
Causal Association between Circulating Metabolites and Dementia: A Mendelian Randomization Study

Abstract
1. Introduction
2. Materials and Methods
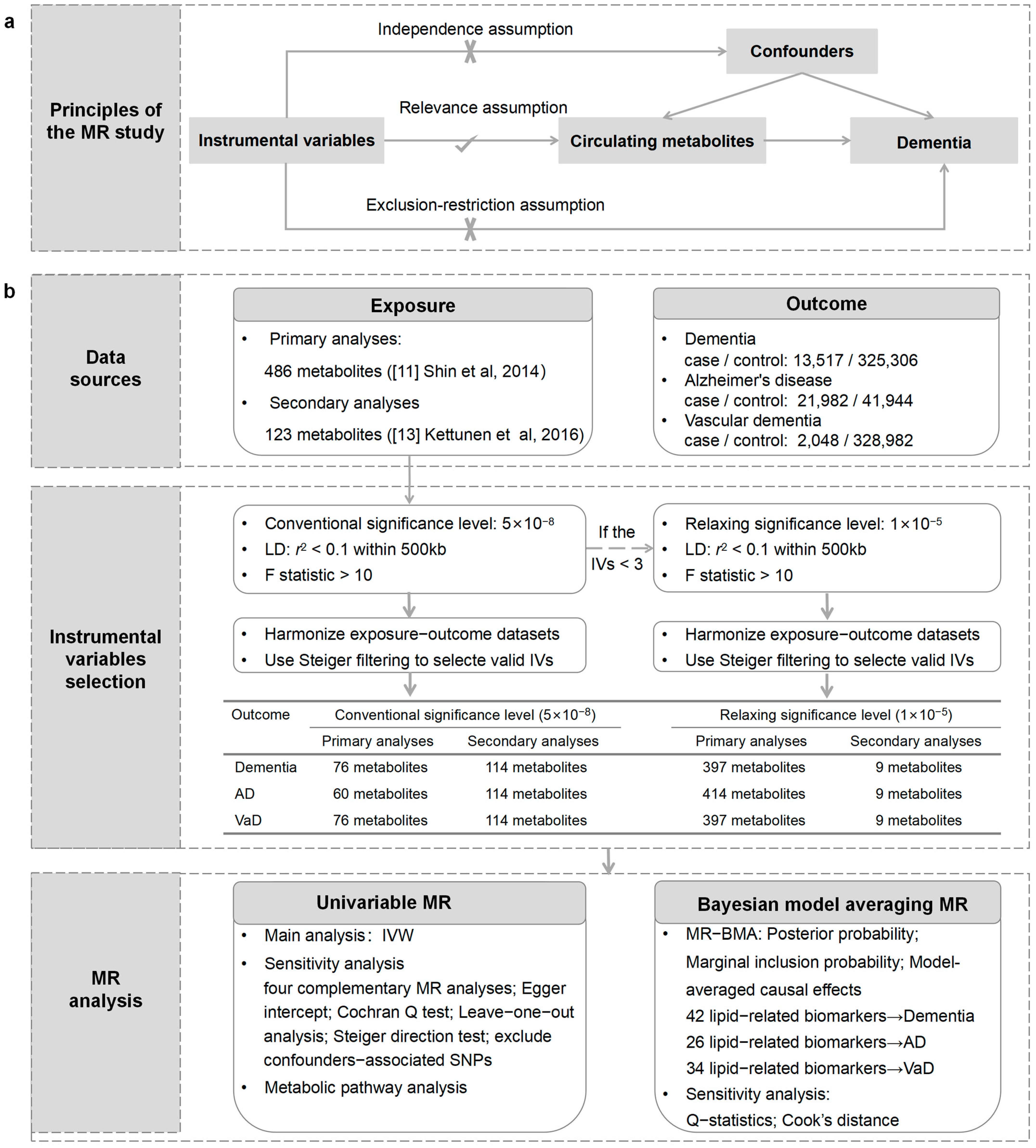
2.1. Study Design
2.2. GWAS Data Sources for Circulating Metabolites and Dementia
2.3. Selection of IVs
2.4. Univariable MR
2.5. MR-BMA Analysis
2.6. Statistical Analysis
2.7. Metabolic Pathway Analysis
3. Results
3.1. Strength of the IVs
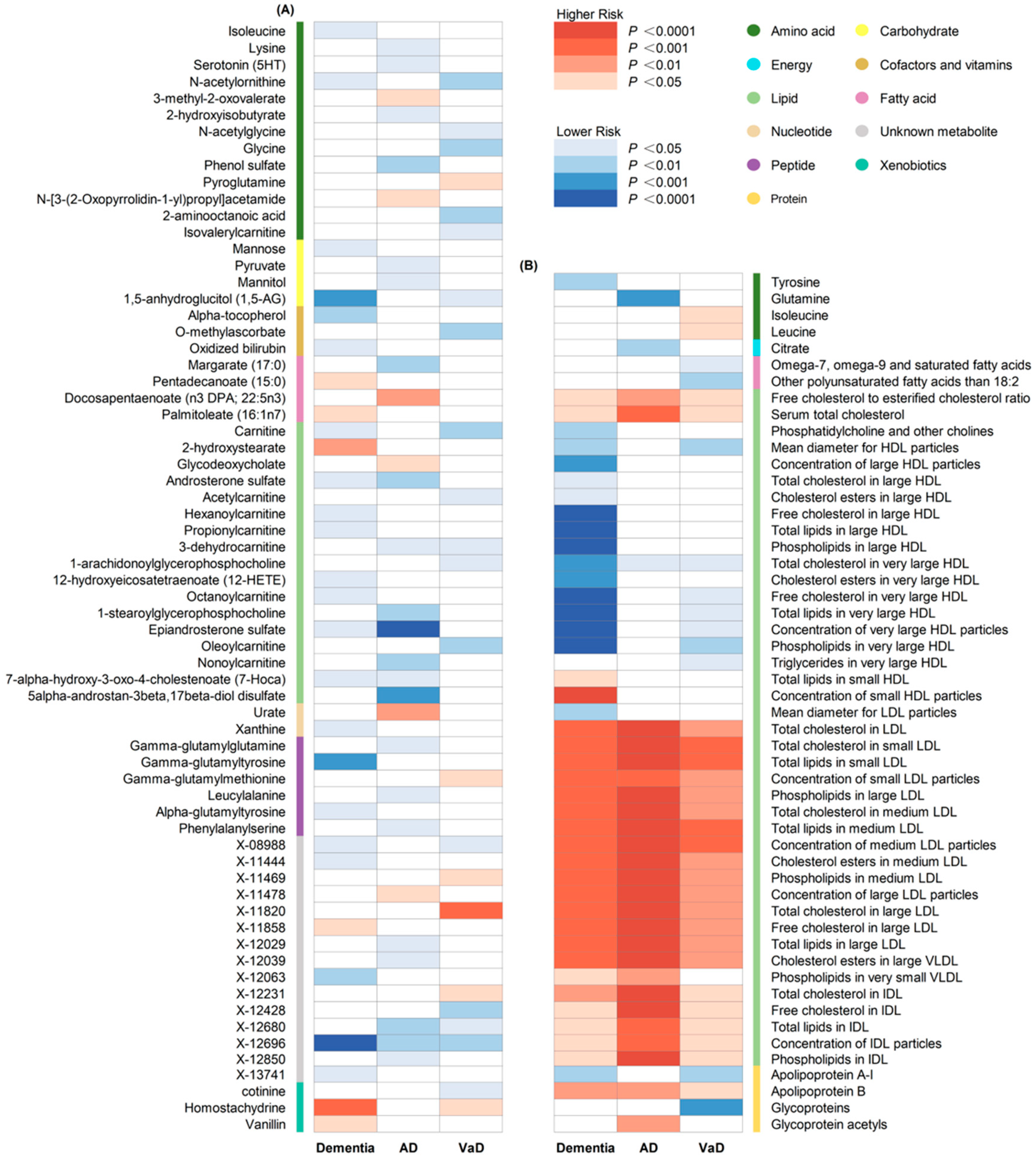
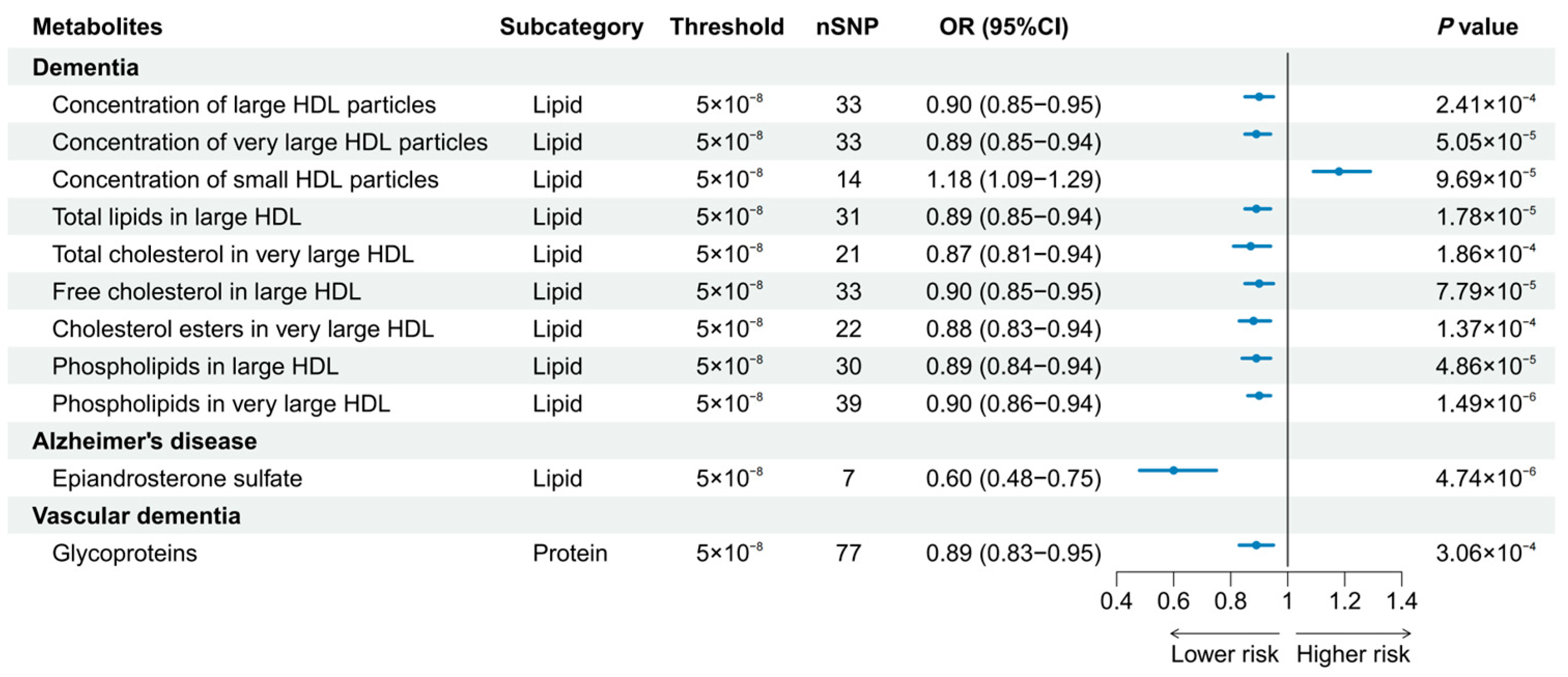
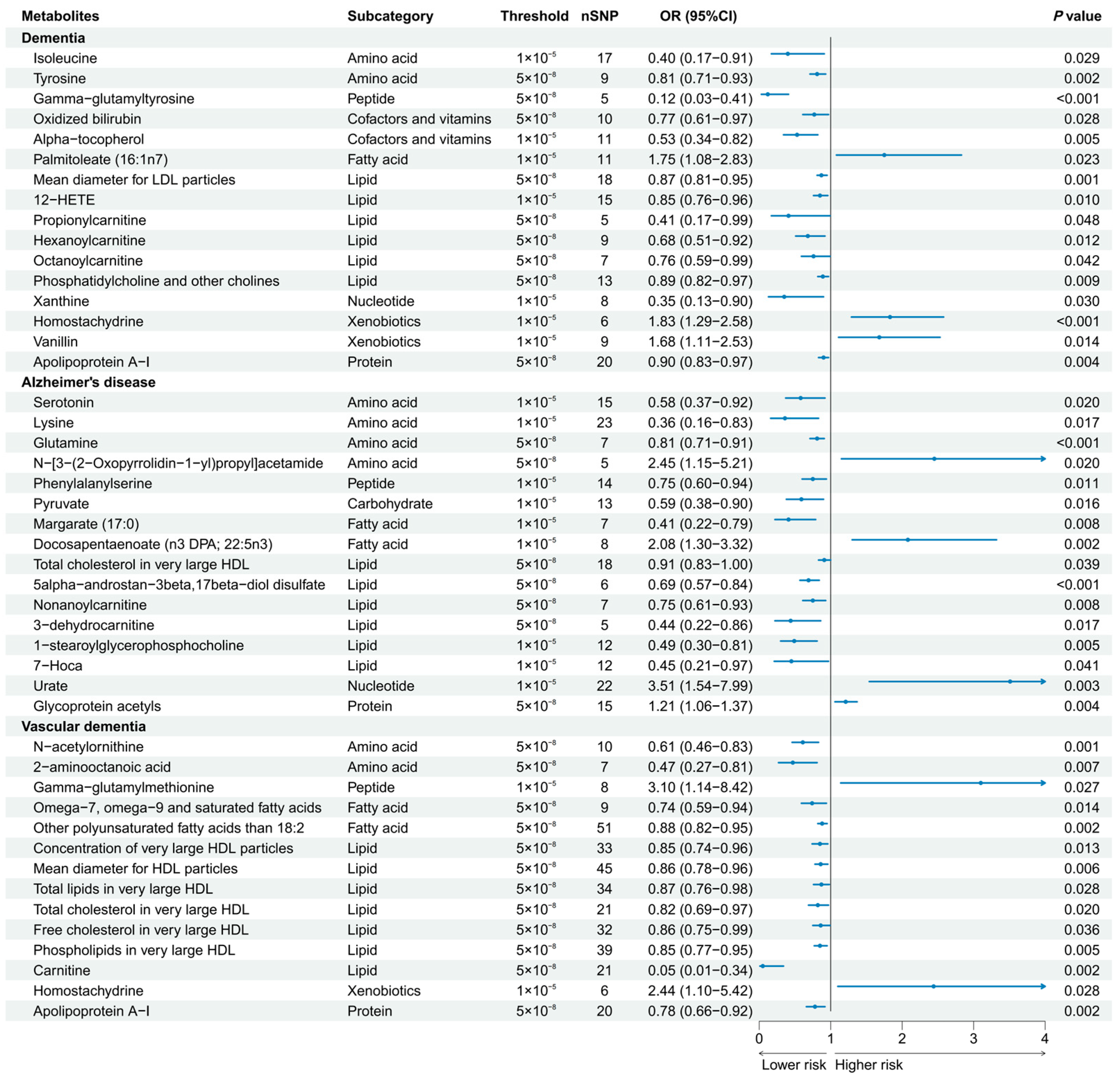
3.2. Univariable MR Analyses
3.3. MR-BMA Analyses
3.4. Metabolic Pathway Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grande, G.; Qiu, C.; Fratiglioni, L. Prevention of dementia in an ageing world: Evidence and biological rationale. Ageing Res. Rev. 2020, 64, 101045. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Dementia Forecasting Collaborators. Estimation of the global prevalence of dementia in 2019 and forecasted prevalence in 2050: An analysis for the Global Burden of Disease Study 2019. Lancet Public Health 2022, 7, e105–e125. [Google Scholar] [CrossRef] [PubMed]
- Lefevre-Arbogast, S.; Wagner, M.; Proust-Lima, C.; Samieri, C. Nutrition and Metabolic Profiles in the Natural History of Dementia: Recent Insights from Systems Biology and Life Course Epidemiology. Curr. Nutr. Rep. 2019, 8, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Lin, Y.; Li, X.; Driver, J.A.; Liang, L. Shared genetic architecture between metabolic traits and Alzheimer’s disease: A large-scale genome-wide cross-trait analysis. Hum. Genet. 2019, 138, 271–285. [Google Scholar] [CrossRef]
- de la Monte, S.M.; Tong, M. Brain metabolic dysfunction at the core of Alzheimer’s disease. Biochem. Pharmacol. 2014, 88, 548–559. [Google Scholar] [CrossRef]
- Zhang, X.; Hu, W.; Wang, Y.; Wang, W.; Liao, H.; Zhang, X.; Kiburg, K.V.; Shang, X.; Bulloch, G.; Huang, Y.; et al. Plasma metabolomic profiles of dementia: A prospective study of 110,655 participants in the UK Biobank. BMC Med. 2022, 20, 252. [Google Scholar] [CrossRef] [PubMed]
- Proitsi, P.; Kuh, D.; Wong, A.; Maddock, J.; Bendayan, R.; Wulaningsih, W.; Hardy, R.; Richards, M. Lifetime cognition and late midlife blood metabolites: Findings from a British birth cohort. Transl. Psychiatry 2018, 8, 203. [Google Scholar] [CrossRef]
- van der Lee, S.J.; Teunissen, C.E.; Pool, R.; Shipley, M.J.; Teumer, A.; Chouraki, V.; Melo van Lent, D.; Tynkkynen, J.; Fischer, K.; Hernesniemi, J.; et al. Circulating metabolites and general cognitive ability and dementia: Evidence from 11 cohort studies. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2018, 14, 707–722. [Google Scholar] [CrossRef]
- Tynkkynen, J.; Chouraki, V.; van der Lee, S.J.; Hernesniemi, J.; Yang, Q.; Li, S.; Beiser, A.; Larson, M.G.; Sääksjärvi, K.; Shipley, M.J.; et al. Association of branched-chain amino acids and other circulating metabolites with risk of incident dementia and Alzheimer’s disease: A prospective study in eight cohorts. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2018, 14, 723–733. [Google Scholar] [CrossRef]
- Sanderson, E.; Richardson, T.G.; Morris, T.T.; Tilling, K.; Davey Smith, G. Estimation of causal effects of a time-varying exposure at multiple time points through multivariable mendelian randomization. PLoS Genet. 2022, 18, e1010290. [Google Scholar] [CrossRef]
- Shin, S.Y.; Fauman, E.B.; Petersen, A.K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.P.; et al. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Skrivankova, V.W.; Richmond, R.C.; Woolf, B.A.R.; Yarmolinsky, J.; Davies, N.M.; Swanson, S.A.; VanderWeele, T.J.; Higgins, J.P.T.; Timpson, N.J.; Dimou, N.; et al. Strengthening the Reporting of Observational Studies in Epidemiology Using Mendelian Randomization: The STROBE-MR Statement. Jama 2021, 326, 1614–1621. [Google Scholar] [CrossRef] [PubMed]
- Kettunen, J.; Demirkan, A.; Wurtz, P.; Draisma, H.H.; Haller, T.; Rawal, R.; Vaarhorst, A.; Kangas, A.J.; Lyytikainen, L.P.; Pirinen, M.; et al. Genome-wide study for circulating metabolites identifies 62 loci and reveals novel systemic effects of LPA. Nat. Commun. 2016, 7, 11122. [Google Scholar] [CrossRef]
- Kurki, M.I.; Karjalainen, J.; Palta, P.; Sipilä, T.P.; Kristiansson, K.; Donner, K.M.; Reeve, M.P.; Laivuori, H.; Aavikko, M.; Kaunisto, M.A.; et al. FinnGen provides genetic insights from a well-phenotyped isolated population. Nature 2023, 613, 508–518. [Google Scholar] [CrossRef]
- Kunkle, B.W.; Grenier-Boley, B.; Sims, R.; Bis, J.C.; Damotte, V.; Naj, A.C.; Boland, A.; Vronskaya, M.; van der Lee, S.J.; Amlie-Wolf, A.; et al. Genetic meta-analysis of diagnosed Alzheimer’s disease identifies new risk loci and implicates Abeta, tau, immunity and lipid processing. Nat. Genet. 2019, 51, 414–430. [Google Scholar] [CrossRef]
- Burgess, S.; Davies, N.M.; Thompson, S.G. Bias due to participant overlap in two-sample Mendelian randomization. Genet. Epidemiol. 2016, 40, 597–608. [Google Scholar] [CrossRef]
- Sanna, S.; van Zuydam, N.R.; Mahajan, A.; Kurilshikov, A.; Vich Vila, A.; Vosa, U.; Mujagic, Z.; Masclee, A.A.M.; Jonkers, D.; Oosting, M.; et al. Causal relationships among the gut microbiome, short-chain fatty acids and metabolic diseases. Nat. Genet. 2019, 51, 600–605. [Google Scholar] [CrossRef]
- Yang, J.; Yan, B.; Zhao, B.; Fan, Y.; He, X.; Yang, L.; Ma, Q.; Zheng, J.; Wang, W.; Bai, L.; et al. Assessing the Causal Effects of Human Serum Metabolites on 5 Major Psychiatric Disorders. Schizophr. Bull. 2020, 46, 804–813. [Google Scholar] [CrossRef]
- Hemani, G.; Tilling, K.; Davey Smith, G. Orienting the causal relationship between imprecisely measured traits using GWAS summary data. PLoS Genet. 2017, 13, e1007081. [Google Scholar]
- Pierce, B.L.; Ahsan, H.; Vanderweele, T.J. Power and instrument strength requirements for Mendelian randomization studies using multiple genetic variants. Int. J. Epidemiol. 2011, 40, 740–752. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Avoiding bias from weak instruments in Mendelian randomization studies. Int. J. Epidemiol. 2011, 40, 755–764. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Li, X.; Wu, S.; Tian, Y.; Zhang, Y.; Wei, Z.; Jin, Z.; Li, X.; Chen, X.; Chen, W.X. Assessing the causal association between human blood metabolites and the risk of epilepsy. J. Transl. Med. 2022, 20, 437. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.; Huang, Y.; Hu, J.; Shao, Z. Mendelian randomization study of inflammatory bowel disease and bone mineral density. BMC Med. 2020, 18, 312. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Shen, X.; Pan, W. Constrained maximum likelihood-based Mendelian randomization robust to both correlated and uncorrelated pleiotropic effects. Am. J. Hum. Genet. 2021, 108, 1251–1269. [Google Scholar] [CrossRef]
- Yeung, C.H.C.; Au Yeung, S.L.; Kwok, M.K.; Zhao, J.V.; Schooling, C.M. The influence of growth and sex hormones on risk of alzheimer’s disease: A mendelian randomization study. Eur. J. Epidemiol. 2023, 38, 745–755. [Google Scholar] [CrossRef]
- Zuber, V.; Colijn, J.M.; Klaver, C.; Burgess, S. Selecting likely causal risk factors from high-throughput experiments using multivariable Mendelian randomization. Nat. Commun. 2020, 11, 29. [Google Scholar] [CrossRef]
- Levin, M.G.; Zuber, V.; Walker, V.M.; Klarin, D.; Lynch, J.; Malik, R.; Aday, A.W.; Bottolo, L.; Pradhan, A.D.; Dichgans, M.; et al. Prioritizing the Role of Major Lipoproteins and Subfractions as Risk Factors for Peripheral Artery Disease. Circulation 2021, 144, 353–364. [Google Scholar] [CrossRef]
- Hottman, D.A.; Chernick, D.; Cheng, S.; Wang, Z.; Li, L. HDL and cognition in neurodegenerative disorders. Neurobiol. Dis. 2014, 72 Pt A, 22–36. [Google Scholar] [CrossRef]
- Kjeldsen, E.W.; Thomassen, J.Q.; Juul Rasmussen, I.; Nordestgaard, B.G.; Tybjærg-Hansen, A.; Frikke-Schmidt, R. Plasma high-density lipoprotein cholesterol and risk of dementia: Observational and genetic studies. Cardiovasc. Res. 2022, 118, 1330–1343. [Google Scholar] [CrossRef]
- Sáiz-Vazquez, O.; Puente-Martínez, A.; Ubillos-Landa, S.; Pacheco-Bonrostro, J.; Santabárbara, J. Cholesterol and Alzheimer’s Disease Risk: A Meta-Meta-Analysis. Brain Sci. 2020, 10, 386. [Google Scholar] [CrossRef]
- Li, R.; Chen, J.X.; Lu, Q.; Geng, T.T.; Xia, P.F.; Wang, Y.; Chen, L.K.; Shan, Z.L.; Pan, A.; Liu, G. Associations of lipoprotein subclasses with risk of all-cause and cardiovascular disease mortality in individuals with type 2 diabetes: A prospective cohort study. Diabetes Obes. Metab. 2023, 25, 3259–3267. [Google Scholar] [CrossRef] [PubMed]
- Proitsi, P.; Lupton, M.K.; Velayudhan, L.; Newhouse, S.; Fogh, I.; Tsolaki, M.; Daniilidou, M.; Pritchard, M.; Kloszewska, I.; Soininen, H.; et al. Genetic predisposition to increased blood cholesterol and triglyceride lipid levels and risk of Alzheimer disease: A Mendelian randomization analysis. PLoS Med. 2014, 11, e1001713. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, S.D.; Mukherjee, S.; Sharp, S.J.; Proitsi, P.; Lotta, L.A.; Day, F.; Perry, J.R.; Boehme, K.L.; Walter, S.; Kauwe, J.S.; et al. Associations between Potentially Modifiable Risk Factors and Alzheimer Disease: A Mendelian Randomization Study. PLoS Med. 2015, 12, e1001841. [Google Scholar] [CrossRef] [PubMed]
- Fazio, S.; Pamir, N. HDL Particle Size and Functional Heterogeneity. Circ. Res. 2016, 119, 704–707. [Google Scholar] [CrossRef] [PubMed]
- Kontush, A.; Lhomme, M.; Chapman, M.J. Unraveling the complexities of the HDL lipidome. J. Lipid Res. 2013, 54, 2950–2963. [Google Scholar] [CrossRef]
- Turri, M.; Marchi, C.; Adorni, M.P.; Calabresi, L.; Zimetti, F. Emerging role of HDL in brain cholesterol metabolism and neurodegenerative disorders. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159123. [Google Scholar] [CrossRef]
- Moayyeri, A.; Cheung, C.L.; Tan, K.C.; Morris, J.A.; Cerani, A.; Mohney, R.P.; Richards, J.B.; Hammond, C.; Spector, T.D.; Menni, C. Metabolomic Pathways to Osteoporosis in Middle-Aged Women: A Genome-Metabolome-Wide Mendelian Randomization Study. J. Bone Min. Res. 2018, 33, 643–650. [Google Scholar] [CrossRef]
- Sun, L.; Guo, D.; Jia, Y.; Shi, M.; Yang, P.; Wang, Y.; Liu, F.; Chen, G.C.; Zhang, Y.; Zhu, Z. Association between Human Blood Metabolome and the Risk of Alzheimer’s Disease. Ann. Neurol. 2022, 92, 756–767. [Google Scholar] [CrossRef]
- Pérez-Jiménez, M.M.; Monje-Moreno, J.M.; Brokate-Llanos, A.M.; Venegas-Calerón, M.; Sánchez-García, A.; Sansigre, P.; Valladares, A.; Esteban-García, S.; Suárez-Pereira, I.; Vitorica, J.; et al. Steroid hormones sulfatase inactivation extends lifespan and ameliorates age-related diseases. Nat. Commun. 2021, 12, 49. [Google Scholar] [CrossRef]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Haran, J.P.; Bhattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s Disease Microbiome Is Associated with Dysregulation of the Anti-Inflammatory P-Glycoprotein Pathway. mBio 2019, 10, e00632-19. [Google Scholar] [CrossRef]
- Llorens, F.; Hermann, P.; Villar-Piqué, A.; Diaz-Lucena, D.; Nägga, K.; Hansson, O.; Santana, I.; Schmitz, M.; Schmidt, C.; Varges, D.; et al. Cerebrospinal fluid lipocalin 2 as a novel biomarker for the differential diagnosis of vascular dementia. Nat. Commun. 2020, 11, 619. [Google Scholar] [CrossRef]
- Horgusluoglu, E.; Neff, R.; Song, W.M.; Wang, M.; Wang, Q.; Arnold, M.; Krumsiek, J.; Galindo-Prieto, B.; Ming, C.; Nho, K.; et al. Integrative metabolomics-genomics approach reveals key metabolic pathways and regulators of Alzheimer’s disease. Alzheimer’s Dement. J. Alzheimer’s Assoc. 2022, 18, 1260–1278. [Google Scholar] [CrossRef] [PubMed]
- Glenn, J.M.; Madero, E.N.; Bott, N.T. Dietary Protein and Amino Acid Intake: Links to the Maintenance of Cognitive Health. Nutrients 2019, 11, 1315. [Google Scholar] [CrossRef] [PubMed]
- Reuter, S.E.; Evans, A.M. Carnitine and acylcarnitines: Pharmacokinetic, pharmacological and clinical aspects. Clin. Pharmacokinet. 2012, 51, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Nie, A.; Sun, B.; Fu, Z.; Yu, D. Roles of aminoacyl-tRNA synthetases in immune regulation and immune diseases. Cell Death Dis. 2019, 10, 901. [Google Scholar] [CrossRef]
- Polis, B.; Samson, A.O. Role of the metabolism of branched-chain amino acids in the development of Alzheimer’s disease and other metabolic disorders. Neural Regen. Res. 2020, 15, 1460–1470. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Castellano-Escuder, P.; Lefèvre-Arbogast, S.; Low, D.Y.; Du Preez, A.; Ruigrok, S.R.; Lee, H.; Helmer, C.; Pallàs, M.; Urpi-Sarda, M.; et al. Apolipoprotein E and sex modulate fatty acid metabolism in a prospective observational study of cognitive decline. Alzheimers Res. Ther. 2022, 14, 1. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite Traits | Rank | MIP | Average Effect | Empirical p Values |
|---|---|---|---|---|
| Dementia | ||||
| Concentration of very large HDL particles | 1 | 0.139 | −0.025 | 0.002 |
| Mean diameter for HDL particles | 2 | 0.127 | −0.018 | 0.005 |
| Free cholesterol in very large HDL particles | 3 | 0.116 | −0.02 | 0.097 |
| Phospholipids in very large HDL particles | 4 | 0.110 | −0.015 | 0.028 |
| Total lipids in very large HDL particles | 5 | 0.084 | −0.007 | 0.059 |
| AD | ||||
| Total cholesterol in very large HDL particles | 1 | 0.579 | −0.094 | 0.005 |
| Serum total cholesterol | 2 | 0.101 | −0.031 | 0.084 |
| Free cholesterol to esterified cholesterol ratio | 3 | 0.062 | 0.012 | 0.291 |
| Glycoprotein acetyls | 4 | 0.044 | 0.005 | 0.628 |
| Phospholipids in medium LDL particles | 5 | 0.044 | 0.007 | 0.985 |
| VaD | ||||
| Omega-7, omega-9 and saturated fatty acids | 1 | 0.311 | −0.13 | 0.012 |
| Serum total cholesterol | 2 | 0.186 | −0.103 | 0.001 |
| Triglycerides in very large HDL particles | 3 | 0.101 | −0.02 | 0.026 |
| Total cholesterol in medium LDL particles | 4 | 0.101 | 0.039 | 0.012 |
| Total cholesterol in LDL particles | 5 | 0.1 | 0.041 | 0.007 |
| Model | Posterior Probability | Causal Estimate |
|---|---|---|
| Dementia | ||
| Mean diameter for HDL particles | 0.055 | −0.109 |
| Concentration of very large HDL particles | 0.054 | −0.124 |
| Phospholipids in very large HDL particles | 0.047 | −0.112 |
| Free cholesterol in very large HDL particles | 0.042 | −0.126 |
| Total lipids in very large HDL particles | 0.029 | −0.115 |
| Cholesterol esters in large HDL particles | 0.028 | −0.113 |
| Concentration of large HDL particles | 0.027 | −0.112 |
| Total lipids in large HDL particles | 0.026 | −0.112 |
| Total cholesterol in large HDL particles | 0.025 | −0.114 |
| Free cholesterol in large HDL particles | 0.023 | −0.114 |
| Concentration of small HDL particles | 0.020 | 0.161 |
| AD | ||
| Total cholesterol in very large HDL particles | 0.362 | −0.161 |
| VaD | ||
| Triglycerides in very large HDL particles | 0.028 | −0.170 |
| Omega-7, omega-9 and saturated fatty acids | 0.025 | −0.246 |
| Metabolic Pathway | Outcome | Database | Metabolites Involved | p Value |
|---|---|---|---|---|
| Aminoacyl-tRNA biosynthesis | Dementia | KEGG | Isoleucine, tyrosine | 8.85 × 10−3 |
| Aminoacyl-tRNA biosynthesis | AD | KEGG | Glutamine, lysine | 2.33 × 10−2 |
| Aminoacyl-tRNA biosynthesis | VaD | KEGG | Glycine, isoleucine, leucine | 1.09 × 10−4 |
| Valine, leucine and isoleucine biosynthesis | Dementia | KEGG, SMPDB | Isoleucine | 2.56 × 10−2 |
| Valine, leucine and isoleucine biosynthesis | AD | KEGG, SMPDB | 3-methyl-2-oxopentanoic acid | 4.06 × 10−2 |
| Valine, leucine and isoleucine biosynthesis | VaD | KEGG, SMPDB | Leucine, isoleucine | 1.39 × 10−4 |
| Oxidation of branched chain fatty acids | Dementia | SMPDB | Carnitine, propionylcarnitine | 6.46 × 10−3 |
| Oxidation of branched chain fatty acids | VaD | SMPDB | Carnitine, acetylcarnitine | 4.36 × 10−3 |
| Phenylalanine, tyrosine and tryptophan biosynthesis | Dementia | KEGG, SMPDB | Tyrosine | 1.29 × 10−2 |
| Ubiquinone and other terpenoid-quinone biosynthesis | Dementia | KEGG, SMPDB | Tyrosine | 2.87 × 10−2 |
| Phenylalanine metabolism | Dementia | KEGG, SMPDB | Tyrosine | 3.19 × 10−2 |
| Arginine biosynthesis | Dementia | KEGG | Tyrosine | 4.44 × 10−2 |
| Glyoxylate and dicarboxylate metabolism | AD | KEGG | Citrate, pyruvate, glutamine | 4.00 × 10−4 |
| Citrate cycle (TCA cycle) | AD | KEGG, SMPDB | Pyruvate, citrate | 4.20 × 10−3 |
| Transfer of acetyl groups into mitochondria | AD | SMPDB | Pyruvate, citrate | 1.01 × 10−2 |
| Purine metabolism | AD | KEGG, SMPDB | Glutamine, urate | 4.12 × 10−2 |
| D-Glutamine and D-glutamate metabolism | AD | KEGG, SMPDB | Glutamine | 3.06 × 10−2 |
| Nitrogen metabolism | AD | KEGG, SMPDB | Glutamine | 3.06 × 10−2 |
| Valine, leucine and isoleucine degradation | VaD | KEGG, SMPDB | Leucine, isoleucine | 3.77 × 10−3 |
| Beta oxidation of very-long-chain fatty acids | VaD | SMPDB | Carnitine, acetylcarnitine | 1.50 × 10−3 |
| Carnitine synthesis | VaD | SMPDB | Carnitine, glycine | 2.29 × 10−3 |
| Arginine biosynthesis | VaD | KEGG | N-acetylornithine | 3.57 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.-M.; Qiu, C.-S.; Du, L.-Y.; Tang, X.-L.; Liao, D.-Q.; Xiong, Z.-Y.; Lai, S.-M.; Huang, H.-X.; Kuang, L.; Zhang, B.-Y.; et al. Causal Association between Circulating Metabolites and Dementia: A Mendelian Randomization Study. Nutrients 2024, 16, 2879. https://doi.org/10.3390/nu16172879
Li H-M, Qiu C-S, Du L-Y, Tang X-L, Liao D-Q, Xiong Z-Y, Lai S-M, Huang H-X, Kuang L, Zhang B-Y, et al. Causal Association between Circulating Metabolites and Dementia: A Mendelian Randomization Study. Nutrients. 2024; 16(17):2879. https://doi.org/10.3390/nu16172879
Chicago/Turabian StyleLi, Hong-Min, Cheng-Shen Qiu, Li-Ying Du, Xu-Lian Tang, Dan-Qing Liao, Zhi-Yuan Xiong, Shu-Min Lai, Hong-Xuan Huang, Ling Kuang, Bing-Yun Zhang, and et al. 2024. "Causal Association between Circulating Metabolites and Dementia: A Mendelian Randomization Study" Nutrients 16, no. 17: 2879. https://doi.org/10.3390/nu16172879
APA StyleLi, H.-M., Qiu, C.-S., Du, L.-Y., Tang, X.-L., Liao, D.-Q., Xiong, Z.-Y., Lai, S.-M., Huang, H.-X., Kuang, L., Zhang, B.-Y., & Li, Z.-H. (2024). Causal Association between Circulating Metabolites and Dementia: A Mendelian Randomization Study. Nutrients, 16(17), 2879. https://doi.org/10.3390/nu16172879