
Preventive and Therapeutic Effects of Lactiplantibacillus plantarum HD02 and MD159 through Mast Cell Degranulation Inhibition in Mouse Models of Atopic Dermatitis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Identification of New Probiotic Strains
2.2. Mice
2.3. Mast Cell Degranulation Assay
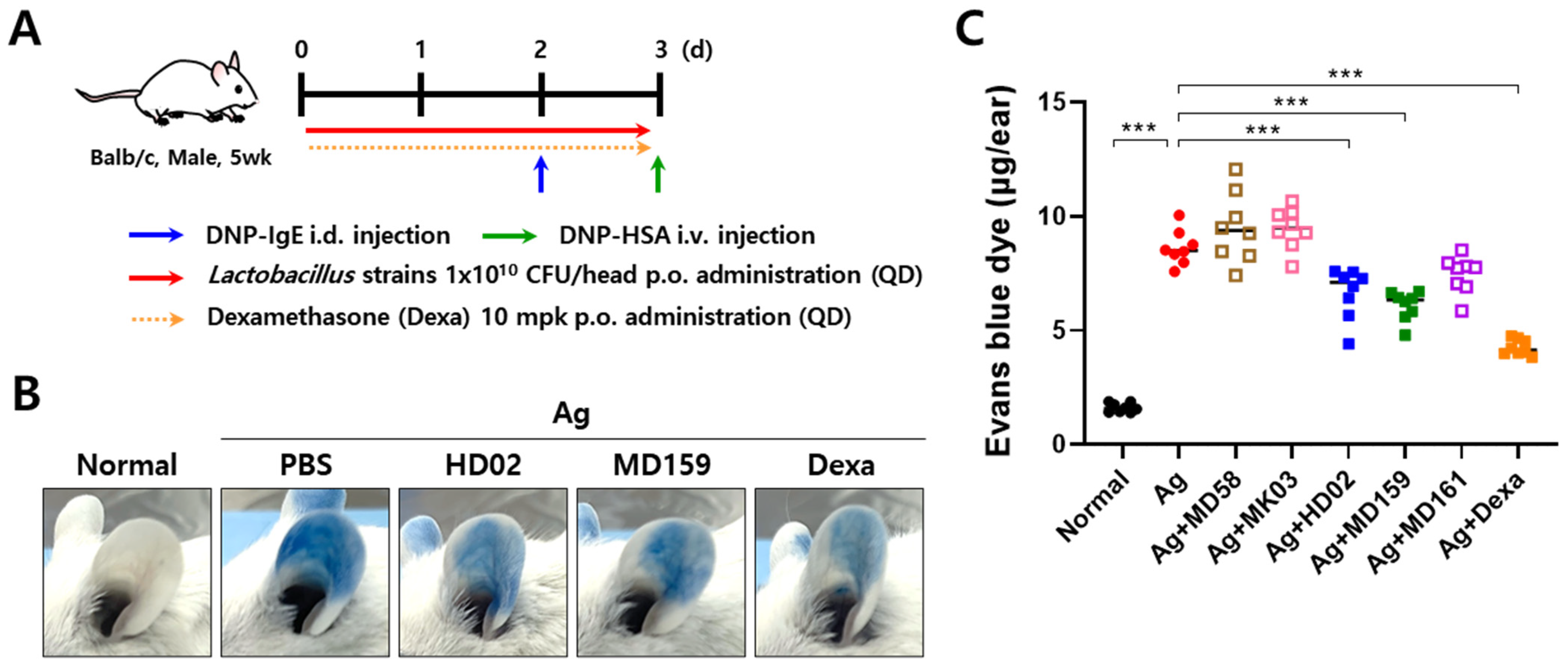
2.4. Passive Cutaneous Anaphylaxis (PCA) Model
2.5. MC903-Induced AD Model
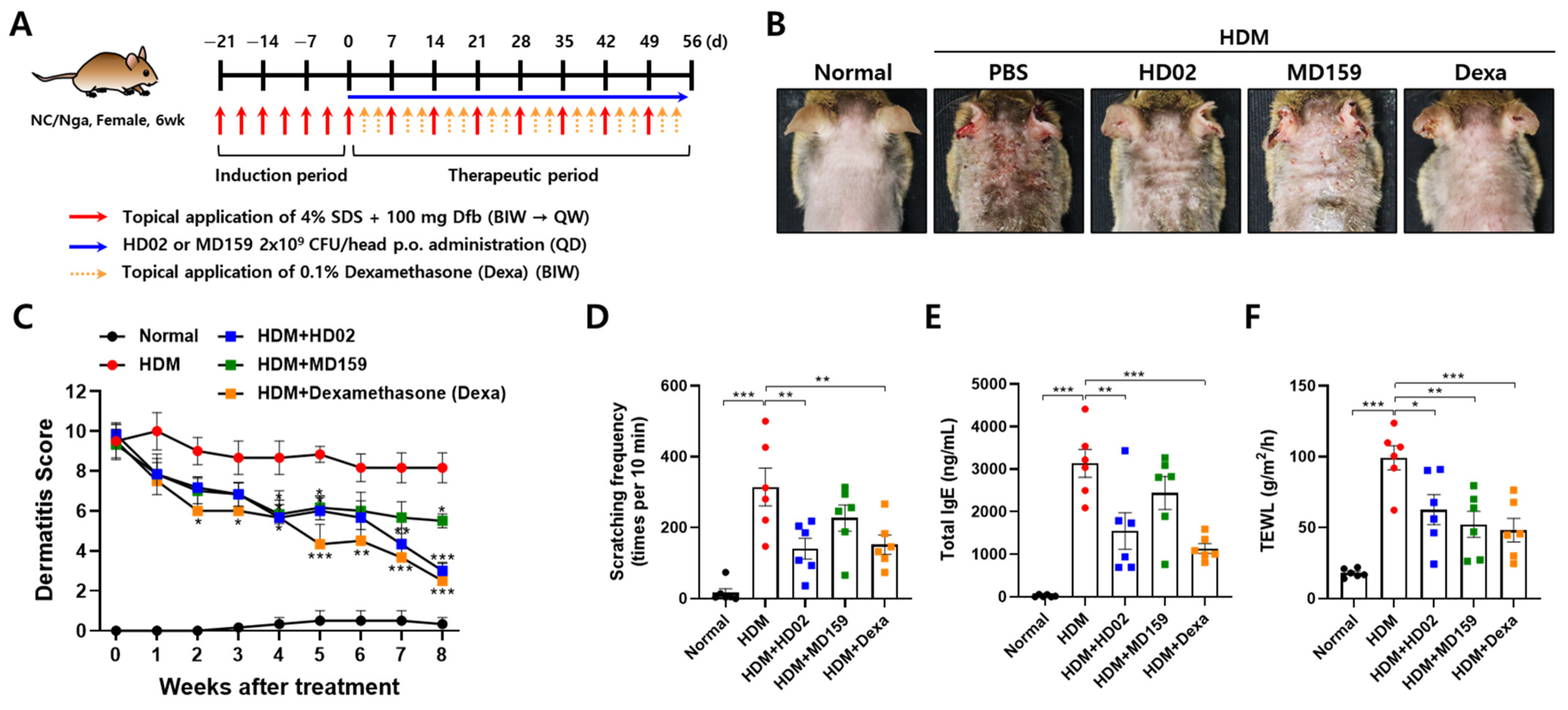
2.6. House Dust Mite (HDM)-Induced AD Model
2.7. RNA Extraction and Quantitative RT-PCR (qRT-PCR) Analysis
2.8. Enzyme-Linked Immunosorbent Assay (ELISA)
2.9. Flow Cytometry Analysis
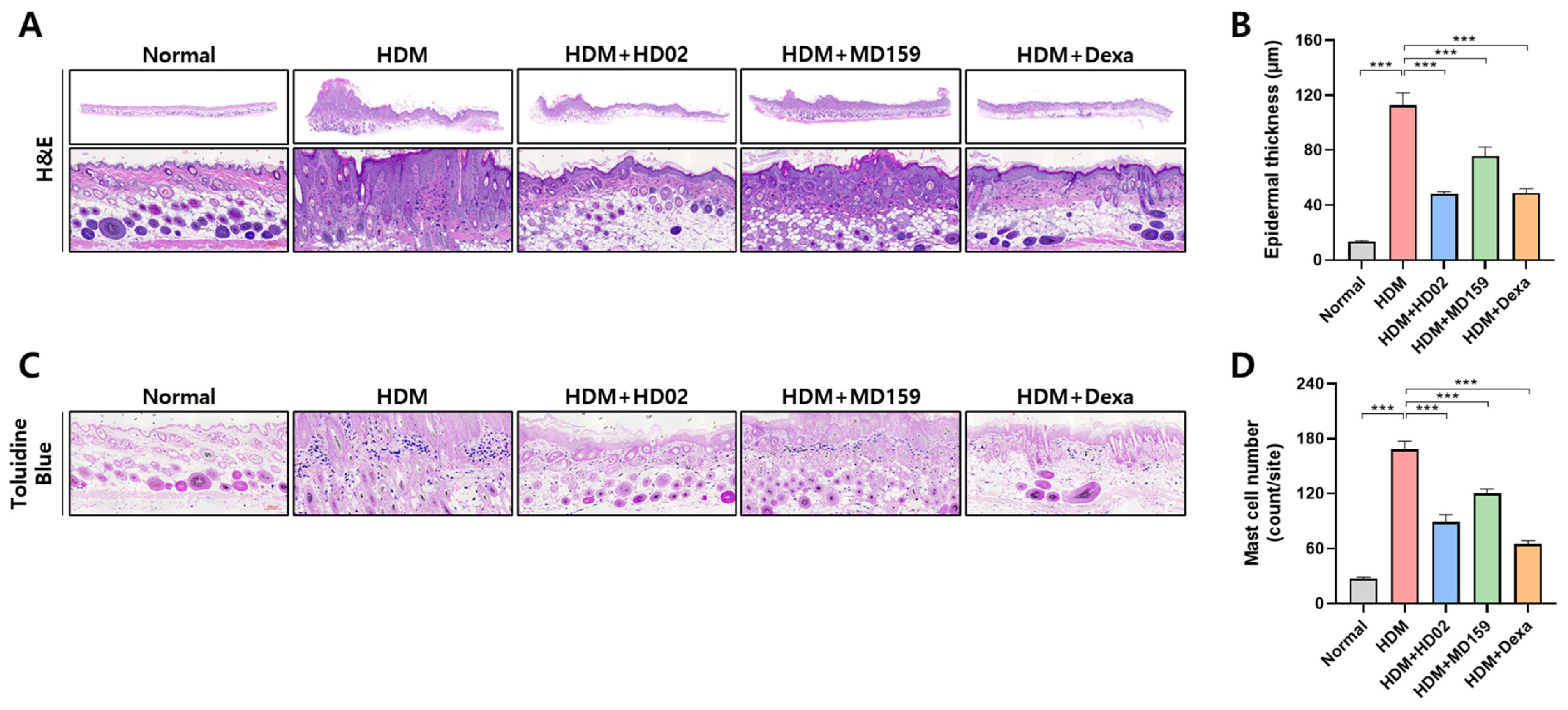
2.10. Histological Analysis
2.11. Whole-Genome Sequencing and Analysis
2.12. Statistical Analysis
3. Results
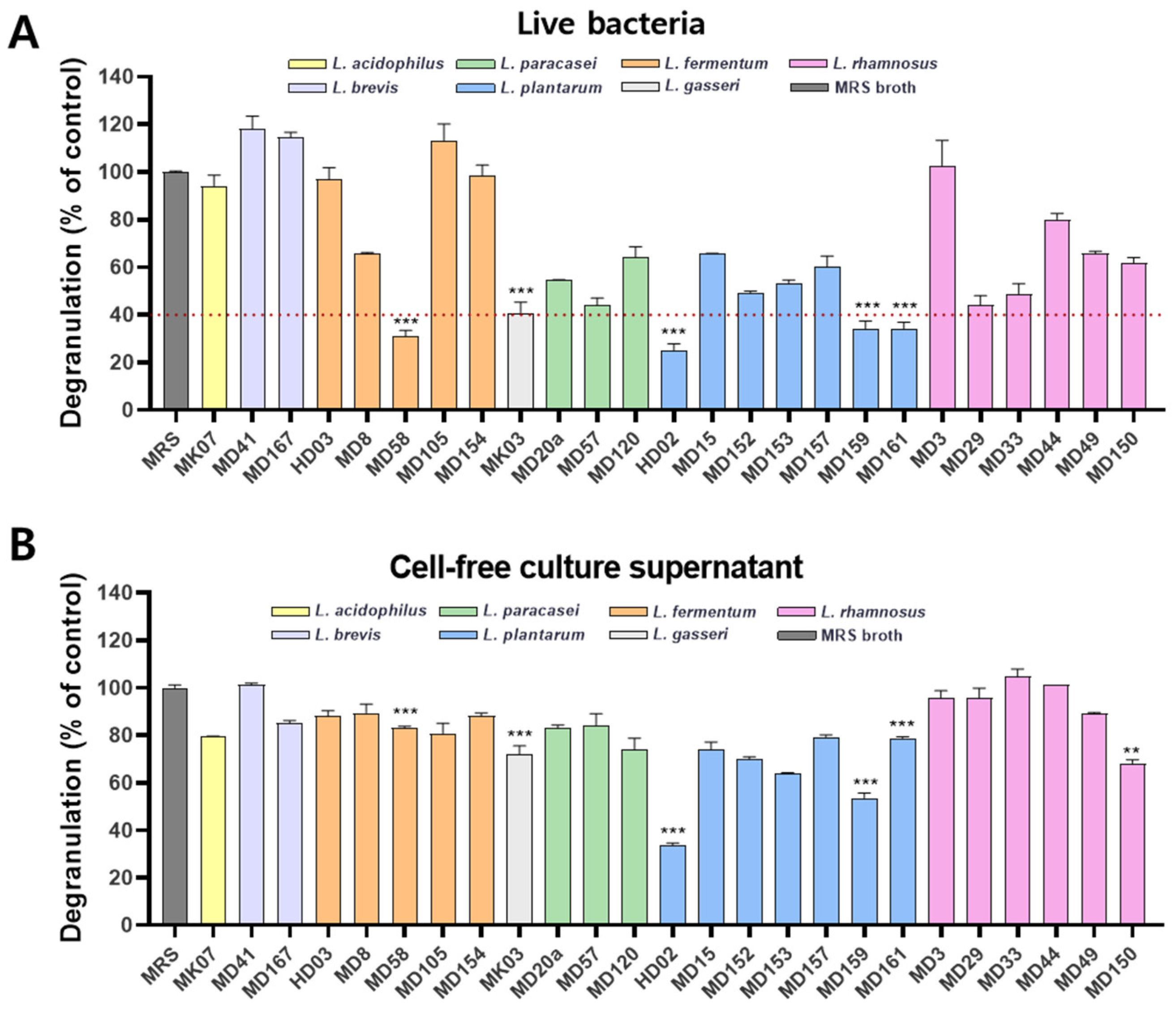
3.1. Identification of L. plantarum HD02 and MD159 as Potent Inhibitors of Mast Cell Degranulation
3.2. Whole-Genome Analysis of L. plantarum HD02 and MD159
3.3. L. plantarum HD02 and MD159 Show Preventive Effects in an MC903-Induced AD Mouse Model
3.4. L. plantarum HD02 and MD159 Show Therapeutic Activity in an HDM-Induced AD Mouse Model
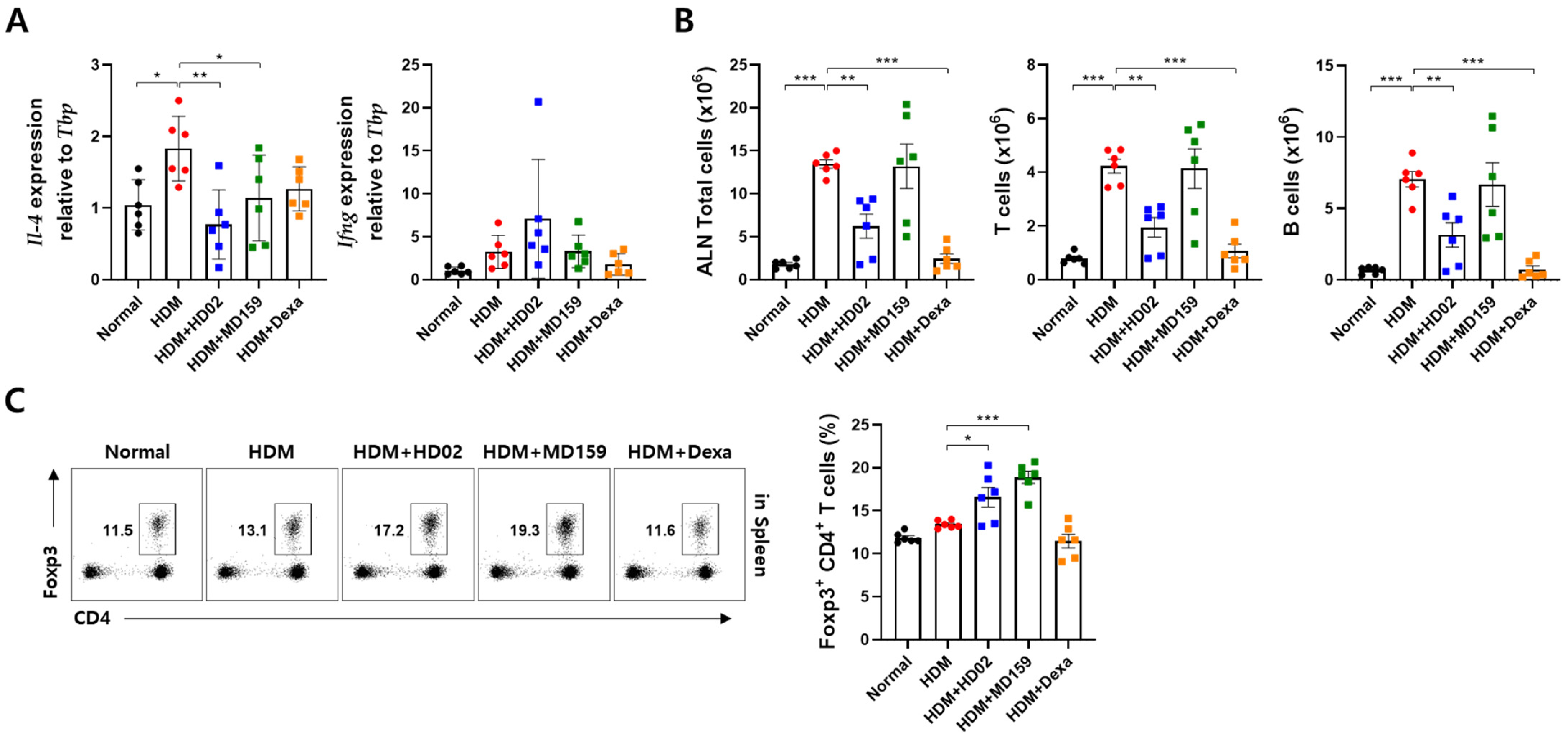
3.5. L. plantarum HD02 and MD159 Regulate Immune Responses in an HDM-Induced AD Mouse Model
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, B.G.; Kim, A.R.; Lee, D.; An, S.B.; Shim, Y.A.; Jang, M.H. Degranulation of Mast Cells as a Target for Drug Development. Cells 2023, 12, 1506. [Google Scholar] [CrossRef] [PubMed]
- Kolkhir, P.; Elieh-Ali-Komi, D.; Metz, M.; Siebenhaar, F.; Maurer, M. Understanding human mast cells: Lesson from therapies for allergic and non-allergic diseases. Nat. Rev. Immunol. 2022, 22, 294–308. [Google Scholar] [CrossRef] [PubMed]
- XOLAIR® (Omalizumab) Injection, for Subcutaneous Use. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2024/103976s5245lbl.pdf (accessed on 15 July 2024).
- Wang, H.H.; Li, Y.C.; Huang, Y.C. Efficacy of omalizumab in patients with atopic dermatitis: A systematic review and meta-analysis. J. Allergy Clin. Immunol. 2016, 138, 1719–1722.e1. [Google Scholar] [CrossRef] [PubMed]
- An, S.B.; Yang, B.G.; Jang, G.; Kim, D.Y.; Kim, J.; Oh, S.M.; Oh, N.; Lee, S.; Moon, J.Y.; Kim, J.A.; et al. Combined IgE neutralization and Bifidobacterium longum supplementation reduces the allergic response in models of food allergy. Nat. Commun. 2022, 13, 5669. [Google Scholar] [CrossRef] [PubMed]
- Ansaldo, E.; Farley, T.K.; Belkaid, Y. Control of immunity by the microbiota. Annu. Rev. Immunol. 2021, 39, 449–479. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. 2017, 66, 515–522. [Google Scholar] [CrossRef]
- Akagawa, S.; Kaneko, K. Gut microbiota and allergic diseases in children. Allergol. Int. 2022, 71, 301–309. [Google Scholar] [CrossRef]
- Cahenzli, J.; Koller, Y.; Wyss, M.; Geuking, M.B.; McCoy, K.D. Intestinal microbial diversity during early-life colonization shapes long-term IgE levels. Cell Host Microbe 2013, 14, 559–570. [Google Scholar] [CrossRef]
- Stefka, A.T.; Feehley, T.; Tripathi, P.; Qiu, J.; McCoy, K.; Mazmanian, S.K.; Tjota, M.Y.; Seo, G.Y.; Cao, S.; Theriault, B.R.; et al. Commensal bacteria protect against food allergen sensitization. Proc. Natl. Acad. Sci. USA 2014, 111, 13145–13150. [Google Scholar] [CrossRef]
- Kozakova, H.; Schwarzer, M.; Tuckova, L.; Srutkova, D.; Czarnowska, E.; Rosiak, I.; Hudcovic, T.; Schabussova, I.; Hermanova, P.; Zakostelska, Z.; et al. Colonization of germ-free mice with a mixture of three lactobacillus strains enhances the integrity of gut mucosa and ameliorates allergic sensitization. Cell Mol. Immunol. 2016, 13, 251–262. [Google Scholar] [CrossRef]
- Hajavi, J.; Esmaeili, S.A.; Varasteh, A.R.; Vazini, H.; Atabati, H.; Mardani, F.; Momtazi-Borojeni, A.A.; Hashemi, M.; Sankian, M.; Sahebkar, A. The immunomodulatory role of probiotics in allergy therapy. J. Cell Physiol. 2019, 234, 2386–2398. [Google Scholar] [CrossRef]
- Kim, J.H.; Jeun, E.J.; Hong, C.P.; Kim, S.H.; Jang, M.S.; Lee, E.J.; Moon, S.J.; Yun, C.H.; Im, S.H.; Jeong, S.G.; et al. Extracellular vesicle-derived protein from Bifidobacterium longum alleviates food allergy through mast cell suppression. J. Allergy Clin. Immunol. 2016, 137, 507–516.e8. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.E.; Bunyavanich, S. Role of the Microbiome in Food Allergy. Curr. Allergy Asthma Rep. 2018, 18, 27. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Zhang, J.; Wang, X.; Jin, Z.; Zhang, P.; Su, H.; Sun, X. Effect of Probiotics on Respiratory Tract Allergic Disease and Gut Microbiota. Front. Nutr. 2022, 9, 821900. [Google Scholar] [CrossRef]
- Hrestak, D.; Matijasic, M.; Cipcic Paljetak, H.; Ledic Drvar, D.; Ljubojevic Hadzavdic, S.; Peric, M. Skin Microbiota in Atopic Dermatitis. Int. J. Mol. Sci. 2022, 23, 3503. [Google Scholar] [CrossRef]
- Losol, P.; Wolska, M.; Wypych, T.P.; Yao, L.; O’Mahony, L.; Sokolowska, M. A cross talk between microbial metabolites and host immunity: Its relevance for allergic diseases. Clin. Transl. Allergy 2024, 14, e12339. [Google Scholar] [CrossRef]
- Hirata, S.I.; Kunisawa, J. Gut microbiome, metabolome, and allergic diseases. Allergol. Int. 2017, 66, 523–528. [Google Scholar] [CrossRef]
- Jeon, H.J.; Ban, O.H.; Bang, W.Y.; Yang, J.; Jung, Y.H. Trends, Functionalities, and Prospects of Probiotics. Microbiol. Biotechnol. Lett. 2022, 50, 465–476. [Google Scholar] [CrossRef]
- Kim, J.H.; Kim, A.R.; Kim, H.S.; Kim, H.W.; Park, Y.H.; You, J.S.; Park, Y.M.; Her, E.; Kim, H.S.; Kim, Y.M.; et al. Rhamnus davurica leaf extract inhibits Fyn activation by antigen in mast cells for anti-allergic activity. BMC Complement. Altern. Med. 2015, 15, 80. [Google Scholar] [CrossRef]
- Moosbrugger-Martinz, V.; Schmuth, M.; Dubrac, S. A Mouse Model for Atopic Dermatitis Using Topical Application of Vitamin D3 or of Its Analog MC903. Methods Mol. Biol. 2017, 1559, 91–106. [Google Scholar] [CrossRef]
- Yamamoto, M.; Haruna, T.; Yasui, K.; Takahashi, H.; Iduhara, M.; Takaki, S.; Deguchi, M.; Arimura, A. A novel atopic dermatitis model induced by topical application with dermatophagoides farinae extract in NC/Nga mice. Allergol. Int. 2007, 56, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Chin, C.S.; Alexander, D.H.; Marks, P.; Klammer, A.A.; Drake, J.; Heiner, C.; Clum, A.; Copeland, A.; Huddleston, J.; Eichler, E.E.; et al. Nonhybrid, finished microbial genome assemblies from long-read SMRT sequencing data. Nat. Methods 2013, 10, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Walker, B.J.; Abeel, T.; Shea, T.; Priest, M.; Abouelliel, A.; Sakthikumar, S.; Cuomo, C.A.; Zeng, Q.; Wortman, J.; Young, S.K.; et al. Pilon: An integrated tool for comprehensive microbial variant detection and genome assembly improvement. PLoS ONE 2014, 9, e112963. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Apweiler, R.; Bairoch, A.; Natale, D.A.; Barker, W.C.; Boeckmann, B.; Ferro, S.; Gasteiger, E.; Huang, H.; Lopez, R.; et al. The Universal Protein Resource (UniProt): An expanding universe of protein information. Nucleic Acids Res. 2006, 34, D187–D191. [Google Scholar] [CrossRef]
- Jain, C.; Rodriguez, R.L.; Phillippy, A.M.; Konstantinidis, K.T.; Aluru, S. High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 2018, 9, 5114. [Google Scholar] [CrossRef]
- Liu, B.; Zheng, D.; Jin, Q.; Chen, L.; Yang, J. VFDB 2019: A comparative pathogenomic platform with an interactive web interface. Nucleic Acids Res. 2019, 47, D687–D692. [Google Scholar] [CrossRef]
- Lebman, D.A.; Coffman, R.L. Interleukin 4 causes isotype switching to IgE in T cell-stimulated clonal B cell cultures. J. Exp. Med. 1988, 168, 853–862. [Google Scholar] [CrossRef]
- Burton, O.T.; Darling, A.R.; Zhou, J.S.; Noval-Rivas, M.; Jones, T.G.; Gurish, M.F.; Chatila, T.A.; Oettgen, H.C. Direct effects of IL-4 on mast cells drive their intestinal expansion and increase susceptibility to anaphylaxis in a murine model of food allergy. Mucosal Immunol. 2013, 6, 740–750. [Google Scholar] [CrossRef]
- Sehra, S.; Yao, Y.; Howell, M.D.; Nguyen, E.T.; Kansas, G.S.; Leung, D.Y.; Travers, J.B.; Kaplan, M.H. IL-4 regulates skin homeostasis and the predisposition toward allergic skin inflammation. J. Immunol. 2010, 184, 3186–3190. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, A.-R.; Jeon, S.-G.; Kim, H.-R.; Hong, H.; Yoon, Y.W.; Lee, B.-M.; Yoon, C.H.; Choi, S.J.; Jang, M.H.; Yang, B.-G. Preventive and Therapeutic Effects of Lactiplantibacillus plantarum HD02 and MD159 through Mast Cell Degranulation Inhibition in Mouse Models of Atopic Dermatitis. Nutrients 2024, 16, 3021. https://doi.org/10.3390/nu16173021
Kim A-R, Jeon S-G, Kim H-R, Hong H, Yoon YW, Lee B-M, Yoon CH, Choi SJ, Jang MH, Yang B-G. Preventive and Therapeutic Effects of Lactiplantibacillus plantarum HD02 and MD159 through Mast Cell Degranulation Inhibition in Mouse Models of Atopic Dermatitis. Nutrients. 2024; 16(17):3021. https://doi.org/10.3390/nu16173021
Chicago/Turabian StyleKim, A-Ram, Seong-Gak Jeon, Hyung-Ran Kim, Heeji Hong, Yong Won Yoon, Byung-Min Lee, Chung Hoo Yoon, Soo Jin Choi, Myoung Ho Jang, and Bo-Gie Yang. 2024. "Preventive and Therapeutic Effects of Lactiplantibacillus plantarum HD02 and MD159 through Mast Cell Degranulation Inhibition in Mouse Models of Atopic Dermatitis" Nutrients 16, no. 17: 3021. https://doi.org/10.3390/nu16173021
APA StyleKim, A.-R., Jeon, S.-G., Kim, H.-R., Hong, H., Yoon, Y. W., Lee, B.-M., Yoon, C. H., Choi, S. J., Jang, M. H., & Yang, B.-G. (2024). Preventive and Therapeutic Effects of Lactiplantibacillus plantarum HD02 and MD159 through Mast Cell Degranulation Inhibition in Mouse Models of Atopic Dermatitis. Nutrients, 16(17), 3021. https://doi.org/10.3390/nu16173021