; Hilliard, A.; Soliman, K. The Inhibitory Effects of Ganoderma lucium on Cell Proliferation, Apoptosis, and TNF-α-Induced CCL2 Release in Genetically Different Triple-Negative Breast Cancer Cells. FASEB J. 2021, 35, S1. [Google Scholar] [CrossRef]
- Kanga, K.J.; Mendonca, P.; Soliman, K.F.; Ferguson, D.T.; Darling-Reed, S.F. Effect of Diallyl Trisulfide on TNF-α-induced CCL2/MCP-1 Release in Genetically Different Triple-negative Breast Cancer Cells. Anticancer. Res. 2021, 41, 5919–5933. [Google Scholar] [CrossRef] [PubMed]
- Messeha, S.S.; Zarmouh, N.O.; Mendonca, P.; Alwagdani, H.; Kolta, M.G.; Soliman, K.F.A. The inhibitory effects of plumbagin on the NF-κB pathway and CCL2 release in racially different triple-negative breast cancer cells. PLOS ONE 2018, 13, e0201116. [Google Scholar] [CrossRef] [PubMed]
- Joshi, N.; Tripathi, D.K.; Nagar, N.; Poluri, K.M. Hydroxyl Groups on Annular Ring-B Dictate the Affinities of Flavonol–CCL2 Chemokine Binding Interactions. ACS Omega 2021, 6, 10306–10317. [Google Scholar] [CrossRef]
- Tzeng, C.-W.; Yen, F.-L.; Wu, T.-H.; Ko, H.-H.; Lee, C.-W.; Tzeng, W.-S.; Lin, C.-C. Enhancement of Dissolution and Antioxidant Activity of Kaempferol Using a Nanoparticle Engineering Process. J. Agric. Food Chem. 2011, 59, 5073–5080. [Google Scholar] [CrossRef]
- Kazmi, I.; Al-Abbasi, F.A.; Afzal, M.; Altayb, H.N.; Nadeem, M.S.; Gupta, G. Formulation and Evaluation of Kaempferol Loaded Nanoparticles against Experimentally Induced Hepatocellular Carcinoma: In Vitro and In Vivo Studies. Pharmaceutics 2021, 13, 2086. [Google Scholar] [CrossRef] [PubMed]
- Raghavan, B.S.; Kondath, S.; Anantanarayanan, R.; Rajaram, R. Kaempferol mediated synthesis of gold nanoparticles and their cytotoxic effects on MCF-7 cancer cell line. Process. Biochem. 2015, 50, 1966–1976. [Google Scholar] [CrossRef]
- Govindaraju, S.; Roshini, A.; Lee, M.-H.; Yun, K. Kaempferol conjugated gold nanoclusters enabled efficient for anticancer therapeutics to A549 lung cancer cells. Int. J. Nanomed. 2019, 14, 5147–5157. [Google Scholar] [CrossRef]
- Colombo, M.; Melchiades, G.d.L.; Michels, L.R.; Figueiró, F.; Bassani, V.L.; Teixeira, H.F.; Koester, L.S. Solid Dispersion of Kaempferol: Formulation Development, Characterization, and Oral Bioavailability Assessment. Aaps Pharmscitech 2019, 20, 106. [Google Scholar] [CrossRef]
- Du, Q.; Chen, J.; Yan, G.; Lyu, F.; Huang, J.; Ren, J.; Di, L. Comparison of different aliphatic acid grafted N-trimethyl chitosan surface-modified nanostructured lipid carriers for improved oral kaempferol delivery. Int. J. Pharm. 2019, 568, 118506. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Nathani, A.; Singh, M.; Kaur, S.; Nathani, A.; Singh, M.; Kaur, S.; Nathani, A.; Singh, M. Exosomal delivery of cannabinoids against cancer. Cancer Lett. 2023, 566, 216243. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, M.; Mizokami, T.; Ito, H.; Ikeda, Y. A randomized, placebo-controlled trial evaluating the safety of excessive administration of kaempferol aglycone. Food Sci. Nutr. 2023, 11, 5427–5437. [Google Scholar] [CrossRef] [PubMed]
- Navarro, S.L.; Schwarz, Y.; Song, X.; Wang, C.-Y.; Chen, C.; Trudo, S.P.; Kristal, A.R.; Kratz, M.; Eaton, D.L.; Lampe, J.W. Cruciferous Vegetables Have Variable Effects on Biomarkers of Systemic Inflammation in a Randomized Controlled Trial in Healthy Young Adults. J. Nutr. 2014, 144, 1850–1857. [Google Scholar] [CrossRef]
- Nejabati, H.R.; Roshangar, L. Kaempferol: A potential agent in the prevention of colorectal cancer. Physiol. Rep. 2022, 10, e15488. [Google Scholar] [CrossRef] [PubMed]
- Noer, J.B.; Hørsdal, O.K.; Xiang, X.; Luo, Y.; Regenberg, B. Extrachromosomal circular DNA in cancer: History, current knowledge, and methods. Trends Genet. 2022, 38, 766–781. [Google Scholar] [CrossRef]
- Li, R.; Wang, Y.; Li, J.; Zhou, X. Extrachromosomal circular DNA (eccDNA): An emerging star in cancer. Biomark. Res. 2022, 10, 53. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Huang, W.; Cui, L.; Zhu, P.; Lin, Q.; Zhang, W.; Li, J.; Deng, C.; Wu, Z.; Huang, Z.; et al. The landscape of extrachromosomal circular DNA (eccDNA) in the normal hematopoiesis and leukemia evolution. Cell Death Discov. 2022, 8, 1–10. [Google Scholar] [CrossRef]
- Wu, S.; Tao, T.; Zhang, L.; Zhu, X.; Zhou, X. Extrachromosomal DNA (ecDNA): Unveiling its role in cancer progression and implications for early detection. Heliyon 2023, 9, e21327. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Veroniaina, H.; Su, N.; Sha, K.; Jiang, F.; Wu, Z.; Qi, X. Applications and developments of gene therapy drug delivery systems for genetic diseases. Asian J. Pharm. Sci. 2021, 16, 687–703. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhang, H.; Zhou, Y.; Shi, J. Extrachromosomal circular DNA: A new potential role in cancer progression. J. Transl. Med. 2021, 19, 257. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Mo, Y.-Y. Role of microRNAs in breast cancer. Cancer Biol. Ther. 2013, 14, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.P.; Pérez-Moreno, P.; Pérez, Y.; Calaf, G.M. The Role of MicroRNAs in Breast Cancer and the Challenges of Their Clinical Application. Diagnostics 2023, 13, 3072. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Sasi, A.K.; Abak, A.; Shoorei, H.; Khoshkar, A.; Taheri, M. Contribution of miRNAs in the Pathogenesis of Breast Cancer. Front. Oncol. 2021, 11, 768949. [Google Scholar] [CrossRef]
- Gutierrez-Uribe, J.A.; Salinas-Santander, M.; Serna-Guerrero, D.; Serna-Saldivar, S.R.O.; Rivas-Estilla, A.M.; Rios-Ibarra, C.P. Inhibition of miR31 and miR92a as Oncological Biomarkers in RKO Colon Cancer Cells Treated with Kaempferol-3-O-Glycoside Isolated from Black Bean. J. Med. Food 2020, 23, 50–55. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Pathway Targeted | Outcomes |
|---|---|---|
| DNA Synthesis Inhibition | ||
| Zava et al., 2023 [78] | DNA synthesis inhibition and cell growth | Kaempferol demonstrated estrogen agonistic activity and showed cell growth and DNA inhibition (at 10 µM concentration, the 0.25 µg DNA in T47D cells). |
| Apoptosis | ||
| Balabhadrapathruni et al., 2000 [43] | Cell proliferation, cell cycle, and apoptosis | Kaempferol inhibits cellular proliferation by targeting the G2/M cell cycle and apoptosis in MDA-MB-468 cells. |
| Brusselmans et al., 2005 [102] | Fatty acid synthetase pathway | Kaempferol inhibited the fatty acid synthetase enzyme, which was overexpressed in human breast cancers, reduced cell growth, and increased apoptosis. |
| Kim et al., 2008 [89] | Cell viability and apoptosis in 2D and 3D cultures | Kaempferol-induced apoptosis by modulating the ERK/MEK1/ELK1 signaling pathway. |
| Targeting the Estrogenic pathway | ||
| Kim et al., 2016 [80] | E2-mediated breast cancer cell proliferation, cell cycle, and apoptosis | Kaempferol antagonized the Triclosan stimulated cell proliferation in MCF-7 cells, upregulated the expression of cathepsin, cyclin D1, and cyclin E, and downregulated the expression of Bax and p21. |
| Oh et al., 2006 [74] | Estrogen-dependent and estrogen-independent pathways in breast cancer cell proliferation and malignant cell transformation | Kaempferol inhibited cell proliferation via an estrogen-dependent pathway, preventing malignant transformation of the human breast cells. |
| Hung et al., 2004 [79] | Cell viability | Kaempferol reduced cell viability (IC50: 35.0 mM and 70 mM for ER-positive and ER-negative breast cancer cells, respectively), decreased ER-alpha mRNA and protein expression, and decreased the progesterone receptor, cyclin D1, and insulin receptor expression. |
| Anti-Oxidative stress | ||
| Zeng et al., 2020 [88] | Neutrophil extracellular traps, ROS | Kaempferol inhibited the formation of NETs, thereby reducing the formation of ROS and inhibiting metastasis in breast tumors. |
| Afzal et al., 2023 [16] | ROS, agonism with another potential anticancer agent: Fisetin, DNA damage, and apoptosis | Kaempferol synergized the anti-oxidative properties of fisetin and activated γ-H2AX, leading to DNA damage and apoptosis. |
| Wu et al., 2020 [19] | ROS, ROS-mediated cardiotoxicity, endotheliotoxicity | Kaempferol reversed the vascular toxicity and cardiotoxicity caused by doxorubicin’s adverse effects by modulating the levels of the 14-3-3 s protein, which regulates ROS levels. |
| Inhibition of Metastasis and Invasion | ||
| Li et al., 2017 [97] | Migration | Kaempferol inhibited the activation of RhoA, Rac, and GTP-binding proteins involved in microfilament arrangement, thereby inhibiting the cancer cell migration of MDA-MB-231 cells. |
| Li et al., 2015 [98] | TNBC cell adhesion, motility, and migration | Kaempferol (IC50: 204.7 mol/L) decreased the expression of MMP-2 and MMP-9 in MDA-MB-231 cells. |
| Cell Cycle Arrest | ||
| Choi et al., 2008 [100] | Cell cycle arrest | Kaempferol downregulated the levels of CCDK1, cyclin A, and cyclin B and induced cell cycle arrest at the G2/M phase of the cell cycle in MDA-MB-453 cells. |
| Varinder Kaur et al., 2018 [101] | Cell cycle arrest | Chloroform extract of Butea monosperma (Lam.) Taub bark rich in kaempferol produced cell cycle arrest at the G1 phase of the cell cycle in MCF-7 cells. |
| Zhu et al., 2019 [75] | Cell cycle arrest | Lower concentration of kaempferol is required to produce cell cycle arrest at G2/M phase in MDA-MB-231 (IC50: 43 µmol/L) cells than in estrogen receptor-positive BT474 breast cancer cells (IC50: 100 µmol/L). |
| Kim et al., 2016 [80] | Cell cycle arrest | Kaempferol downregulated the expression of cyclin D1 and cyclin E and upregulated the expression of p21 in triclosan-treated MCF-7 cell models. |
| Study | Formulation | Results |
|---|---|---|
| Luo et al., 2012 [60] | Prepared PEO-PPO-PEO nanoparticles, poly(lactic acid-co-glycolic acid) (PLGA) nanoparticles, PLGA polyethyleneimine (PEI), poly(amidoamine) (PAMAM) dendrimer, and glycol chitosan nanoparticles | Out of all, PEO-PPO-PEO nanoparticles of kaempferol (25 µM concentration of kaempferol) showed significant cytotoxicity towards A2780/CP70 and OVCAR-3 cancer cells in comparison to kaempferol alone. |
| Raghvan et al., 2015 [138] | Developed KAuNPs. | KAuNPs showed tremendous biocompatibility, demonstrated increased cytotoxic potential towards MCF-7 cells, and exhibited induction of apoptosis and anti-angiogenic activity compared to the individual compound. |
| Govindaraju et al., 2019 [139] | Developed kaempferol-conjugated gold nanoclusters (K-AuNCs). | K-AuNCs showed preferential nuclear localization and enhanced cytotoxicity towards A549 lung carcinoma cells compared to normal human cells. |
| Colombo et al., 2018 [140] | Developed kaempferol-loaded nanoemulsions, MNE (made with chitosan for mucoadhesion to achieve nasal delivery) and NE (made without chitosan), for the targeted delivery of kaempferol to the glioma cells. | Both ex vivo and in vivo studies revealed significantly higher permeation across the nasal mucosa with MNE while maintaining its antioxidant activities. They also showed higher antitumor activity against C6 glioma cells than NE and kaempferol alone. |
| Kazmi et al., 2021 [137] | Developed KFP-Np with HPMC-AS and Kollicoat MAE 30 DP using a quasi-emulsion solvent diffusion technique. | In their in vivo model of CdCl2-induced HCC, KFP-Np significantly improved liver function by lowering liver enzymes and oxidative stress markers, boosting physiological antioxidant enzymes, and reducing inflammatory markers. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, S.; Mendonca, P.; Soliman, K.F.A. The Anticancer Effects and Therapeutic Potential of Kaempferol in Triple-Negative Breast Cancer. Nutrients 2024, 16, 2392. https://doi.org/10.3390/nu16152392
Kaur S, Mendonca P, Soliman KFA. The Anticancer Effects and Therapeutic Potential of Kaempferol in Triple-Negative Breast Cancer. Nutrients. 2024; 16(15):2392. https://doi.org/10.3390/nu16152392
Chicago/Turabian StyleKaur, Sukhmandeep, Patricia Mendonca, and Karam F. A. Soliman. 2024. "The Anticancer Effects and Therapeutic Potential of Kaempferol in Triple-Negative Breast Cancer" Nutrients 16, no. 15: 2392. https://doi.org/10.3390/nu16152392