Abstract
Diabetes has become one of the most prevalent global epidemics, significantly impacting both the economy and the health of individuals. Diabetes is associated with numerous complications, such as obesity; hyperglycemia; hypercholesterolemia; dyslipidemia; metabolic endotoxemia; intestinal barrier damage; insulin-secretion defects; increased oxidative stress; and low-grade, systemic, and chronic inflammation. Diabetes cannot be completely cured; therefore, current research has focused on developing various methods to control diabetes. A promising strategy is the use of probiotics for diabetes intervention. Probiotics are a class of live, non-toxic microorganisms that can colonize the human intestine and help improve the balance of intestinal microbiota. In this review, we summarize the current clinical studies on using probiotics to control diabetes in humans, along with mechanistic studies conducted in animal models. The primary mechanism by which probiotics regulate diabetes is improved intestinal barrier integrity, alleviated oxidative stress, enhanced immune response, increased short-chain fatty acid production, etc. Therefore, probiotic supplementation holds great potential for the prevention and management of diabetes.
1. Introduction
1.1. Diabetes Mellitus
Diabetes mellitus is a chronic metabolic disease that is characterized by high blood sugar levels and circulation insulin, leading to significant morbidity and mortality worldwide [1]. According to the International Diabetes Federation Diabetes Atlas (9th edition), approximately 463 million adults globally suffer from diabetes. This number is projected to rise to 578 million (10.2% of the total population) by 2030 [2], making diabetes one of the most serious threats to human health in the 21st century. Diabetes mellitus is characterized by elevated blood glucose levels due to defects in insulin secretion and/or action [3]. Diabetes is classified into type 1 diabetes mellitus (T1DM), type 2 diabetes mellitus (T2DM), and gestational diabetes mellitus (GDM) based on genetics, etiology, and diagnostic criteria. The most common form of diabetes is T2DM, which accounts for 90% of global diabetes cases. Therefore, T2DM is the main target of diabetes prevention and treatment [4]. The main objective of this review is to discuss the effects of probiotics on T2DM and the underlying mechanisms.
The detrimental effects of diabetes have long been recognized. The main clinical symptoms of diabetes are increased food and water intake, increased urination, and body weight loss [4]. Diabetes can cause complications in multiple organs, such as the cardiovascular system, eyes, kidneys, and nerves, severely reducing the quality of life of patients. These complications have significant impacts on health—for example, significantly increased risk of heart disease and stroke [4,5]. Recent studies showed that the incidence and mortality of cardiovascular and cerebrovascular diseases in patients with diabetes were about 3.5-fold higher than those in non-diabetic patients. Diabetes patients are prone to hypertension, coronary heart disease, and myocardial infarction [6]. Diabetes can also cause serious damage to the glomeruli, leading to proteinuria, hypertension, a gradual decline in renal function, and potentially renal failure [7]. Moreover, diabetes can cause neuritis and peripheral neuropathy, resulting in foot ulcers and necrosis and sometimes necessitating amputation in severe cases [8]. In addition, diabetes can cause retinopathy, accompanied by the risk of blindness [9]. Recent research showed that patients with diabetes are at a higher risk of being infected with the 2019 novel coronavirus (COVID-19) [10,11]. Currently, the key strategies for managing diabetes include blood sugar monitoring, diet control, and exercise to maintain blood sugar in the appropriate range.
Currently, insulin injections, oral hypoglycemic drugs, and lifestyle management are the main treatments for diabetes [12]. However, a long-term regime of insulin injection may lead to insulin resistance, which may aggravate T2DM symptoms [13]. Moreover, T2DM is often associated with insulin resistance, making direct insulin injections less effective [14]. Hypoglycemic drugs may have side effects, including gastrointestinal discomfort and allergic reactions, as well as causing tissue damage to the liver, kidney, and nervous system [15]. Therefore, many researchers have been seeking treatments that have minimal side effects and can quickly and effectively control or even cure diabetes.
Besides innate genetic genes, certain unhealthy lifestyles are closely related to diabetes, including a lack of exercise, frequent intake of a high-sugar and high-fat diet, smoking, and alcohol abuse [16,17]. Overnutrition due to ingestion of a diet rich in sugar and fat is the most common cause of the development of diabetes. Intestinal microorganisms serve as a crucial link between diet and human health. Studies have shown that the metabolites produced by the host’s intestinal microorganisms are closely related to the risk of diabetes [18]. Therefore, probiotics can be used to regulate the structure and metabolism of intestinal flora when ingested as dietary supplements, thereby managing and inhibiting the development of diabetes.
1.2. Gut Microbiota
Recent epidemiological, physiological, and omics findings, along with cell-based and animal experimental results, indicate that a significant portion of the environmental impact on human health and disease risk may be mediated or modified by the microbial community [19]. These microbiotas include numerous interacting bacteria, archaea, phages, eukaryotic viruses, and fungi that coexist on the surface of the human body and in all body cavities, with most of them being mutually beneficial [19]. The number of genomes of all intestinal microbial genes in an individual is more than an order of magnitude greater than the human genome. Most of the microorganisms that inhabit the human body reside in the intestine and are influenced by lifestyle, drugs, and host genetics, especially dietary feeding during infancy. The intestinal microbiota regulates host immunity [20], digestive ability [21], intestinal endocrine function [22], neural signaling [23], drug metabolism [24], and the elimination of toxins by producing a variety of compounds that affect the host [25].
Much evidence has shown that gut microbiota influences the capacity of the distal intestine to secret hormones regulating blood glucose. Patients who have undergone complete colectomy have an increased risk of T2DM compared to those who do not have the surgery [26]. Mechanistic studies in mice have shown that hyperglycemia may increase intestinal barrier permeability by disrupting the tight junction integrity of glucose transporter 2 (GLUT2)-dependent intestinal epithelial cells, leading to mucosal leakage [27]. Therefore, there is significant interest in understanding whether abnormal gut microbiota contributes to the onset or maintenance of elevated blood glucose in T2DM and its precursor states.
1.3. Probiotics
Probiotics are live microorganisms that provide health benefits to the host when taken as dietary supplements. Probiotics can reduce the abundance of harmful bacteria in the intestinal flora and increase the abundance of beneficial bacteria, thereby regulating intestinal metabolism. In addition, probiotics increase the integrity of the intestinal barrier, thereby alleviating intestinal inflammation and reducing the risk of pathogen infection. When administered in adequate amounts (at least 106 CFU), probiotics can improve the balance of intestinal microorganisms and engage in the metabolism of the host [28]. In recent years, probiotics have been used to beneficially modulate the abundance of intestinal microbiota. According to a new classification of the genus previously known as Lactobacillus proposed in 2020 [29,30], new names were used when referring to the former genus Lactobacillus in this review.
Commonly reported probiotics include lactic acid bacteria (e.g., lactobacilli (formerly Lactobacillus), Bifidobacterium, Streptococcus), non-lactic acid-producing bacteria (e.g., Bacillus, Propionibacterium), non-pathogenic yeasts (e.g., Saccharomyces cerevisiae), and non-spore-forming and non-flagellated cocci [31]. Among them, lactobacilli and Bifidobacterium have been the most extensively studied extensively. Lactobacilli includes different species, among which the probiotics are Lab. acidophilus, Lbs. rhamnosus, Lab. delbrueckii subsp. bulgaricus, Lmb. reuteri, Lbs. casei, Lab. johnsonii, and Lpb. plantarum. Bifidobacterium belongs to the phylum Actinobacteria, with common probiotic species including B. animalis, B. bifidum, B. breve, B. infantis, B. lactis, and B. longum [32]. The ability of different probiotics to exert their biological activity in vivo depends on their specific properties, such as tolerance to acidic pH environments, resistance to digestion by bile and pancreatic juice, and high efficiency of colonization in the intestine [33].
Probiotics have been reported to offer numerous beneficial biological functions in the host intestine. For instance, probiotic supplementation may strengthen the junction of intestinal epithelial cells and improve the integrity of the gastric mucosal barrier, resulting in enhanced intestinal barrier function [28,34]. Moreover, probiotics can regulate intestinal motility via mutual communication between the probiotic flora in the intestine and the enteric nervous system, thereby regulating intestinal motility [35,36]. The use of specific probiotics, such as Lacticaseibacillus (Lbs.) rhamnosus CNCM I-3690, Lbs. rhamnosus GG, and Ultrabiotique [Lactobacillus (Lab.) acidophilus, B. lactis, Lactiplantibacillus (Lpb.) plantarum, and B. breve], can significantly reduce inflammation, improve colitis, and promote mucosal healing [37]. Probiotics [Lbs. rhamnosus CNCM I-3690, Lpb. plantarum MB452, Lpb. plantarum GOS42, and Limosilactobacillus (Lmb). fermentum GOS57] were also shown to reduce the risk of pathogen infection by enhancing the production of intestinal mucins [38,39]. In addition, probiotics (such as Lbs. rhamnosus JB-1, Lbs. rhamnosus GG, and Lpb. plantarum N14) activate the intestinal immune system by stimulating innate immune receptors (e.g., Toll-like receptors (TLRs) and C-type lectin receptors (CLRs)), which promotes the production of proinflammatory cytokines and stimulates macrophages to initiate phagocytosis [40]. When probiotics interact with other microbiota in the intestine, they engage in cross-feed and other interactions, thereby affecting the metabolic capacity of the host’s intestinal microbiota.
2. Negative Effects of Diabetes on the Gut Microbiome
Some bacterial genera are negatively correlated with type 2 diabetes. In the gut microbiota of T2DM patients, the abundance of Bacteroidetes, Bifidobacterium, Faecalibacterium, Akkermansia, and Roseburia was reduced. In contrast, the abundance of Fusobacterium, Ruminococcus, and Blautia in the gut microbiota of T2DM patients is higher than that in healthy controls [41]. Of note, T2DM can also lead to significant decreases in the abundance of Bacteroides in the intestine [42,43]. Patients with T2DM have a reduced abundance of Enterobacteriaceae, Bacteroides 20_3, and Bacteroides vulgaris [44,45,46]. Roseburia, Faecalibacterium, lactobacilli, Ruminococcus, and Blautia belong to the phylum Firmicutes and are negatively affected by diabetes [42,46,47]. Patients with T2DM generally have reduced levels of Roseburia as compared with their healthy cohorts [48]. R. intestinalis is positively associated with diabetes, while R. inulinivorans and Roseburia_272 are negatively associated with diabetes [44,46,49]. Other studies have observed a decreased abundance of Faecalibacterium and F. prausnitzii in T2DM patients. F. prausnitzii is a type of Gram-positive bacterium that can exert anti-inflammatory effects [47,50,51]. Lactobacilli species, such as Lab. acidophilus and Ligilactobacillus (Lgb). salivarius, are positively associated with T2DM. By contrast, Ligilactobacillus (Lgb). gasseri, another species within the genus lactobacilli, is inversely related to diabetes [42,44].
Patients with T2DM exhibit an imbalance in the gastrointestinal microbiota, characterized by an increase in the ratio of Firmicutes to Bacteroidetes and a decrease in lactic acid-producing species of the genera lactobacilli, Bifidobacterium, and Streptococcus [52]. These microbiotas have the potential to produce short-chain fatty acids (SCFAs) such as acetate. Acetate can be converted to butyrate through a cross-feeding mechanism [44,53,54]. SCFAs such as butyrate and propionate stimulate glucagon-like peptide-1 (GLP-1), an incretin hormone that regulates postprandial insulin secretion by increasing insulin release after glucose ingestion [55,56,57]. Moreover, SCFAs can regulate intestinal gluconeogenesis and glucose absorption into the portal vein [56,58]. In addition, T2DM may lead to an increase in pathogenic bacteria, including Enterobacteriaceae, in the gastrointestinal tract [52]. Dysbiosis and inflammation can weaken the intestinal barrier function, thereby increasing the risk of leaky gut syndrome [52].
In animal research, mice models are usually chosen because the intestinal structure of mice is very similar to that of humans. Researchers can induce T2DM in mice to closely observe the potential causal relationship and possible mechanisms between diabetes and intestinal microorganisms [59]. Previous studies showed that germ-free mice displayed a significant increase in adiposity and insulin resistance after receiving gut microbiota transplants from diabetic mice [60]. Consistent with human studies, diabetic rats undergoing Roux-en-Y gastric bypass (RYGB) surgery had altered gut microbiota, with an increased abundance of Bacteroidetes, Proteobacteria, Fusobacteria, and Actinobacteria and reduced levels of Firmicutes and Verrucomicrobia. RYGB surgery reduced body weight and significantly improved glucose tolerance and insulin sensitivity in diabetic rats [61]. These findings are comparable to the observations in human studies. Increased intestinal permeability in T2DM mice leads to higher levels of lipopolysaccharides (LPS) in the blood circulation, contributing to the progression of obesity and insulin resistance [62,63]. Therefore, diabetes may increase the abundance of harmful bacteria in the intestinal flora to aggravate intestinal inflammation and insulin resistance. Increasing the abundance of intestinal probiotics may alleviate diabetes.
3. Clinical Trial Study on the Use of Probiotics to Manage Diabetes in Humans
Some strains of lactobacilli, Bifidobacterium, and Streptococcus have been reported to control blood glucose by regulating satiety signals, maintaining gut barrier integrity, and enhancing the antioxidant activity of pancreatic cells [64,65]. The gut microbiota increases insulin sensitivity through the TGR5 pathway and reduces the expression of proinflammatory cytokines (e.g., tumor necrosis factor α (TNF-α), interleukin-6 (IL-6), and interleukin-1 (IL-1)) through nuclear factor kappa-B (NF-κB), which are associated with insulin resistance and oxidative damage to pancreatic β cells [66,67,68,69,70]. Therefore, the gut microbial dysbiosis observed in patients with T2DM may contribute to decreased insulin sensitivity, reduced insulin production, and impaired glucose tolerance.
Probiotic consumption may improve several metabolic disorders caused by T2DM, including upregulating insulin secretion pathways and reducing systemic inflammation and oxidative stress [53,71,72]. Currently, there is still uncertainty as to whether the gut microbial dysbiosis observed in patients with T2DM is a cause or consequence of glycemic dysregulation. However, in clinical studies, the administration of probiotics containing bacteria from the genera lactobacilli, Bifidobacterium, and Streptococcus can reverse the gut microbial imbalance, ultimately positively improving glucose metabolism and glycemic control [53]. This review summarizes some reports from human randomized controlled trials (RCTs) focusing on the effects of probiotic supplementation on glycemic outcomes in adults with T2DM (Table 1), specifically fasting plasma glucose (FPG), fasting plasma insulin (FPI), hemoglobin A1c (HbA1c), and homeostasis model assessment of insulin resistance (HOMA-IR). In these studies, some strains, including Lab. acidophilus, Lbs. casei, Lbs. rhamnosus, Lab. delbrueckii subsp. bulgaricus, B. breve, B. longum, and S. thermophilus, significantly improved blood glucose metabolism and reduced inflammatory damage in T2DM patients [73]. Lpb. plantarum A7 had no effect on glucose metabolism but changed fatty acid metabolism in T2DM patients [74]. Lab. acidophilus, Lbs. casei, Lab. delbrueckii subsp. lactis, Bifidobacterium, B. longum, and B. infantis increased insulin production in T2DM patients [75]. Lactobacilli, Lactococcus, Bifidobacterium, Propionibacterium, and Acetobacter reduced insulin resistance and systemic inflammation in patients with T2DM [76]. As well as some other probiotics (Lab. acidophilus Bb12, Lab. acidophilus La5, Lab. acidophilus NCFM, B. bifidum, Limosilactobacillus (Lmb.) reuteri ADR-1/3, and Lmb. fermentum), they all have a significant alleviating effect on T2DM [77,78,79,80,81,82]. These anti-diabetic probiotics have great potential to become a new clinical treatment for T2DM.

Table 1.
T2DM management with probiotics: studies of randomized, double-blind, controlled clinical trials using human subjects. (FPG: fasting plasma glucose; FPI: fasting plasma insulin; HbA1c: hemoglobin A1c; HOMA-IR: homeostasis model assessment of insulin resistance; hs-CRP: high-sensitivity C-reactive protein).
4. Effects of Various Probiotics on Diabetes in Rodent Studies
Several studies have demonstrated that probiotics can reduce blood sugar levels to varying degrees. Specifically, Latilactobacillus (Ltb.) sakei OK67 [83], Lbs. rhamnosus CCFM0528 [84], Lbs. paracasei subsp. paracasei NTU 101 [85], Lpb. plantarum NCU116 [86], and Lbs. casei CCFM0412 have been shown to effectively lower blood sugar concentrations in animal models of T2DM [87]. Different probiotics employ different mechanisms for inhibiting the development of diabetes. The treatment models, mechanisms of action, action cycles, and dosages of several probiotics are summarized in Table 2. The main mechanisms by which probiotics control diabetes through regulating intestinal microbiota lie in improving intestinal barrier integrity, reducing oxidative stress, enhancing immune response, increasing SCFA production, and providing liver protection (Figure 1).
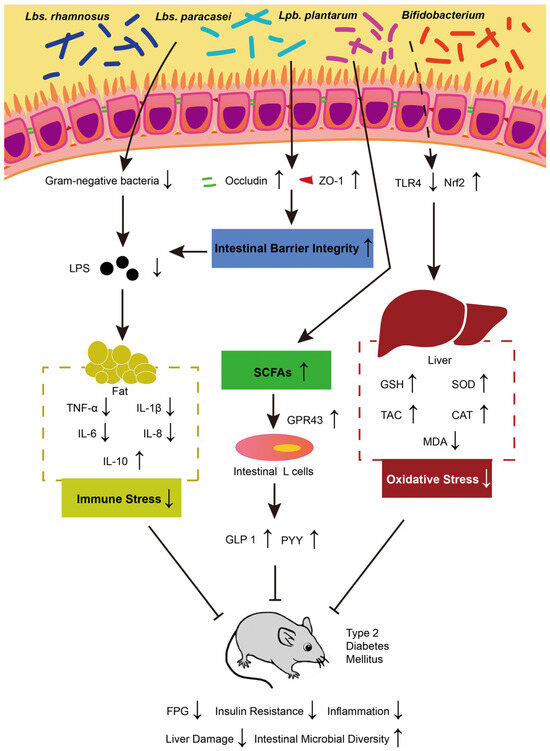
Figure 1.
The possible mechanisms underlying probiotics’ impact on T2DM. TNF-α: tumor necrosis factor α; IL: interleukin; SCFAs: short-chain fatty acids; GLP-1: glucagon-like peptide-1; PYY: peptide YY; GPR43: G-protein coupled receptor 43; TLR 4: Toll-like receptor 4; Nrf 2: nuclear factor erythroid 2-related factor 2; SOD: superoxide dismutase; GSH: glutathione; CAT: catalase; MDA: malondialdehyde; TAC: total antioxidant capacity.
For intestinal barrier integrity, some studies have shown that Lpb. plantarum IS 20506 can improve intestinal permeability and reduce LPS entering the blood by increasing the levels of occludin and ZO-1, thereby strengthening the tight junctions of the intestine [88,89]. Systemic low-grade inflammation arising from immune response in T2DM mice was also reduced. Research indicates that Lbs. rhamnosus CCFM0528 can significantly inhibit the levels of pro-inflammatory factors TNF-α, IL-6, IL-1β, and IL-8 in T2DM mice, while increase the production of the anti-inflammatory factor IL-10 [84]. Similarly, there is also a report showing that Ltb. sakei Probio-65 and Lpb. plantarum Probio-93 reduced the abundance of harmful bacteria and consequently reduced LPS levels in blood [90]. Regarding oxidative stress, B. animalis 01 and Lbs. paracasei NL41 can significantly increase the activities of superoxide dismutase (SOD), glutathione (GSH), and catalase (CAT) and significantly decrease malondialdehyde (MDA) in the liver of T2DM rats, thereby significantly improving antioxidant capacity (TAC) [91,92,93]. This process is regulated by Toll-like receptor 4 (TLR 4) and nuclear factor erythroid 2-related factor 2 (Nrf 2) and plays a protective role against oxidative damage by alleviating redox stress [94]. G-protein coupled receptor 43 (GPR43) can be activated by the increasing level of SCFAs (such as butyrate) produced by compound probiotics (Lpb. plantarum, Lab. delbrueckii subsp. bulgaricus, Lbs. casei, Lab. acidophilus, B. infantis, B. longum, and B. breve). GPR43 modulates intestinal signals of glucagon-like peptide-1 (GLP-1) and peptide YY (PYY) in intestinal L cells, promotes β-cell proliferation, reduces appetite, and thus alleviates glucose tolerance, and enhances energy utilization [91,95,96,97]. Therefore, probiotics alleviate T2DM by regulating the intestinal barrier integrity, oxidative stress, immune response, and SCFA production.
4.1. Lacticaseibacillus rhamnosus
Lbs. rhamnosus GG, first isolated in 1983, is a probiotic known for its potent gastric acid resistance and affinity for intestinal cells. It is now widely used to help control blood sugar in diabetic patients [98]. In rodent studies, the daily administration of Lbs. rhamnosus GG (1 × 108 CFU/mL) to mice for four weeks increased glucose tolerance by reducing endoplasmic reticulum stress [99]. The daily oral administration of 109 CFU/mL of Lbs. rhamnosus HAO 9 to diabetic mice induced by a high-fat diet significantly lowered insulin levels, fasting blood sugar, and proinflammatory cytokines IL-6 and TNF-a [100]. In addition, the administration of Lbs. rhamnosus GG to diabetic mice reduced insulin, glycated hemoglobin, and fasting blood sugar levels and increased serum GLP-1 levels [101]. In diabetic rats, the oral administration of Lbs. rhamnosus BSL and Lbs. rhamnosus R23 reduced insulin resistance by downregulating the expression of glucose-6-phosphatase [102]. Similar results were observed using 3-month-old male zebrafish [103]. In T2DM mice, a blend of three types of probiotics containing Lbs. rhamnosus, Lab. acidophilus, and B. bifidum species (1.8 × 109 CFU) significantly reduced hypothalamic TLR4, IL-6, and NPY and reduced the serine kinases JNK and IKK [104]. These findings highlight the potential value of these probiotics of Lbs. rhamnosus species for diabetes management.
4.2. Lacticaseibacillus paracasei
Endotoxemia, characterized by elevated levels of circulating bacterial lipopolysaccharide, has been identified as a trigger for insulin resistance in mice. Suppressing endotoxemia by probiotic supplementation is considered an effective approach [105]. Treatment with Lbs. paracasei subsp. paracasei NTU101 has been reported to reduce the risk of T2DM by increasing levels of Bifidobacterium animalis subsp. lactis 420 and improving the intestinal environment, which helps maintain intestinal barrier integrity and prevent the transfer of bacterial lipopolysaccharide into the systemic circulation [106]. Similarly, the presence of Lbs. paracasei subsp. paracasei G15 and Lbs. casei Q14 in the intestine has been significantly associated with reduced intestinal mucosal permeability and improved epithelial barrier function. Additionally, Lbs. paracasei subsp. paracasei BCRC12188 has been shown to reduce circulating levels of LPS and inflammatory cytokines, including IL-1β and IL-8, and may alleviate inflammatory states and pancreatic β-cell dysfunction [107]. In SD rats with T2DM, the daily oral administration of 1010 CFU Lbs. paracasei NL41 for 12 weeks reduced insulin resistance, HbA1c, glucagon, leptin, and oxidative stress [93]. Therefore, these strains of Lbs. paracasei genus can alleviate T2DM by regulating intestinal barrier integrity to reduce LPS and inflammatory damage.
4.3. Lactiplantibacillus plantarum
Among lactic acid bacteria, Lpb. plantarum is a facultative heterofermentative member that has been shown to have immunomodulatory and anti-inflammatory effects, as well as promote mucosal barrier integrity [108]. In a high-fat and streptozotocin-induced T2DM rat model, the oral administration of Lpb. plantarum SS18-5 can control body weight, reduce fasting blood glucose and insulin levels, and increase liver glycogen levels [109]. The oral administration of Lpb. plantarum CCFM0236 to diabetic mice not only reduced food intake, blood glucose, glycated hemoglobin, and leptin levels but also regulated serum insulin content and HOMA-IR index [110]. In diabetic mice induced by a high-fat diet, treatment with Lpb. plantarum Probio-093 significantly reduced body weight and improved blood glucose levels [90]. In addition, the oral administration of 4 × 109 CFU of Lpb. plantarum HAC01 daily for 8 weeks reduced FBG, HbA1c, HOMA-IR, and OGTT-AUC in T2DM mice through AMPK and AKT pathways and increased the area of insulin-positive β cells in pancreatic islet tissue [111]. Therefore, these strains of the Lpb. plantarum genus have the potential to alleviate T2DM.
4.4. Bifidobacterium
Bifidobacterium are considered to be the main inhabitants of the intestinal microbiota [112]. Bifidobacterium can metabolize host-derived glycans such as human milk oligosaccharides and mucins [113]. Diabetic patients have lower numbers of Bifidobacterium and Faecalibacterium prausnitzii in their intestines, both of which are Gram-positive bacteria with anti-inflammatory activity [114]. Bifidobacterium has been reported to control the development of diabetes. In Wistar rats with high-fat diet-induced diabetes, the oral administration of B. longum Bb46 (1 × 107 CFU/mL) for 28 days reduced fasting blood glucose, glycated hemoglobin, triglycerides, and total cholesterol [115]. The combination therapy of different Bifidobacterium species (including B. longum, B. bifidum, B. infantis, and B. animalis) improved insulin resistance and reduced blood glucose levels in mice [116]. A probiotic mixture (containing 3 × 1011 CFU/g of B. longum, B. infantis, and B. breve) improved insulin signaling and reduced inflammation in adipose tissue of ApoE-/- rats [117]. The administration of B. animalis subsp. lactis 420 (1 × 109 CFU/mL) to high-fat-diet-induced diabetic rats reduced inflammatory cytokines TNF-α and IL-1β, plasminogen activator inhibitor-1 (PAI-1), and IL-6 in mesenteric adipose tissue while increasing insulin sensitivity [118]. In HFD-fed and STZ-injected T2DM mice, the oral administration of 109 CFU of B. longum DD98 daily for 3 weeks increased butyrate levels in the intestine and decreased pro-inflammatory cytokine levels in the pancreas, thereby improving insulin resistance [119]. Inactivated B. longum BR-108 (3.4 × 1012 cells/g) increased body weight and glucose tolerance while decreasing fat tissue weight, FBG, TC, TG, and nonestesterified fatty acid in Tsumura Suzuki obese diabetes (TSOD) mice [120]. In another study on T2DM rats, the daily oral administration of B. animalis 01 (109 CFU) for 15 weeks reduced body weight, food and water intake, FBG, OGTT-AUC, HbA1c, HOMA-IR, TC, LDL-C, LPS, TNF-α, ALT, AST, and MDA while increasing IL-10, CAT, GSH, GSH-Px, and SOD. This probiotic inhibited the development of T2DM through IRS/PI3K/AKT and Keap1/Nrf2 signaling pathway [92]. In summary, these strains of the Bifidobacterium genus significantly improved the development of T2DM.
Table 2.
T2DM management with probiotics: in vivo studies in rodents (FPG: fasting plasma glucose; FPI: fasting plasma insulin; HbA1c: hemoglobin A1c; HOMA-IR: homeostasis model assessment of insulin resistance); TC: total cholesterol; TG: total triglycerides; OGTT-AUC: oral glucose tolerance test-area under the curve; LDL-C: low-density lipoprotein-cholesterol; ALT: alanine transaminase; AST: aspartate transaminase; GSH-Px: glutathione peroxidase; SOD: superoxide dismutase; GSH: glutathione; CAT: catalase; MDA: malondialdehyde; IL: interleukin; TLR4: Toll-like receptor 4; NPY: neuropeptide Y; SCFAs: short-chain fatty acids.
Table 2.
T2DM management with probiotics: in vivo studies in rodents (FPG: fasting plasma glucose; FPI: fasting plasma insulin; HbA1c: hemoglobin A1c; HOMA-IR: homeostasis model assessment of insulin resistance); TC: total cholesterol; TG: total triglycerides; OGTT-AUC: oral glucose tolerance test-area under the curve; LDL-C: low-density lipoprotein-cholesterol; ALT: alanine transaminase; AST: aspartate transaminase; GSH-Px: glutathione peroxidase; SOD: superoxide dismutase; GSH: glutathione; CAT: catalase; MDA: malondialdehyde; IL: interleukin; TLR4: Toll-like receptor 4; NPY: neuropeptide Y; SCFAs: short-chain fatty acids.
| Probiotics | Subjects | Model Type | Effect | Mechanism | Treatment | Ref. |
|---|---|---|---|---|---|---|
| Lpb. plantarum, Lab. delbrueckii subsp. bulgaricus bulgaricus, Lbs. casei, Lab. acidophilus, B. infantis, B. longum, B. breve | 40 Wistar rats | HFD + STZ | In probiotic group, FPG and insulin resistance decreased, and total antioxidant capacity increased. | Control of T2DM by increasing GLP-1 levels and reducing oxidative stress. | 5 × 1010 CFU/mL in water, 4 weeks | [91] |
| Lbs. paracasei subsp. paracasei NL41 | 18 Sprague Dawley (SD) rats | HFD + STZ | In probiotic group, insulin resistance, HbA1c, glucagon, leptin, and oxidative stress decreased. | N/A. | 1010 CFU, oral administration, once per day for 12 weeks | [93] |
| B. longum DD98 and selenium-enriched B. longum DD98 | 48 C57BL/6J mice | HFD + STZ | In probiotic group, FBG, HbA1c, and insulin resistance decreased. | Probiotics increase butyrate levels in the intestine and decrease pro-inflammatory cytokine levels in the pancreas, thereby improving insulin resistance. | 1 × 109 CFU, oral administration, once per day for 3 weeks | [119] |
| Inactivated B. longum BR-108 (IBL) | 25 Tsumura Suzuki obese diabetes (TSOD) mice | Spontaneous obesity | In probiotic group, body weight and glucose tolerance increased, fat tissue weight, FBG, TC, TG, and nonestesterified fatty acid decreased. | Probiotics absorb cholesterol and produce short-chain fatty acids inhibiting cholesterol synthesis in the liver. | IBL (3.4 × 1012 cells/g), 50, 100, and 150 mg/kg BW, for 30 days | [120] |
| B. animalis 01 | 24 Sprague Dawley (SD) rats | HFD + STZ | In probiotic group, body weight, food and water intake, FBG, OGTT-AUC, HbA1c, HOMA-IR, TC, LDL-C, LPS, TNF-α, ALT, AST, and MDA decreased, and IL-10, CAT, GSH, GSH-Px, and SOD increased. | Activation of IRS/PI3K/AKT and Keap1/Nrf2 signaling. | 109 CFU, oral administration, once per day for 15 weeks | [92] |
| Lbs. rhamnosus, Lab. acidophilus and B. bifidum | 24 Swiss mice | DIO (diet-induced obesity) | In probiotic group, FBG, food intake, intestinal permeability, LPS translocation, and systemic low-grade inflammation decreased. | Probiotics significantly reduce hypothalamic TLR4, IL-6, NPY, and reduce the serine kinases JNK and IKK. | 1.8 × 109 CFU, once per day for 5 weeks | [104] |
| Lpb. plantarum HAC01 | 50 C57BL/6J mice | HFD + STZ | In probiotic group, FBG, HbA1c, HOMA-IR, and OGTT-AUC decreased, and islet insulin-positive β cell area, and butyric acid increased. | Activating AMPK and Akt pathways in the liver. | 4 × 109 CFU, once per day for 10 weeks | [111] |
| Lpb. plantarum Probio-093 | 40 C57BL/6J mice | HFD | In probiotic group, α-glucosidase, α-amylase activity, body weight, FPG, and intestinal inflammation decreased, and SCFAs increased. | Probiotics reduce the abundance of Deferribacteria and Proteobacteria, increases the abundance of Actinobacteria and Bacteroidetes, regulates the intestinal barrier, and enhances immune response. | 108 CFU, once per day for 8 weeks | [90] |
5. Conclusions
This review discussed the correlation between probiotics, intestinal flora, and diabetes and the potential ability of probiotics to alleviate diabetes. Firstly, probiotics reduce the abundance of Gram-negative bacteria by regulating intestinal flora, thereby reducing LPS levels and leading to reduced immune stress; secondly, probiotics strengthen the tight junctions of the intestinal epithelial barrier, which also leads to reduced LPS levels; third, probiotics produce more beneficial short-chain fatty acids (such as butyrate, acetate, and propionate), thereby enhancing intestinal metabolism; finally, probiotics protect the liver by reducing oxidative stress. These are the main mechanisms by which probiotics alleviate diabetes. Some probiotics from Lbs. rhamnosus, Lbs. paracasei, Lpb. plantarum, and Bifidobacterium species have many experimental reports proving their anti-diabetic effects. Currently, anti-diabetic probiotic supplements are still in the clinical trial stage, and there are no mature commercial products as a treatment method. The present study may contribute to the development of probiotic supplements with anti-diabetic effects. However, the underlying mechanism of how the imbalance of intestinal microbes affects diabetes or vice versa awaits further investigations. In addition, despite the beneficial effects of probiotics on metabolic diseases, including diabetes, the side effects and health risks due to long-term intake of probiotics have not yet been fully validated.
Author Contributions
S.L.: conceptualization; investigation; writing—original draft; writing—review and editing; visualization. Z.L., Q.Z. (Qi Zhang), D.S., Y.L. and P.W.: writing—review and editing; W.S. and Q.Z. (Qian Zhang): conceptualization; funding acquisition; project administration; writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Beijing Nova Program 20220484144, the National Natural Science Foundation of China Grant 82272566, the Beijing Natural Science Foundation No. 7244368, the 111 Project from the Education Ministry of China B18053, and the Chinese Universities Scientific Fund 2024RC013.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhou, C.; Byard, R.W. An analysis of the morbidity and mortality of diabetes mellitus in a forensic context. J. Forensic Sci. 2018, 63, 1149–1154. [Google Scholar] [CrossRef] [PubMed]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef]
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850. [Google Scholar] [CrossRef] [PubMed]
- Li, B.-Y.; Xu, X.-Y.; Gan, R.-Y.; Sun, Q.-C.; Meng, J.-M.; Shang, A.; Mao, Q.-Q.; Li, H.-B. Targeting gut microbiota for the prevention and management of diabetes mellitus by dietary natural products. Foods 2019, 8, 440. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Mazurakova, A.; Kubatka, P.; Büsselberg, D. Flavonoids’ Dual Benefits in Gastrointestinal Cancer and Diabetes: A Potential Treatment on the Horizon? Cancers 2022, 14, 6073. [Google Scholar] [CrossRef]
- Arnold, S.V.; Bhatt, D.L.; Barsness, G.W.; Beatty, A.L.; Deedwania, P.C.; Inzucchi, S.E.; Kosiborod, M.; Leiter, L.A.; Lipska, K.J.; Newman, J.D. Clinical management of stable coronary artery disease in patients with type 2 diabetes mellitus: A scientific statement from the American Heart Association. Circulation 2020, 141, e779–e806. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Wei, X.; Lu, Z.; Li, L.; Hu, Y.; Sun, F.; Jiang, Y.; Ma, H.; Zheng, H.; Yang, G. Activation of TRPV1 channel antagonizes diabetic nephropathy through inhibiting endoplasmic reticulum-mitochondria contact in podocytes. Metabolism 2020, 105, 154182. [Google Scholar] [CrossRef] [PubMed]
- Banik, P.C.; Barua, L.; Moniruzzaman, M.; Mondal, R.; Zaman, F.; Ali, L. Risk of diabetic foot ulcer and its associated factors among Bangladeshi subjects: A multicentric cross-sectional study. BMJ Open 2020, 10, e034058. [Google Scholar] [CrossRef]
- Tan, T.-E.; Wong, T.Y. Diabetic retinopathy: Looking forward to 2030. Front. Endocrinol. 2023, 13, 1077669. [Google Scholar] [CrossRef]
- Erener, S. Diabetes, infection risk and COVID-19. Mol. Metab. 2020, 39, 101044. [Google Scholar] [CrossRef]
- Selvin, E.; Juraschek, S.P. Diabetes epidemiology in the COVID-19 pandemic. Diabetes Care 2020, 43, 1690–1694. [Google Scholar] [CrossRef] [PubMed]
- Kgosidialwa, O.; Egan, A.M.; Carmody, L.; Kirwan, B.; Gunning, P.; Dunne, F.P. Treatment with diet and exercise for women with gestational diabetes mellitus diagnosed using IADPSG criteria. J. Clin. Endocrinol. Metab. 2015, 100, 4629–4636. [Google Scholar] [CrossRef] [PubMed]
- Khalili, L.; Alipour, B.; Jafar-Abadi, M.A.; Faraji, I.; Hassanalilou, T.; Mesgari Abbasi, M.; Vaghef-Mehrabany, E.; Sani, M.A. The Effects of Lactobacillus casei on glycemic response, serum Sirtuin1 and Fetuin-A levels in patients with type 2 diabetes Mellitus: A Randomized Controlled Trial. Iran Biomed J. 2019, 23, 68–77. [Google Scholar] [CrossRef]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms linking obesity to insulin resistance and type 2 diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef] [PubMed]
- Blaslov, K.; Naranđa, F.S.; Kruljac, I.; Renar, I.P. Treatment approach to type 2 diabetes: Past, present and future. World J. Diabetes 2018, 9, 209. [Google Scholar] [CrossRef] [PubMed]
- Walicka, M.; Krysiński, A.; La Rosa, G.R.M.; Sun, A.; Campagna, D.; Di Ciaula, A.; Dugal, T.; Kengne, A.; Le Dinh, P.; Misra, A. Influence of quitting smoking on diabetes-related complications: A scoping review with a systematic search strategy. Diabetes Metab. Syndr. Clin. Res. Rev. 2024, 18, 103044. [Google Scholar] [CrossRef] [PubMed]
- Leong, C.; Bolton, J.M.; Ekuma, O.; Prior, H.J.; Singal, D.; Nepon, J.; Konrad, G.; Paille, M.; Finlayson, G.; Nickel, N. Association of alcohol use disorder on alcohol-related cancers, diabetes, ischemic heart disease and death: A population-based, matched cohort study. Addiction 2022, 117, 368–381. [Google Scholar] [CrossRef]
- Qi, Q.; Li, J.; Yu, B.; Moon, J.-Y.; Chai, J.C.; Merino, J.; Hu, J.; Ruiz-Canela, M.; Rebholz, C.; Wang, Z. Host and gut microbial tryptophan metabolism and type 2 diabetes: An integrative analysis of host genetics, diet, gut microbiome and circulating metabolites in cohort studies. Gut 2022, 71, 1095–1105. [Google Scholar] [CrossRef] [PubMed]
- Lynch, S.V.; Pedersen, O. The human intestinal microbiome in health and disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef]
- Diao, H.; Yan, H.; Xiao, Y.; Yu, B.; Yu, J.; He, J.; Zheng, P.; Zeng, B.; Wei, H.; Mao, X. Intestinal microbiota could transfer host Gut characteristics from pigs to mice. BMC Microbiol. 2016, 16, 238. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Yun, C.; Pang, Y.; Qiao, J. The impact of the gut microbiota on the reproductive and metabolic endocrine system. Gut Microbes 2021, 13, 1894070. [Google Scholar] [CrossRef] [PubMed]
- Jameson, K.G.; Olson, C.A.; Kazmi, S.A.; Hsiao, E.Y. Toward understanding microbiome-neuronal signaling. Mol. Cell 2020, 78, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhang, J.; Wang, R. Gut microbiota modulates drug pharmacokinetics. Drug Metab. Rev. 2018, 50, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Zhao, T.; Zhang, H.; Yin, X.; Zhao, H.; Ma, L.; Yan, M.; Peng, L.; Wang, Q.; Dong, X.; Li, P. Tangshen formula modulates gut microbiota and reduces gut-derived toxins in diabetic nephropathy rats. Biomed. Pharmacother. 2020, 129, 110325. [Google Scholar] [CrossRef] [PubMed]
- Jensen, A.B.; Sørensen, T.I.; Pedersen, O.; Jess, T.; Brunak, S.; Allin, K.H. Increase in clinically recorded type 2 diabetes after colectomy. Elife 2018, 7, e37420. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Levy, M.; Grosheva, I.; Zheng, D.; Soffer, E.; Blacher, E.; Braverman, S.; Tengeler, A.C.; Barak, O.; Elazar, M. Hyperglycemia drives intestinal barrier dysfunction and risk for enteric infection. Science 2018, 359, 1376–1383. [Google Scholar] [CrossRef]
- Bezirtzoglou, E.; Stavropoulou, E. Immunology and probiotic impact of the newborn and young children intestinal microflora. Anaerobe 2011, 17, 369–374. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Todorov, S.D.; Baretto Penna, A.L.; Venema, K.; Holzapfel, W.H.; Chikindas, M.L. Recommendations for the use of standardised abbreviations for the former Lactobacillus genera, reclassified in the year 2020. Benef. Microbes 2023, 15, 1–4. [Google Scholar] [CrossRef]
- Gomes, A.C.; Bueno, A.A.; de Souza, R.G.M.; Mota, J.F. Gut microbiota, probiotics and diabetes. Nutr. J. 2014, 13, 60. [Google Scholar] [CrossRef] [PubMed]
- Stavropoulou, E.; Bezirtzoglou, E. Probiotics in medicine: A long debate. Front. Immunol. 2020, 11, 554558. [Google Scholar] [CrossRef] [PubMed]
- Vassos, D.; Bezirtzoglou, E.; Voidarou, C.; Alexopoulos, A.; Maipa, V. Biochemical and antimicrobial profile of Enterococcus faecium and E. faecalis isolated from traditional dairy products and infant intestinal microbiota. Microb. Ecol. Health Dis. 2009, 21, 241–250. [Google Scholar]
- Krishna Rao, R.; Samak, G. Protection and restitution of gut barrier by probiotics: Nutritional and clinical implications. Curr. Nutr. Food Sci. 2013, 9, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Dimidi, E.; Christodoulides, S.; Scott, S.M.; Whelan, K. Mechanisms of action of probiotics and the gastrointestinal microbiota on gut motility and constipation. Adv. Nutr. 2017, 8, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, A.; Dash, J.; Kancharla, S.; Kolli, P.; Mahajan, D.; Senapati, S.; Jena, M.K. Probiotics: A promising candidate for management of colorectal cancer. Cancers 2021, 13, 3178. [Google Scholar] [CrossRef] [PubMed]
- Filidou, E.; Kolios, G. Probiotics in intestinal mucosal healing: A new therapy or an old friend? Pharmaceuticals 2021, 14, 1181. [Google Scholar] [CrossRef] [PubMed]
- Gou, H.-Z.; Zhang, Y.-L.; Ren, L.-F.; Li, Z.-J.; Zhang, L. How do intestinal probiotics restore the intestinal barrier? Front. Microbiol. 2022, 13, 929346. [Google Scholar] [CrossRef] [PubMed]
- Pothuraju, R.; Chaudhary, S.; Rachagani, S.; Kaur, S.; Roy, H.K.; Bouvet, M.; Batra, S.K. Mucins, gut microbiota, and postbiotics role in colorectal cancer. Gut Microbes 2021, 13, 1974795. [Google Scholar] [CrossRef]
- Javanshir, N.; Hosseini, G.N.G.; Sadeghi, M.; Esmaeili, R.; Satarikia, F.; Ahmadian, G.; Allahyari, N. Evaluation of the Function of Probiotics, Emphasizing the Role of their Binding to the Intestinal Epithelium in the Stability and their Effects on the Immune System. Biol. Proced. Online 2021, 23, 23. [Google Scholar] [CrossRef]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EBioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed]
- Candela, M.; Biagi, E.; Soverini, M.; Consolandi, C.; Quercia, S.; Severgnini, M.; Peano, C.; Turroni, S.; Rampelli, S.; Pozzilli, P. Modulation of gut microbiota dysbioses in type 2 diabetic patients by macrobiotic Ma-Pi 2 diet. Br. J. Nutr. 2016, 116, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Adachi, K.; Sugiyama, T.; Shimozato, A.; Ebi, M.; Ogasawara, N.; Funaki, Y.; Goto, C.; Sasaki, M.; Kasugai, K. Association of intestinal microbiota with metabolic markers and dietary habits in patients with type 2 diabetes. Digestion 2016, 94, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, F.H.; Tremaroli, V.; Nookaew, I.; Bergström, G.; Behre, C.J.; Fagerberg, B.; Nielsen, J.; Bäckhed, F. Gut metagenome in European women with normal, impaired and diabetic glucose control. Nature 2013, 498, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ma, C.; Han, L.; Nawaz, M.; Gao, F.; Zhang, X.; Yu, P.; Zhao, C.a.; Li, L.; Zhou, A. Molecular characterisation of the faecal microbiota in patients with type II diabetes. Curr. Microbiol. 2010, 61, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, D.; Fang, Z.; Jie, Z.; Qiu, X.; Zhang, C.; Chen, Y.; Ji, L. Human gut microbiota changes reveal the progression of glucose intolerance. PLoS ONE 2013, 8, e71108. [Google Scholar] [CrossRef]
- Salamon, D.; Sroka-Oleksiak, A.; Kapusta, P.; Szopa, M.; Mrozińska, S.; Ludwig-Słomczyńska, A.H.; Wołkow, P.P.; Bulanda, M.; Klupa, T.; Małecki, M.T. Characteristics of gut microbiota in adult patients with type 1 and type 2 diabetes based on next-generation sequencing of the 16S rRNA gene fragment. Pol. Arch. Intern. Med. 2018, 128, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Larsen, N.; Vogensen, F.K.; Van Den Berg, F.W.; Nielsen, D.S.; Andreasen, A.S.; Pedersen, B.K.; Al-Soud, W.A.; Sørensen, S.J.; Hansen, L.H.; Jakobsen, M. Gut microbiota in human adults with type 2 diabetes differs from non-diabetic adults. PLoS ONE 2010, 5, e9085. [Google Scholar] [CrossRef]
- Murphy, R.; Tsai, P.; Jüllig, M.; Liu, A.; Plank, L.; Booth, M. Differential changes in gut microbiota after gastric bypass and sleeve gastrectomy bariatric surgery vary according to diabetes remission. Obes. Surg. 2017, 27, 917–925. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, D.; Murphy, S.; Boileau, T.; Park, J.; O’Brien, F.; Groeger, D.; Konieczna, P.; Ziegler, M.; Scully, P.; Shanahan, F. Bifidobacterium animalis AHC7 protects against pathogen-induced NF-κB activation in vivo. BMC Immunol. 2010, 11, 63. [Google Scholar]
- Gao, R.; Zhu, C.; Li, H.; Yin, M.; Pan, C.; Huang, L.; Kong, C.; Wang, X.; Zhang, Y.; Qu, S. Dysbiosis signatures of gut microbiota along the sequence from healthy, young patients to those with overweight and obesity. Obesity 2018, 26, 351–361. [Google Scholar] [CrossRef]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [PubMed]
- Kasińska, M.A.; Drzewoski, J. Effectiveness of probiotics in type 2 diabetes: A meta-analysis. Pol. Arch. Med. Wewn. 2015, 125, 803–813. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Reichardt, N.; Duncan, S.H.; Young, P.; Belenguer, A.; McWilliam Leitch, C.; Scott, K.P.; Flint, H.J.; Louis, P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014, 8, 1323–1335. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Tolhurst, G.; Heffron, H.; Lam, Y.S.; Parker, H.E.; Habib, A.M.; Diakogiannaki, E.; Cameron, J.; Grosse, J.; Reimann, F.; Gribble, F.M. Short-chain fatty acids stimulate glucagon-like peptide-1 secretion via the G-protein–coupled receptor FFAR2. Diabetes 2012, 61, 364–371. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Kovatcheva-Datchary, P.; Goncalves, D.; Vinera, J.; Zitoun, C.; Duchampt, A.; Bäckhed, F.; Mithieux, G. Microbiota-generated metabolites promote metabolic benefits via gut-brain neural circuits. Cell 2014, 156, 84–96. [Google Scholar] [CrossRef]
- Nguyen, T.L.A.; Vieira-Silva, S.; Liston, A.; Raes, J. How informative is the mouse for human gut microbiota research? Dis. Model. Mech. 2015, 8, 1–16. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, H.; Wang, X.; Yu, X.; Hu, C.; Zhang, X. The family Coriobacteriaceae is a potential contributor to the beneficial effects of Roux-en-Y gastric bypass on type 2 diabetes. Surg. Obes. Relat. Dis. 2018, 14, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, H.; Zou, H.; Chen, Z.; Jiang, T.; Huang, M. Correlation of Differentially Expressed lncRNAs with Intestinal Flora Imbalance, Small Intestinal Permeability, and Glucose Uptake in T2DM Mice. Appl. Biochem. Biotechnol. 2024, 1–19. [Google Scholar] [CrossRef]
- Chen, X.; Chen, C.; Fu, X. Dendrobium officinale polysaccharide alleviates type 2 diabetes mellitus by restoring gut microbiota and repairing intestinal barrier via the LPS/TLR4/TRIF/NF-kB Axis. J. Agric. Food Chem. 2023, 71, 11929–11940. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, M.; Basoulis, D.; Konstantopoulou, S.; Dimitriadi, D.; Gyftopoulou, K.; Skarmoutsou, N.; Fakiri, E.M. Health benefits of probiotics: A review. Int. Sch. Res. Not. 2013, 2013, 481651. [Google Scholar] [CrossRef] [PubMed]
- Yadav, H.; Jain, S.; Sinha, P. Antidiabetic effect of probiotic dahi containing Lactobacillus acidophilus and Lactobacillus casei in high fructose fed rats. Nutrition 2007, 23, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Prawitt, J.; Caron, S.; Staels, B. Bile acid metabolism and the pathogenesis of type 2 diabetes. Curr. Diab. Rep. 2011, 11, 160–166. [Google Scholar] [CrossRef]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef]
- Shyangdan, D.S.; Royle, P.; Clar, C.; Sharma, P.; Waugh, N.; Snaith, A. Glucagon-like peptide analogues for type 2 diabetes mellitus. Cochrane Database Syst. Rev. 2011, 2011, CD006423. [Google Scholar] [CrossRef]
- Borst, S.E. The role of TNF-α in insulin resistance. Endocrine 2004, 23, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Sun, Z. Antiaging gene klotho attenuates pancreatic β-cell apoptosis in type 1 diabetes. Diabetes 2015, 64, 4298–4311. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, H.; Lu, C.; Kang, Z.; Luo, Y.; Feng, L.; Lu, X. Fermented milk supplemented with probiotics and prebiotics can effectively alter the intestinal microbiota and immunity of host animals. J. Dairy Sci. 2012, 95, 4813–4822. [Google Scholar] [CrossRef] [PubMed]
- Tonucci, L.B.; Dos Santos, K.M.O.; de Oliveira, L.L.; Ribeiro, S.M.R.; Martino, H.S.D. Clinical application of probiotics in type 2 diabetes mellitus: A randomized, double-blind, placebo-controlled study. Clin. Nutr. 2017, 36, 85–92. [Google Scholar] [CrossRef]
- Asemi, Z.; Zare, Z.; Shakeri, H.; Sabihi, S.-s.; Esmaillzadeh, A. Effect of multispecies probiotic supplements on metabolic profiles, hs-CRP, and oxidative stress in patients with type 2 diabetes. Ann. Nutr. Metab. 2013, 63, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feizollahzadeh, S.; Ghiasvand, R.; Rezaei, A.; Khanahmad, H.; Sadeghi, A.; Hariri, M. Effect of probiotic soy milk on serum levels of adiponectin, inflammatory mediators, lipid profile, and fasting blood glucose among patients with type II diabetes mellitus. Probiotics Antimicrob. Proteins 2017, 9, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Firouzi, S.; Majid, H.A.; Ismail, A.; Kamaruddin, N.A.; Barakatun-Nisak, M.-Y. Effect of multi-strain probiotics (multi-strain microbial cell preparation) on glycemic control and other diabetes-related outcomes in people with type 2 diabetes: A randomized controlled trial. Eur. J. Nutr. 2017, 56, 1535–1550. [Google Scholar] [CrossRef] [PubMed]
- Kobyliak, N.; Falalyeyeva, T.; Mykhalchyshyn, G.; Kyriienko, D.; Komissarenko, I. Effect of alive probiotic on insulin resistance in type 2 diabetes patients: Randomized clinical trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2018, 12, 617–624. [Google Scholar] [CrossRef]
- Mohamadshahi, M.; Veissi, M.; Haidari, F.; Shahbazian, H.; Kaydani, G.-A.; Mohammadi, F. Effects of probiotic yogurt consumption on inflammatory biomarkers in patients with type 2 diabetes. BioImpacts BI 2014, 4, 83. [Google Scholar]
- Ejtahed, H.S.; Mohtadi-Nia, J.; Homayouni-Rad, A.; Niafar, M.; Asghari-Jafarabadi, M.; Mofid, V. Probiotic yogurt improves antioxidant status in type 2 diabetic patients. Nutrition 2012, 28, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Andreasen, A.S.; Larsen, N.; Pedersen-Skovsgaard, T.; Berg, R.M.; Møller, K.; Svendsen, K.D.; Jakobsen, M.; Pedersen, B.K. Effects of Lactobacillus acidophilus NCFM on insulin sensitivity and the systemic inflammatory response in human subjects. Br. J. Nutr. 2010, 104, 1831–1838. [Google Scholar] [CrossRef]
- Moroti, C.; Souza Magri, L.F.; de Rezende Costa, M.; Cavallini, D.C.; Sivieri, K. Effect of the consumption of a new symbiotic shake on glycemia and cholesterol levels in elderly people with type 2 diabetes mellitus. Lipids Health Dis. 2012, 11, 29. [Google Scholar] [CrossRef]
- Hsieh, M.-C.; Tsai, W.-H.; Jheng, Y.-P.; Su, S.-L.; Wang, S.-Y.; Lin, C.-C.; Chen, Y.-H.; Chang, W.-W. The beneficial effects of Lactobacillus reuteri ADR-1 or ADR-3 consumption on type 2 diabetes mellitus: A randomized, double-blinded, placebo-controlled trial. Sci. Rep. 2018, 8, 16791. [Google Scholar] [CrossRef] [PubMed]
- Raygan, F.; Ostadmohammadi, V.; Bahmani, F.; Asemi, Z. The effects of vitamin D and probiotic co-supplementation on mental health parameters and metabolic status in type 2 diabetic patients with coronary heart disease: A randomized, double-blind, placebo-controlled trial. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 84, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.-M.; Jeong, J.-J.; Woo, K.H.; Han, M.J.; Kim, D.-H. Lactobacillus sakei OK67 ameliorates high-fat diet–induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr. Res. 2016, 36, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Oral administration of Lactobacillus rhamnosus CCFM0528 improves glucose tolerance and cytokine secretion in high-fat-fed, streptozotocin-induced type 2 diabetic mice. J. Funct. Foods 2014, 10, 318–326. [Google Scholar] [CrossRef]
- Hung, S.-C.; Tseng, W.-T.; Pan, T.-M. Lactobacillus paracasei subsp. paracasei NTU 101 ameliorates impaired glucose tolerance induced by a high-fat, high-fructose diet in Sprague-Dawley rats. J. Funct. Foods 2016, 24, 472–481. [Google Scholar] [CrossRef]
- Li, C.; Ding, Q.; Nie, S.-P.; Zhang, Y.-S.; Xiong, T.; Xie, M.-Y. Carrot Juice Fermented with Lactobacillus plantarum NCU116 Ameliorates Type 2 Diabetes in Rats. J. Agric. Food Chem. 2014, 62, 11884–11891. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Zhang, Q.; Dang, H.; Liu, X.; Tian, F.; Zhao, J.; Chen, Y.; Zhang, H.; Chen, W. Antidiabetic effect of Lactobacillus casei CCFM0412 on mice with type 2 diabetes induced by a high-fat diet and streptozotocin. Nutrition 2014, 30, 1061–1068. [Google Scholar] [CrossRef] [PubMed]
- Ranuh, R.; Subijanto, M.; Surono, I. The Role of probiotic lactobacillus plantarum IS 20506 on Occludin and ZO-1 of intestinal tight junctions rehabilitation. Adv. Nat. Appl. Sci. 2013, 7, 480–484. [Google Scholar]
- Rose, E.C.; Odle, J.; Blikslager, A.T.; Ziegler, A.L. Probiotics, prebiotics and epithelial tight junctions: A promising approach to modulate intestinal barrier function. Int. J. Mol. Sci. 2021, 22, 6729. [Google Scholar] [CrossRef]
- Gulnaz, A.; Nadeem, J.; Han, J.-H.; Lew, L.-C.; Son, J.-D.; Park, Y.-H.; Rather, I.A.; Hor, Y.-Y. Lactobacillus SPS in reducing the risk of diabetes in high-fat diet-induced diabetic mice by modulating the gut microbiome and inhibiting key digestive enzymes associated with diabetes. Biology 2021, 10, 348. [Google Scholar] [CrossRef]
- Pegah, A.; Abbasi-Oshaghi, E.; Khodadadi, I.; Mirzaei, F.; Tayebinia, H. Probiotic and resveratrol normalize GLP-1 levels and oxidative stress in the intestine of diabetic rats. Metab. Open 2021, 10, 100093. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, S.; Zeng, Z.; Qin, Y.; Shen, Q.; Li, P. Anti-diabetic effects of Bifidobacterium animalis 01 through improving hepatic insulin sensitivity in type 2 diabetic rat model. J. Funct. Foods 2020, 67, 103843. [Google Scholar] [CrossRef]
- Zeng, Z.; Yuan, Q.; Yu, R.; Zhang, J.; Ma, H.; Chen, S. Ameliorative effects of probiotic Lactobacillus paracasei NL41 on insulin sensitivity, oxidative stress, and beta-cell function in a type 2 diabetes mellitus rat model. Mol. Nutr. Food Res. 2019, 63, 1900457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Xu, Q.; Gao, Y.; Cao, H.; Lian, Y.; Li, Z.; Xu, J.; Zhong, M.; Li, J.; Wei, R. Polysaccharides from Dicliptera chinensis ameliorate liver disturbance by regulating TLR-4/NF-κB and AMPK/Nrf2 signalling pathways. J. Cell. Mol. Med. 2020, 24, 6397–6409. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ren, J.; Huang, L.; Pang, B.; Liu, X.; Liu, X.; Li, B.; Shan, Y. Antidiabetic effects of Lactobacillus casei fermented yogurt through reshaping gut microbiota structure in type 2 diabetic rats. J. Agric. Food Chem. 2018, 66, 12696–12705. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Sailike, J.; Sun, X.; Abuduwaili, N.; Tuoliuhan, H.; Yusufu, M.; Nabi, X.-h. Fourteen composite probiotics alleviate type 2 diabetes through modulating gut microbiota and modifying M1/M2 phenotype macrophage in db/db mice. Pharmacol. Res. 2020, 161, 105150. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Inoue, D.; Hirano, K.; Tsujimoto, G. The SCFA receptor GPR43 and energy metabolism. Front. Endocrinol. 2014, 5, 93736. [Google Scholar] [CrossRef] [PubMed]
- Capurso, L. Thirty years of Lactobacillus rhamnosus GG: A review. J. Clin. Gastroenterol. 2019, 53, S1–S41. [Google Scholar] [CrossRef] [PubMed]
- Park, K.-Y.; Kim, B.; Hyun, C.-K. Lactobacillus rhamnosus GG improves glucose tolerance through alleviating ER stress and suppressing macrophage activation in db/db mice. J. Clin. Biochem. Nutr. 2015, 56, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Liao, W.; Dong, Y.; Bai, C.; Gai, Z. Lacticaseibacillus rhamnosus Hao9 exerts antidiabetic effects by regulating gut microbiome, glucagon metabolism, and insulin levels in type 2 diabetic mice. Front. Nutr. 2023, 9, 1081778. [Google Scholar] [CrossRef]
- Yan, F.; Li, N.; Yue, Y.; Wang, C.; Zhao, L.; Evivie, S.E.; Li, B.; Huo, G. Screening for potential novel probiotics with dipeptidyl peptidase IV-inhibiting activity for type 2 diabetes attenuation in vitro and in vivo. Front. Microbiol. 2020, 10, 2855. [Google Scholar] [CrossRef] [PubMed]
- Farida, E.; Nuraida, L.; Giriwono, P.E.; Jenie, B.S. Lactobacillus rhamnosus reduces blood glucose level through downregulation of gluconeogenesis gene expression in streptozotocin-induced diabetic rats. Int. J. Food Sci. 2020, 2020, 6108575. [Google Scholar] [CrossRef]
- Bootorabi, F.; Saadat, F.; Falak, R.; Manouchehri, H.; Changizi, R.; Mohammadi, H.; Safavifar, F.; Khorramizadeh, M.R. Gut micobiota alteration by Lactobacillus rhamnosus reduces pro-inflammatory cytokines and glucose level in the adult model of Zebrafish. BMC Res. Notes 2021, 14, 302. [Google Scholar] [CrossRef]
- Bagarolli, R.A.; Tobar, N.; Oliveira, A.G.; Araújo, T.G.; Carvalho, B.M.; Rocha, G.Z.; Vecina, J.F.; Calisto, K.; Guadagnini, D.; Prada, P.O. Probiotics modulate gut microbiota and improve insulin sensitivity in DIO mice. J. Nutr. Biochem. 2017, 50, 16–25. [Google Scholar] [CrossRef]
- Sigalet, D.L.; Wallace, L.E.; Holst, J.J.; Martin, G.R.; Kaji, T.; Tanaka, H.; Sharkey, K.A. Enteric neural pathways mediate the anti-inflammatory actions of glucagon-like peptide 2. Am. J. Physiol.-Gastrointest. Liver Physiol. 2007, 293, G211–G221. [Google Scholar] [CrossRef]
- Stenman, L.K.; Waget, A.; Garret, C.; Briand, F.; Burcelin, R.; Sulpice, T.; Lahtinen, S. Probiotic B420 and prebiotic polydextrose improve efficacy of antidiabetic drugs in mice. Diabetol. Metab. Syndr. 2015, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.-H.; Chen, Y.-P.; Chen, M.-J. Selecting probiotics with the abilities of enhancing GLP-1 to mitigate the progression of type 1 diabetes in vitro and in vivo. J. Funct. Foods 2015, 18, 473–486. [Google Scholar] [CrossRef]
- Chong, H.-X.; Yusoff, N.A.A.; Hor, Y.-Y.; Lew, L.-C.; Jaafar, M.H.; Choi, S.-B.; Yusoff, M.S.; Wahid, N.; Abdullah, M.F.I.; Zakaria, N. Lactobacillus plantarum DR7 improved upper respiratory tract infections via enhancing immune and inflammatory parameters: A randomized, double-blind, placebo-controlled study. J. Dairy Sci. 2019, 102, 4783–4797. [Google Scholar] [CrossRef]
- Yang, F.; Wang, J.; Zhang, H.; Xie, Y.; Jin, J.; Liu, H.; Pang, X.; Hao, H. Hypoglycemic effects of space-induced Lactobacillus plantarum SS18-5 on type 2 diabetes in a rat model. J. Food Biochem. 2021, 45, e13899. [Google Scholar] [CrossRef]
- Li, X.; Wang, N.; Yin, B.; Fang, D.; Jiang, T.; Fang, S.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Effects of Lactobacillus plantarum CCFM0236 on hyperglycaemia and insulin resistance in high-fat and streptozotocin-induced type 2 diabetic mice. J. Appl. Microbiol. 2016, 121, 1727–1736. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, D.; Park, G.-S.; Ko, S.-H.; Park, J.; Lee, Y.-K.; Kang, J. Lactobacillus plantarum HAC01 ameliorates type 2 diabetes in high-fat diet and streptozotocin-induced diabetic mice in association with modulating the gut microbiota. Food Funct. 2021, 12, 6363–6373. [Google Scholar] [CrossRef] [PubMed]
- Tannock, G.W.; Lawley, B.; Munro, K.; Gowri Pathmanathan, S.; Zhou, S.J.; Makrides, M.; Gibson, R.A.; Sullivan, T.; Prosser, C.G.; Lowry, D. Comparison of the compositions of the stool microbiotas of infants fed goat milk formula, cow milk-based formula, or breast milk. Appl. Environ. Microbiol. 2013, 79, 3040–3048. [Google Scholar] [CrossRef] [PubMed]
- Turroni, F.; Bottacini, F.; Foroni, E.; Mulder, I.; Kim, J.-H.; Zomer, A.; Sánchez, B.; Bidossi, A.; Ferrarini, A.; Giubellini, V. Genome analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways for host-derived glycan foraging. Proc. Natl. Acad. Sci. USA 2010, 107, 19514–19519. [Google Scholar] [CrossRef]
- Furet, J.-P.; Kong, L.-C.; Tap, J.; Poitou, C.; Basdevant, A.; Bouillot, J.-L.; Mariat, D.; Corthier, G.; Doré, J.; Henegar, C. Differential adaptation of human gut microbiota to bariatric surgery–induced weight loss: Links with metabolic and low-grade inflammation markers. Diabetes 2010, 59, 3049–3057. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Kurpad, A.V.; Puri, S. Potential of probiotics in hypercholesterolemia: A meta-analysis. Indian J. Public Health 2016, 60, 280–286. [Google Scholar] [PubMed]
- Le, T.K.C.; Hosaka, T.; Le, T.T.T.; Nguyen, T.G.; Tran, Q.B.; Le, T.H.H.; Da Pham, X. Oral administration of Bifidobacterium spp. improves insulin resistance, induces adiponectin, and prevents inflammatory adipokine expressions. Biomed. Res. 2014, 35, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Mencarelli, A.; Cipriani, S.; Renga, B.; Bruno, A.; D’Amore, C.; Distrutti, E.; Fiorucci, S. VSL#3 resets insulin signaling and protects against NASH and atherosclerosis in a model of genetic dyslipidemia and intestinal inflammation. PLoS ONE 2012, 7, e45425. [Google Scholar]
- Amar, J.; Chabo, C.; Waget, A.; Klopp, P.; Vachoux, C.; Bermúdez-Humarán, L.G.; Smirnova, N.; Bergé, M.; Sulpice, T.; Lahtinen, S. Intestinal mucosal adherence and translocation of commensal bacteria at the early onset of type 2 diabetes: Molecular mechanisms and probiotic treatment. EMBO Mol. Med. 2011, 3, 559–572. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Zhu, H.; Gao, F.; Qian, Z.; Mao, W.; Yin, Y.; Tan, J.; Chen, D. Antidiabetic effects of selenium-enriched Bifidobacterium longum DD98 in type 2 diabetes model of mice. Food Funct. 2020, 11, 6528–6541. [Google Scholar] [CrossRef]
- Othman, M.B.; Sakamoto, K. Effect of inactivated Bifidobacterium longum intake on obese diabetes model mice (TSOD). Food Res. Int. 2020, 129, 108792. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).