

Treatment with Gac Fruit Extract and Probiotics Reduces Serum Trimethylamine N-Oxide in Chronic Kidney Disease Rats
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Study
2.2. Measurement of Biochemical Indicators
2.3. Measurement of TMA Lyase (CutC)
2.4. Determination of Vascular Calcification
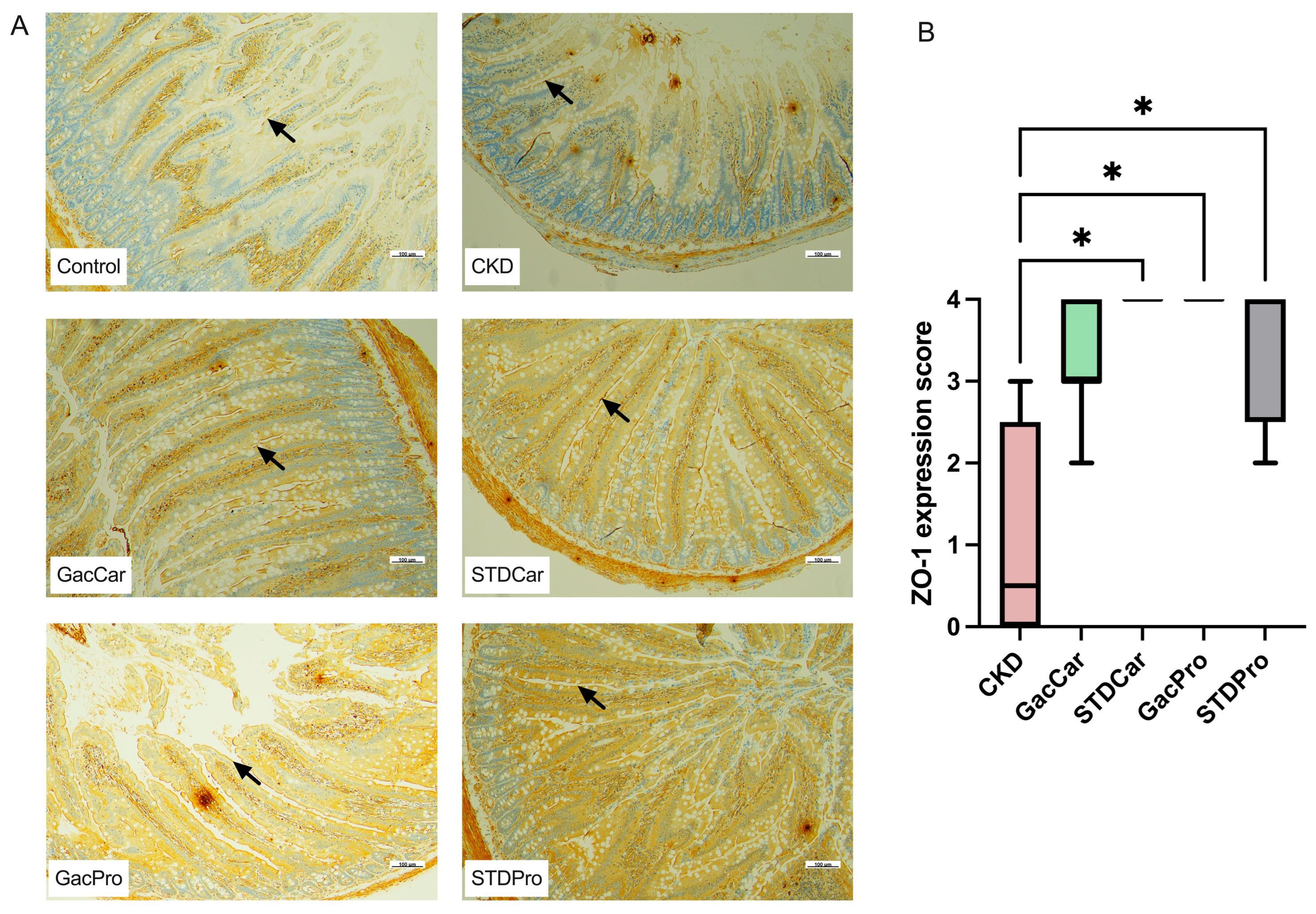
2.5. Immunocytochemistry to Determine Intestinal Zonula Occludens-1
2.6. Statistical Analysis
2.7. Ethical Considerations
3. Results
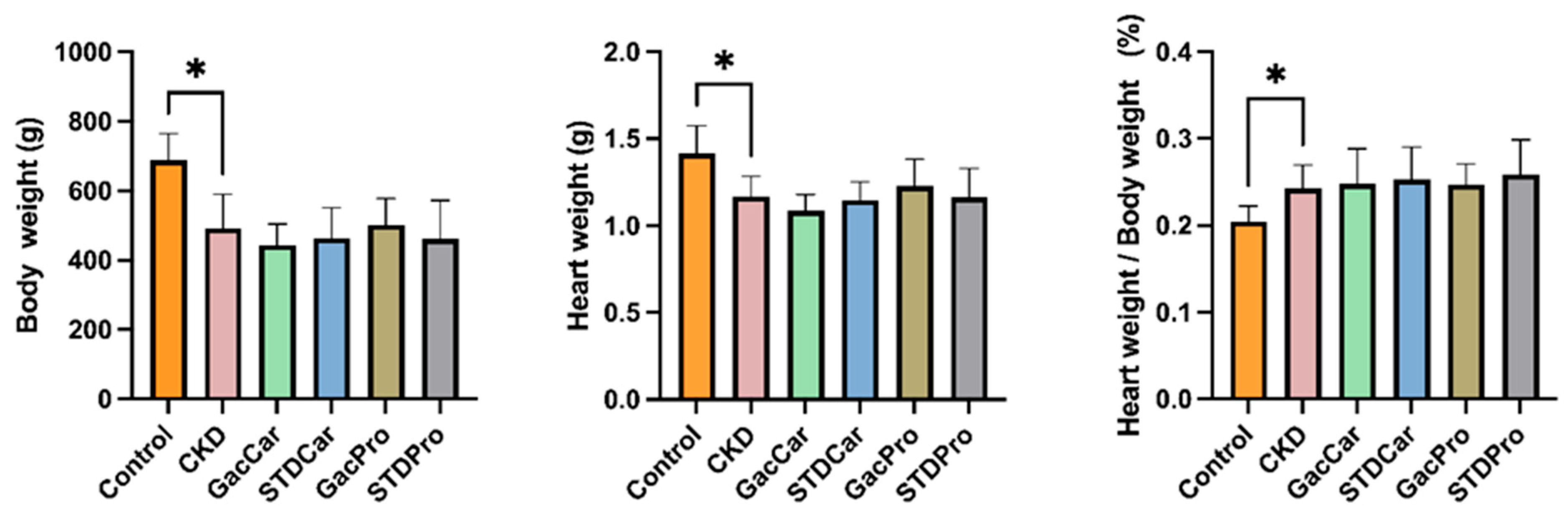
3.1. Body Weight and Heart Weight
3.2. Uremic Toxin
3.3. Histopathology of Cardiovascular Tissues and Immunohistochemistry of the Intestinal Tract
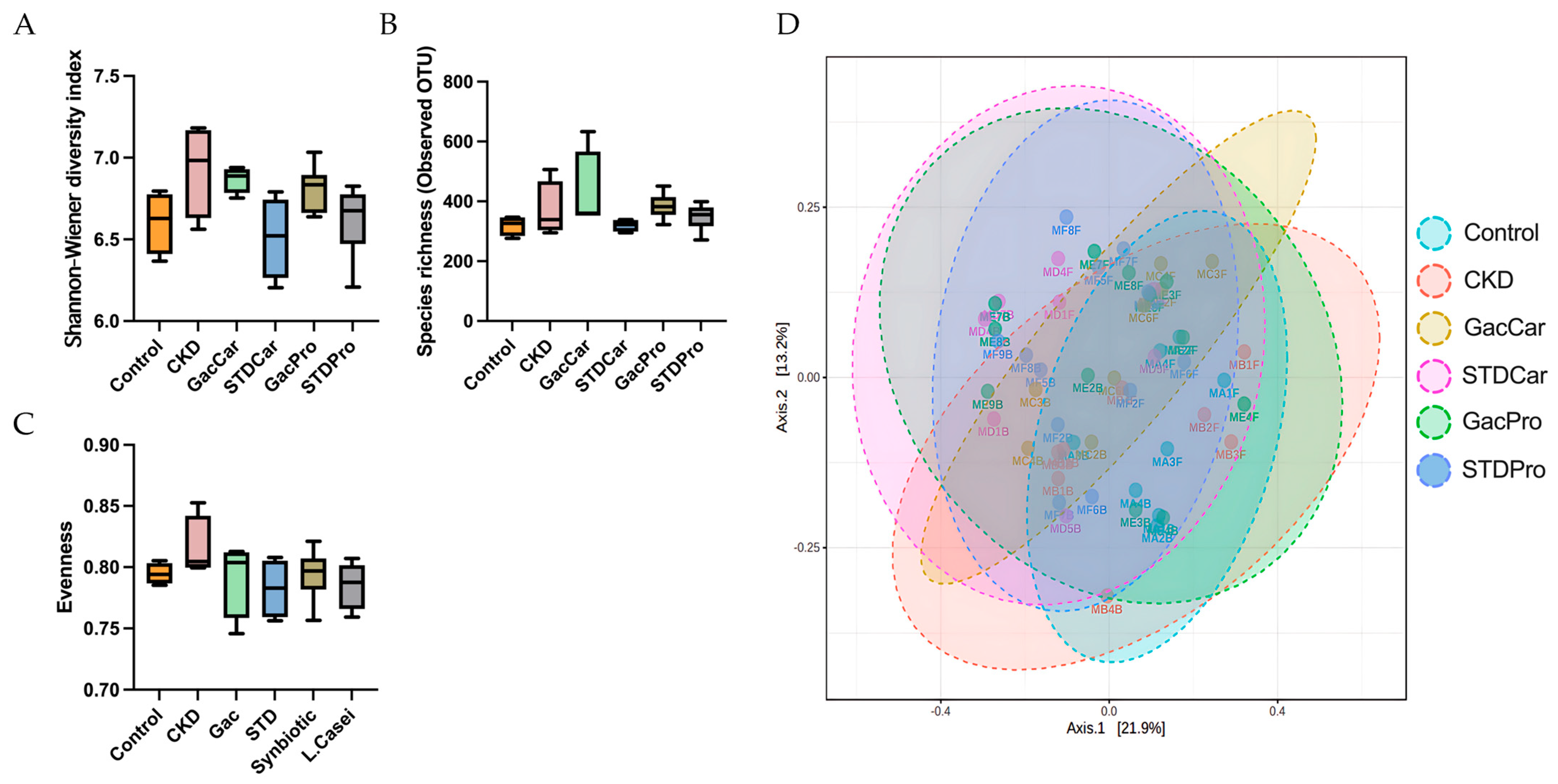
3.4. Gut Microbiome Studies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farrell, D.R.; Vassalotti, J.A. Screening, identifying, and treating chronic kidney disease: Why, who, when, how, and what? BMC Nephrol. 2024, 25, 34. [Google Scholar] [CrossRef] [PubMed]
- Gatarek, P.; Kaluzna-Czaplinska, J. Trimethylamine N-oxide (TMAO) in human health. Excli. J. 2021, 20, 301–319. [Google Scholar] [CrossRef]
- Zixin, Y.; Lulu, C.; Xiangchang, Z.; Qing, F.; Binjie, Z.; Chunyang, L.; Tai, R.; Dongsheng, O. TMAO as a potential biomarker and therapeutic target for chronic kidney disease: A review. Front. Pharmacol. 2022, 13, 929262. [Google Scholar] [CrossRef]
- Hsu, C.N.; Chang-Chien, G.P.; Lin, S.; Hou, C.Y.; Lu, P.C.; Tain, Y.L. Association of Trimethylamine, Trimethylamine N-oxide, and Dimethylamine with Cardiovascular Risk in Children with Chronic Kidney Disease. J. Clin. Med. 2020, 9, 336. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Lu, H.; Guo, J.; Zhang, M.; Zheng, H.; Liu, Y.; Liu, W. Gut microbiota-derived trimethylamine N-oxide is associated with the risk of all-cause and cardiovascular mortality in patients with chronic kidney disease: A systematic review and dose-response meta-analysis. Ann. Med. 2023, 55, 2215542. [Google Scholar] [CrossRef]
- Cantero, M.A.; Guedes, M.R.A.; Fernandes, R.; Lollo, P.C.B. Trimethylamine N-oxide reduction is related to probiotic strain specificity: A systematic review. Nutr. Res. 2022, 104, 29–35. [Google Scholar] [CrossRef]
- Ramireddy, L.; Tsen, H.Y.; Chiang, Y.C.; Hung, C.Y.; Wu, S.R.; Young, S.L.; Lin, J.S.; Huang, C.H.; Chiu, S.H.; Chen, C.C.; et al. Molecular Identification and Selection of Probiotic Strains Able to Reduce the Serum TMAO Level in Mice Challenged with Choline. Foods 2021, 10, 2931. [Google Scholar] [CrossRef]
- Abdulqader, A.; Ali, F.; Ismail, A.; Esa, N. Antioxidant compounds and capacities of Gac (Momordica cochinchinensis Spreng) fruits. Asian Pac. J. Trop. Biomed. 2019, 9, 158–167. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chu, Y.H.; Wang, C.C.; Wang, C.H.; Tain, Y.L.; Yang, H.W. Rapid Detection of Gut Microbial Metabolite Trimethylamine N-Oxide for Chronic Kidney Disease Prevention. Biosensors 2021, 11, 339. [Google Scholar] [CrossRef] [PubMed]
- Rath, S.; Rud, T.; Pieper, D.H.; Vital, M. Potential TMA-Producing Bacteria Are Ubiquitously Found in Mammalia. Front. Microbiol. 2019, 10, 2966. [Google Scholar] [CrossRef]
- Mulkearns-Hubert, E.E.; Esakov Rhoades, E.; Ben-Salem, S.; Bharti, R.; Hajdari, N.; Johnson, S.; Myers, A.; Smith, I.N.; Bandyopadhyay, S.; Eng, C.; et al. Targeting NANOG and FAK via Cx26-derived Cell-penetrating Peptides in Triple-negative Breast Cancer. Mol. Cancer Ther. 2024, 23, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Ketteler, M.; Block, G.A.; Evenepoel, P.; Fukagawa, M.; Herzog, C.A.; McCann, L.; Moe, S.M.; Shroff, R.; Tonelli, M.A.; Toussaint, N.D.; et al. Executive summary of the 2017 KDIGO Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD) Guideline Update: What’s changed and why it matters. Kidney Int. 2017, 92, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Neofytou, I.E.; Stamou, A.; Demopoulos, A.; Roumeliotis, S.; Zebekakis, P.; Liakopoulos, V.; Stamellou, E.; Dounousi, E. Vitamin K for Vascular Calcification in Kidney Patients: Still Alive and Kicking, but Still a Lot to Learn. Nutrients 2024, 16, 1798. [Google Scholar] [CrossRef]
- Yang, C.; Wei, Z.; Shi, W.; Xing, J.; Zhang, X. SNF472: A novel therapeutic agent for vascular calcification and calciphylaxis. J. Nephrol. 2024, 37, 851–863. [Google Scholar] [CrossRef]
- Oktaviono, Y.H.; Dyah Lamara, A.; Saputra, P.B.T.; Arnindita, J.N.; Pasahari, D.; Saputra, M.E.; Suasti, N.M.A. The roles of trimethylamine-N-oxide in atherosclerosis and its potential therapeutic aspect: A literature review. Biomol. Biomed. 2023, 23, 936–948. [Google Scholar] [CrossRef]
- Koeth, R.A.; Wang, Z.; Levison, B.S.; Buffa, J.A.; Org, E.; Sheehy, B.T.; Britt, E.B.; Fu, X.; Wu, Y.; Li, L.; et al. Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat. Med. 2013, 19, 576–585. [Google Scholar] [CrossRef]
- Martínez-del Campo, A.; Bodea, S.; Hamer, H.A.; Marks, J.A.; Haiser, H.J.; Turnbaugh, P.J.; Balskus, E.P. Characterization and detection of a widely distributed gene cluster that predicts anaerobic choline utilization by human gut bacteria. MBio 2015, 6, 10–1128. [Google Scholar] [CrossRef]
- Buysse, J.; Matunas, R.; Simpson, C.V. A Novel, Small Molecule Inhibitor of Gut Microbial Choline Trimethylamine Lyase (CutC) Slows the Loss of Kidney Function in a Rat Model of CKD: FR-PO1002. J. Am. Soc. Nephrol. 2023, 34, 684. [Google Scholar] [CrossRef]
- Mafra, D.; Kemp, J.A.; Borges, N.A.; Wong, M.; Stenvinkel, P. Gut Microbiota Interventions to Retain Residual Kidney Function. Toxins 2023, 15, 499. [Google Scholar] [CrossRef]
- Chen, R.; Zhu, D.; Yang, R.; Wu, Z.; Xu, N.; Chen, F.; Zhang, S.; Chen, H.; Li, M.; Hou, K. Gut microbiota diversity in middle-aged and elderly patients with end-stage diabetic kidney disease. Ann. Transl. Med. 2022, 10, 750. [Google Scholar] [CrossRef]
- He, X.; Sun, J.; Liu, C.; Yu, X.; Li, H.; Zhang, W.; Li, Y.; Geng, Y.; Wang, Z. Compositional Alterations of Gut Microbiota in Patients with Diabetic Kidney Disease and Type 2 Diabetes Mellitus. Diabetes Metab. Syndr. Obes. 2022, 15, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Prokopienko, A.J.; West, R.E., 3rd; Schrum, D.P.; Stubbs, J.R.; Leblond, F.A.; Pichette, V.; Nolin, T.D. Metabolic Activation of Flavin Monooxygenase-mediated Trimethylamine-N-Oxide Formation in Experimental Kidney Disease. Sci. Rep. 2019, 9, 15901. [Google Scholar] [CrossRef]
- Pelletier, C.C.; Croyal, M.; Ene, L.; Aguesse, A.; Billon-Crossouard, S.; Krempf, M.; Lemoine, S.; Guebre-Egziabher, F.; Juillard, L.; Soulage, C.O. Elevation of Trimethylamine-N-Oxide in Chronic Kidney Disease: Contribution of Decreased Glomerular Filtration Rate. Toxins 2019, 11, 635. [Google Scholar] [CrossRef]
- Zhang, W.; Miikeda, A.; Zuckerman, J.; Jia, X.; Charugundla, S.; Zhou, Z.; Kaczor-Urbanowicz, K.E.; Magyar, C.; Guo, F.; Wang, Z.; et al. Inhibition of microbiota-dependent TMAO production attenuates chronic kidney disease in mice. Sci. Rep. 2021, 11, 518. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Buffa, J.A.; Roberts, A.B.; Sangwan, N.; Skye, S.M.; Li, L.; Ho, K.J.; Varga, J.; DiDonato, J.A.; Tang, W.H.W.; et al. Targeted Inhibition of Gut Microbial Trimethylamine N-Oxide Production Reduces Renal Tubulointerstitial Fibrosis and Functional Impairment in a Murine Model of Chronic Kidney Disease. Arterioscler. Thromb Vasc. Biol. 2020, 40, 1239–1255. [Google Scholar] [CrossRef] [PubMed]
- Moludi, J.; Saiedi, S.; Ebrahimi, B.; Alizadeh, M.; Khajebishak, Y.; Ghadimi, S.S. Probiotics Supplementation on Cardiac Remodeling Following Myocardial Infarction: A Single-Center Double-Blind Clinical Study. J. Cardiovasc. Transl. Res. 2021, 14, 299–307. [Google Scholar] [CrossRef]
- Dong, Y.; Gai, Z.; Han, M.; Xu, J.; Zou, K. Reduction in Serum Concentrations of Uremic Toxins Driven by Bifidobacterium Longum Subsp. Longum BL21 is Associated with Gut Microbiota Changes in a Rat Model of Chronic Kidney Disease. Probiotics Antimicrob. Proteins 2024. [Google Scholar] [CrossRef]
- Karmazyn, M.; Gan, X.T. Probiotics as potential treatments to reduce myocardial remodelling and heart failure via the gut-heart axis: State-of-the-art review. Mol. Cell Biochem. 2023, 478, 2539–2551. [Google Scholar] [CrossRef]
- Rubin, L.P.; Ross, A.C.; Stephensen, C.B.; Bohn, T.; Tanumihardjo, S.A. Metabolic Effects of Inflammation on Vitamin A and Carotenoids in Humans and Animal Models. Adv. Nutr. 2017, 8, 197–212. [Google Scholar] [CrossRef]
- Bas-Bellver, C.; Andrés, C.; Seguí, L.; Barrera, C.; Jiménez-Hernández, N.; Artacho, A.; Betoret, N.; Gosalbes, M.J. Valorization of Persimmon and Blueberry Byproducts to Obtain Functional Powders: In Vitro Digestion and Fermentation by Gut Microbiota. J. Agric. Food Chem. 2020, 68, 8080–8090. [Google Scholar] [CrossRef]
- Zhao, B.; Wu, J.; Li, J.; Bai, Y.; Luo, Y.; Ji, B.; Xia, B.; Liu, Z.; Tan, X.; Lv, J.; et al. Lycopene Alleviates DSS-Induced Colitis and Behavioral Disorders via Mediating Microbes-Gut-Brain Axis Balance. J. Agric. Food Chem. 2020, 68, 3963–3975. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Song, Y.; Liu, H.; Wu, M.; Gong, H.; Lan, H.; Zheng, X. Gut microbiota regulation and anti-inflammatory effect of β-carotene in dextran sulfate sodium-stimulated ulcerative colitis in rats. J. Food Sci. 2021, 86, 2118–2130. [Google Scholar] [CrossRef] [PubMed]
- Eroglu, A.; Al’Abri, I.S.; Kopec, R.E.; Crook, N.; Bohn, T. Carotenoids and Their Health Benefits as Derived via Their Interactions with Gut Microbiota. Adv. Nutr. 2023, 14, 238–255. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cai, X.; Zhang, N.; Li, Y.; Mao, Y.; Ge, S.; Yao, Y.; Gao, H. Relation Between Dietary Carotenoid Intake, Serum Concentration, and Mortality Risk of CKD Patients Among US Adults: National Health and Nutrition Examination Survey 2001–2014. Front. Med. 2022, 9, 871767. [Google Scholar] [CrossRef]
- Shi, Y.; Xu, Y.; Zhou, W. Dietary carotenoids intake and sex differences in relation to chronic kidney disease a cross-sectional assessment in the NHANES study. BMC Public Health 2024, 24, 293. [Google Scholar] [CrossRef]
- Zhong, Q.; Piao, Y.; Yin, S.; Zhang, K. Association of serum lycopene concentrations with all-cause and cardiovascular mortality among individuals with chronic kidney disease: A cohort study. Front. Nutr. 2022, 9, 1048884. [Google Scholar] [CrossRef]
- Santamarina, A.B.; de Souza Mesquita, L.M.; Casagrande, B.P.; Sertorio, M.N.; Vitor de Souza, D.; Mennitti, L.V.; Ribeiro, D.A.; Estadella, D.; Ventura, S.P.M.; de Rosso, V.V.; et al. Supplementation of carotenoids from peach palm waste (Bactris gasipaes) obtained with an ionic liquid mediated process displays kidney anti-inflammatory and antioxidant outcomes. Food Chem. X 2022, 13, 100245. [Google Scholar] [CrossRef]
- Tang, W.H.; Wang, Z.; Kennedy, D.J.; Wu, Y.; Buffa, J.A.; Agatisa-Boyle, B.; Li, X.S.; Levison, B.S.; Hazen, S.L. Gut microbiota-dependent trimethylamine N-oxide (TMAO) pathway contributes to both development of renal insufficiency and mortality risk in chronic kidney disease. Circ. Res. 2015, 116, 448–455. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Primer | Sequence | Product Size (bp) |
|---|---|---|---|
| TMA lyase | Forward | TTYGCIGGITAYCARCCNTT | 275 |
| Reverse | TGNGGYTCIACRCAICCCAT | ||
| 16S rRNA | Forward | AGRGTTHGATYMTGGCTCAG | 177 |
| Reveres | TGCTGCCTCCCGTAGGAGT |
| Comparison Group | Phylum/Genus Level | Bacteria | Change in Relative Abundance (Week 15 vs. Week 0) | p Value | |
|---|---|---|---|---|---|
| CKD | Comparison Group | ||||
| CKD vs. Control | Phylum | Actinobacteriota | −0.062 (0.53) | −0.226 (0.38) | 0.043 * |
| Genus | Muribaculaceae | 2.355 (7.24) | −2.310 (6.05) | 0.021 * | |
| Genus | Colidextribacter | 0.829 (2.90) | 0.716 (0.81) | 0.043 * | |
| Genus | UCG008 | 0.379 (0.80) | −0.071 (0.49) | 0.021 * | |
| CKD vs. GacPro | Phylum | Actinobacteriota | −0.062 (0.53) | −0.377 (0.43) | 0.011 * |
| Comparison Group | Phylum/Genus Level | Bacteria | Change in Relative Abundance (Week 15 vs. Week 0) | p Value | |
|---|---|---|---|---|---|
| CKD | Comparison Group | ||||
| CKD vs. Control | Genus | Bacteroides | −1.190 (3.66) | 0.851 (1.39) | 0.021 * |
| Genus | Phascolarctobacterium | −0.346 (2.25) | 1.098 (2.08) | 0.021 * | |
| CKD vs. GacCar | Phylum | Desulfobacterota | −0.367 (0.47) | 0.204 (0.59) | 0.021 * |
| Phylum | Elusimicrobiota | 0.020 (1.21) | 2.464 (3.31) | 0.043 * | |
| Genus | Bacteroides | −1.190 (3.66) | −0.376 (0.19) | 0.021 * | |
| Genus | Phascolarctobacterium | −0.346 (2.25) | 1.098 (2.08) | 0.021 * | |
| Genus | Bacteroides | −1.190 (3.66) | −0.191 (0.59) | 0.021 * | |
| CKD vs. STDCar | Phylum | Verrucomicrobiota | −0.7660(1.11) | 0.296 (1.05) | 0.021 * |
| Phylum | Desulfobacterota | −0.367 (0.47) | 0.336 (1.50) | 0.021 * | |
| Phylum | Elusimicrobiota | 0.020 (1.21) | 0.027 (1.56) | 0.021 * | |
| Genus | Prevotellaceae_NK3B31_group | −6.089 (2.67) | −2.949 (4.05) | 0.021 * | |
| Genus | Akkermansia | −0.780 (1.11) | 0.296 (1.05) | 0.021 * | |
| CKD vs. GacPro | Phylum | Desulfobacterota | −0.367 (0.47) | 0.275 (0.36) | 0.011 * |
| Phylum | Campylobacterota | 0.014 (0.14) | 0.283 (0.44) | 0.033 * | |
| Genus | Bacteroides | −1.190 (3.66) | −0.027 (1.41) | 0.011 * | |
| Genus | Prevotellaceae_NK3B31_group | −6.089 (2.67) | 0.221 (3.19) | 0.019 * | |
| CKD vs. STDPro | Phylum | Campylobacterota | 0.014 (0.14) | 0.357 (0.24) | 0.011 * |
| Genus | Bacteroides | −1.190 (3.66) | −0.090 (1.28) | 0.011 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamkang, P.; Rattanachaisit, P.; Anegkamol, W.; Taweevisit, M.; Sapwarobol, S.; Tumwasorn, S.; Chuaypen, N.; Dissayabutra, T. Treatment with Gac Fruit Extract and Probiotics Reduces Serum Trimethylamine N-Oxide in Chronic Kidney Disease Rats. Nutrients 2024, 16, 2997. https://doi.org/10.3390/nu16172997
Kamkang P, Rattanachaisit P, Anegkamol W, Taweevisit M, Sapwarobol S, Tumwasorn S, Chuaypen N, Dissayabutra T. Treatment with Gac Fruit Extract and Probiotics Reduces Serum Trimethylamine N-Oxide in Chronic Kidney Disease Rats. Nutrients. 2024; 16(17):2997. https://doi.org/10.3390/nu16172997
Chicago/Turabian StyleKamkang, Panumas, Pakkapon Rattanachaisit, Weerapat Anegkamol, Mana Taweevisit, Suwimol Sapwarobol, Somying Tumwasorn, Natthaya Chuaypen, and Thasinas Dissayabutra. 2024. "Treatment with Gac Fruit Extract and Probiotics Reduces Serum Trimethylamine N-Oxide in Chronic Kidney Disease Rats" Nutrients 16, no. 17: 2997. https://doi.org/10.3390/nu16172997