
Rosehip Extract Decreases Reactive Oxygen Species Production and Lipid Accumulation in Hypertrophic 3T3-L1 Adipocytes with the Modulation of Inflammatory State



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Rosehip Extract
2.2. Adipocyte Differentiation and Experiment Procedure
2.3. Intracellular Reactive Oxygen Species Determination
2.4. Lipid Content Quantification
2.5. Quantitative Real-Time PCR Analysis
2.6. Quantification of Adipokines
2.7. Statistical Data Analysis
3. Results and Discussion
3.1. The Effect of Rosehip Extract on Lipid Accumulation and Lipogenic Genes Expression
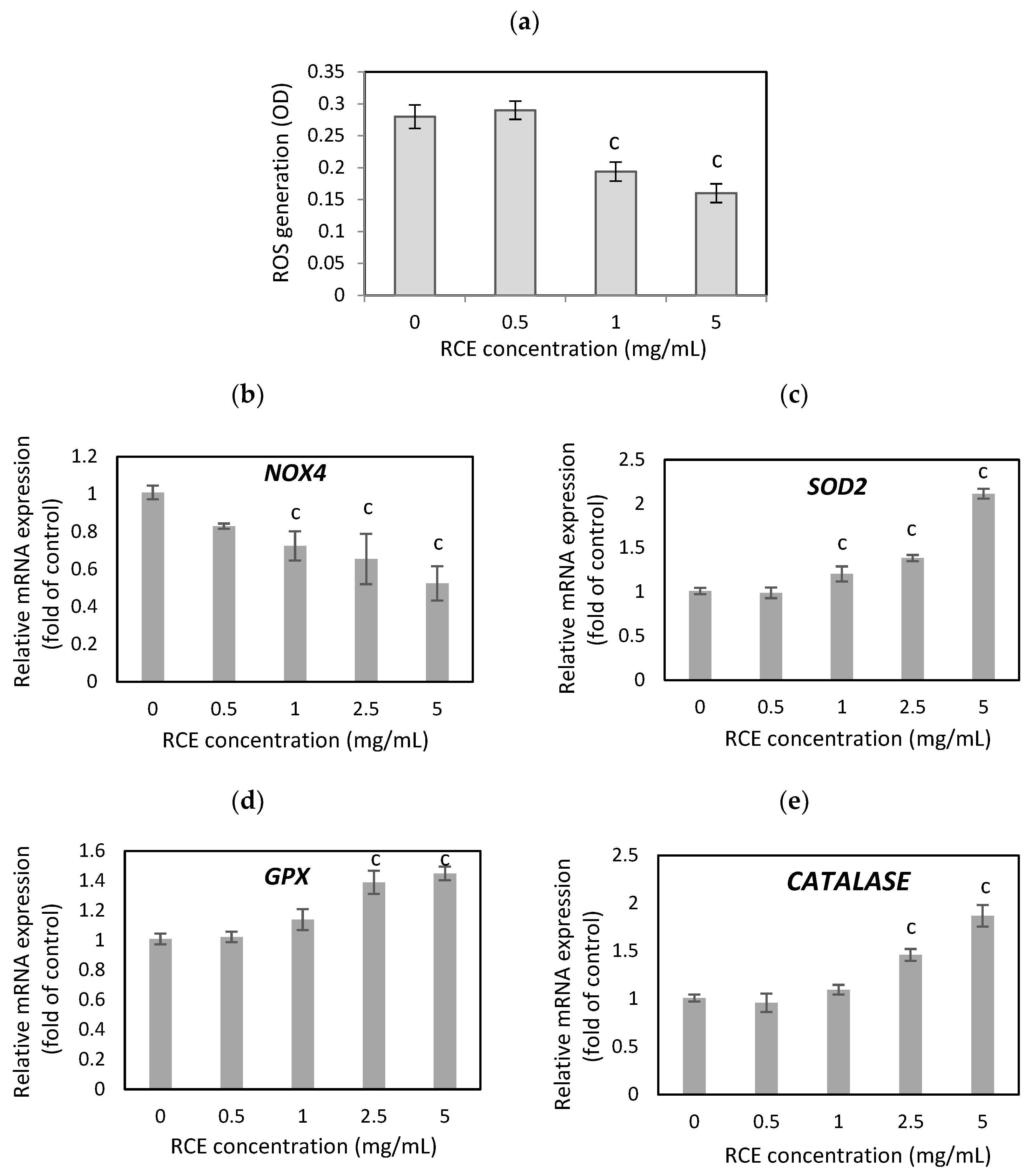
3.2. The Effect of Rosehip Extract on Oxidative Stress
3.3. The Effect of Rosehip Extract on Adipokines Expression and Secretion
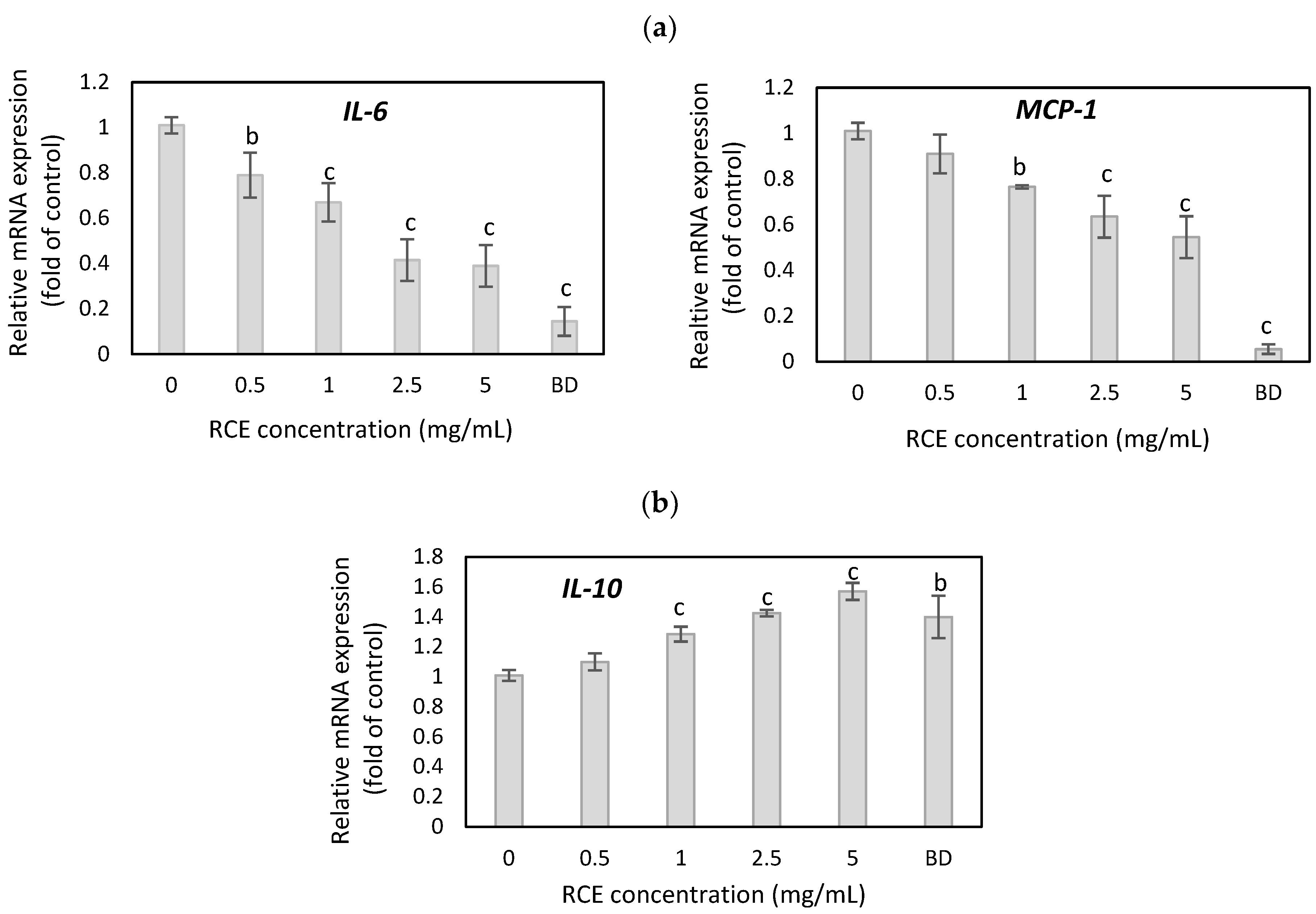
3.4. The Effect of Rosehip Extract on Inflammatory Cytokine Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 4, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Qiu, T.; Li, L.; Yu, R.; Chen, X.; Li, C.; Proud, C.G.; Jiang, T. Pathophysiology of obesity and its associated diseases. Acta Pharm. Sin. B 2023, 6, 2403–2424. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.Z.; Pedersen, L.; Halberg, N. Cellular mechanisms linking cancers to obesity. Cell Stress 2021, 5, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Haider, N.; Larose, L. Harnessing adipogenesis to prevent obesity. Adipocyte 2019, 1, 98–104. [Google Scholar] [CrossRef]
- García-Barrado, M.J.; Iglesias-Osma, M.C.; Pérez-García, E.; Carrero, S.; Blanco, E.J.; Carretero-Hernández, M.; Carretero, J. Role of Flavonoids in The Interactions among Obesity, Inflammation, and Autophagy. Pharmaceuticals 2020, 11, 342. [Google Scholar] [CrossRef]
- Khalilpourfarshbafi, M.; Gholami, K.; Murugan, D.D.; Abdul Sattar, M.Z.; Abdullah, N.A. Differential effects of dietary flavonoids on adipogenesis. Eur. J. Nutr. 2019, 58, 5–25. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Liu, X.; Li, J.; Zhang, J.; Liu, D. Chemical constituents and pharmacological activities of medicinal plants from Rosa genus. Chin. Herb. Med. 2022, 14, 187–209. [Google Scholar] [CrossRef]
- Patel, S. Rose hip as an underutilized functional food: Evidence-based review. Trends Food Sci. 2017, 63, 29–38. [Google Scholar] [CrossRef]
- Fetni, S.; Bertella, N.; Ouahab, A. LC-DAD/ESI-MS/MS characterization of phenolic constituents in Rosa canina L. and its protective effect in cells. Biomed. Chromatogr. 2020, 34, e4961. [Google Scholar] [CrossRef]
- Montazeri, N.; Baher, E.; Mirzajani, F.; Barami, Z.; Yousefian, S. Phytochemical contents and biological activities of Rosa canina fruit from Iran. J. Med. Plants Res. 2011, 5, 4584–4589. [Google Scholar]
- Peña, F.; Valencia, S.; Tereucán, G.; Nahuelcura, J.; Jiménez-Aspee, F.; Cornejo, P.; Ruiz, A. Bioactive Compounds and Antioxidant Activity in the Fruit of Rosehip (Rosa canina L. and Rosa rubiginosa L.). Molecules 2023, 28, 3544. [Google Scholar] [CrossRef]
- Mourabit, Y.; El Hajjaji, S.; Taha, D.; Badaoui, B.; El Yadini, M.; Rusu, M.E.; Lee, L.H.; Bouyahya, A.; Bourais, I. HPLC-DAD-ESI/MS phytochemical investigation, antioxidant, and antidiabetic activities of Moroccan Rosa canina L. extracts, Biocatal. Agric. Biotechnol. 2023, 52, 102817. [Google Scholar] [CrossRef]
- Ilyasoglu, H. Characterization of rosehip (Rosa canina L.) seed and seed oil. Int. J. Food Prop. 2014, 17, 1591–1598. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Rychlik, J.; Grajek, W. Cranberries (Oxycoccus quadripetalus) inhibit adipogenesis and lipogenesis in 3T3-L1 cells. Food Chem. 2014, 148, 246–252. [Google Scholar] [CrossRef]
- Kowalska, K.; Olejnik, A.; Zielińska, J.; Olkowicz, M. Inhibitory effects of lingonberry (Vaccinium vitis-idaea L.) fruit extract on obesity-induced inflammation in 3T3-L1 adipocytes and RAW 264.7 macrophages. J. Funct. Foods. 2019, 54, 371–380. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, J.W.; Cha, Y.N.; Kim, C. A quantitative nitroblue tetrazolium assay for determining intracellular superoxide anion production in phagocytic cells. J. Immunoass. Immunochem. 2006, 27, 31–44. [Google Scholar] [CrossRef]
- Laforest, S.; Labrecque, J.; Michaud, A.; Cianflone, K.; Tchernof, A. Adipocyte size as a determinant of metabolic disease and adipose tissue dysfunction. Crit. Rev. Clin. Lab. Sci. 2015, 52, 301–313. [Google Scholar] [CrossRef]
- Moseti, D.; Regassa, A.; Kim, W.K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Kim, J.B.; Spiegelman, B.M. ADD1/SREBP1 promotes adipocyte differentiation and gene expression linked to fatty acid metabolism. Genes Dev. 1996, 10, 1096–1107. [Google Scholar] [CrossRef]
- Loftus, T.M.; Jaworsky, D.E.; Frehywot, G.L.; Townsend, C.A.; Ronnett, G.V.; Lane, M.D.; Kuhajda, F.P. Reduced food intake and body weight in mice treated with fatty acid synthase inhibitors. Science 2000, 288, 2379–2381. [Google Scholar] [CrossRef]
- Josan, C.; Kakar, S.; Raha, S. Matrigel® enhances 3T3-L1 cell differentiation. Adipocyte 2021, 10, 361–377. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, A.; Nishida, N.; Matsuura, Y.; Shibata, N. Rosehip Extract Inhibits Lipid Accumulation in White Adipose Tissue by Suppressing the Expression of Peroxisome Proliferator-activated Receptor Gamma. Prev. Nutr. Food Sci. 2013, 18, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Sanz-Lamora, H.; Nicola-Llorente, M.; Torres-Oteros, D.; Pérez-Martí, A.; Aghziel, I.; Lozano-Castellón, J.; Vallverdú-Queralt, A.; Canudas, S.; Marrero, P.F.; Haro, D.; et al. The Antiobesity Effects of Rosehip (Rosa canina) Flesh by Antagonizing the PPAR Gamma Activity in High-Fat Diet-Fed Mice. Mol. Nutr. Food Res. 2024, 68, 2300539. [Google Scholar] [CrossRef] [PubMed]
- Hashem, D.F.; Abdollahifard, M.; Khalighi, S.F.; Taghavi, S.M.; Shojaee, A.; Sabet, Z. Effects of Rosa canina L. fruit on glycemia and lipid profile in type 2 diabetic patients: A randomized, double-blind, placebo-controlled clinical trial. J. Med. Plants 2015, 14, 95–104. [Google Scholar]
- Nagatomo, A.; Nishida, N.; Fukuhara, I.; Noro, A.; Kozai, Y.; Sato, H.; Matsuura, Y. Daily intake of rose hip extract decreases abdominal visceral fat in preobese subjects: A randomized, double-blind, placebo-controlled clinical trial. Diabetes Metab. Syndr. Obes. 2015, 8, 147–156. [Google Scholar] [CrossRef]
- Ma, Q.; Cui, Y.; Xu, S.; Zhao, Y.; Yuan, H.; Piao, G. Synergistic Inhibitory Effects of Acacetin and 11 Other Flavonoids Isolated from Artemisia sacrorum on Lipid Accumulation in 3T3-L1 Cells. J. Agric. Food Chem. 2018, 66, 12931–12940. [Google Scholar] [CrossRef]
- Marseglia, L.; Manti, S.; D’Angelo, G.; Nicotera, A.; Parisi, E.; Di Rosa, G.; Gitto, E.; Arrigo, T. Oxidative stress in obesity: A critical component in human diseases. Int. J. Mol. Sci. 2014, 16, 378–400. [Google Scholar] [CrossRef]
- Kanda, Y.; Hinata, T.; Kang, S.W.; Watanabe, Y. Reactive oxygen species mediate adipocyte differentiation in mesenchymal stem cells. Life Sci. 2011, 89, 250–258. [Google Scholar] [CrossRef]
- Hua, H.; Wu, M.; Wu, T.; Ji, Y.; Jin, L.; Du, Y.; Zhang, Y.; Huang, S.; Zhang, A.; Ding, G.; et al. Reduction of NADPH oxidase 4 in adipocytes contributes to the anti-obesity effect of dihydroartemisinin. Heliyon 2023, 9, 14028. [Google Scholar] [CrossRef]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Matoušková, P.; Hanousková, B.; Skálová, L. MicroRNAs as Potential Regulators of Glutathione Peroxidases Expression and Their Role in Obesity and Related Pathologies. Int. J. Mol. Sci. 2018, 19, 1199. [Google Scholar] [CrossRef] [PubMed]
- Forouzanfar, F.; Tabatabaei, Z.; Emami, S.A.; Ayati, Z.; Tayarani-Najaran, Z. Protective effects of fruit extract of Rosa canina and quercetin on human umbilical vein endothelial cell injury induced by hydrogen peroxide. Food Sci. Nutr. 2023, 11, 7618–7625. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, H.; Hosseinzadeh, S.; Akbartabar Touri, M.; Ghavamzadeh, M.; Jafari Barmak, M.; Sayahi, M.; Sadeghi, H. Hepatoprotective effect of Rosa canina fruit extract against carbon tetrachloride induced hepatotoxicity in rat. Avicenna J. Phytomed. 2016, 6, 181–188. [Google Scholar]
- Sadeghi, H.; Karimizadeh, E.; Sadeghi, H.; Panahi Kokhdan, E.; Mansourian, M.; Abbaszadeh-Goudarzi, K.; Shokripour, M.; Asfaram, A.; Doustimotlagh, A.H. Protective Effects of Hydroalcoholic Extract of Rosa canina Fruit on Vancomycin-Induced Nephrotoxicity in Rats. J. Toxicol. 2021, 2021, 5525714. [Google Scholar] [CrossRef]
- Erfani, M.; Ghazi Tabatabaei, Z.; Sadigh-Eteghad, S.; Farokhi-Sisakht, F.; Farajdokht, F.; Mahmoudi, J.; Karimi, P.; Nasrolahi, A. Rosa canina L. methanolic extract prevents heat stress-induced memory dysfunction in rats. Exp. Physiol. 2019, 104, 1544–1554. [Google Scholar] [CrossRef] [PubMed]
- Chrubasik, C.; Roufogalis, B.D.; Müller-Ladner, U.; Chrubasik, S. A systematic review on the Rosa canina effect and efficacy profiles. Phytother. Res. 2008, 22, 725–733. [Google Scholar] [CrossRef]
- Ayati, Z.; Amiri, M.S.; Ramezani, M.; Delshad, E.; Sahebkar, A.; Emami, S.A. Phytochemistry, Traditional Uses and Pharmacological Profile of Rose Hip: A Review. Curr. Pharm. Des. 2018, 24, 4101–4124. [Google Scholar] [CrossRef]
- Su, H.M.; Feng, L.N.; Zheng, X.D.; Chen, W. Myricetin protects against diet-induced obesity and ameliorates oxidative stress in C57BL/6 mice. J. Zhejiang Univ. Sci. B 2016, 17, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Preston, K.J.; Scalia, R.G.; Autieri, M.V. Adipocyte Phenotype Flexibility and Lipid Dysregulation. Cells 2022, 11, 882. [Google Scholar] [CrossRef]
- Zelissen, P.M.J.; Stenlof, K.; Lean, M.E.J.; Fogteloo, J.; Keulen, E.T.P.; Wilding, J.; Finer, N.; Rössner, S.; Lawrence, E.; Fletcher, C.; et al. Effect of three treatment schedules of recombinant methionyl human leptin on body weight in obese adults: A randomized, placebo-controlled trial. Diabetes Obes. Metab. 2005, 7, 755–761. [Google Scholar] [CrossRef]
- Aprahamian, T.R. Elevated adiponectin expression promotes adipose tissue vascularity under conditions of diet-induced obesity. Metabolism 2013, 62, 1730–1738. [Google Scholar] [CrossRef] [PubMed]
- Muse, E.D.; Obici, S.; Bhanot, S.; Monia, B.P.; MacKay, R.A.; Rajala, M.W.; Scherer, P.E.; Rossetti, L. Role of resistin in diet-induced hepatic insulin resistance. J. Clin. Investig. 2004, 114, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Lazar, M.A. Resistin- and Obesity-associated metabolic diseases. Horm. Metab. Res. 2007, 39, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Reilly, M.P.; Lehrke, M.; Wolfe, M.L.; Rohatgi, A.; Lazar, M.A.; Rader, D.J. Resistin is an inflammatory marker of atherosclerosis in humans. Circulation 2005, 111, 932–939. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Z.; Zhou, H.H.; Feng, Y.; Bu, Y.; Zhai, D.; Zhang, G.; Ding, S.; Wang, E.; Mi, Y.; et al. Effect of flavonoid intake on circulating levels of adiponectin and leptin: A systematic review and meta-analysis of randomized controlled clinical trials. Phytother. Res. 2022, 36, 4139–4154. [Google Scholar] [CrossRef]
- Mahmoud, A.M. Hematological alterations in diabetic rats—Role of adipocytokines and effect of citrus flavonoids. EXCLI J. 2013, 12, 647–657. [Google Scholar]
- Mohri, S.; Takahashi, H.; Sakai, M.; Waki, N.; Takahashi, S.; Aizawa, K.; Suganuma, H.; Ara, T.; Sugawara, T.; Shibata, D.; et al. Integration of bioassay and non-target metabolite analysis of tomato reveals that β-carotene and lycopene activate the adiponectin signaling pathway, including AMPK phosphorylation. PLoS ONE 2022, 17, 0267248. [Google Scholar] [CrossRef]
- Khorshidi, M.; Moini, A.; Alipoor, E.; Rezvan, N.; Gorgani-Firuzjaee, S.; Yaseri, M.; Hosseinzadeh-Attar, M.J. The effects of quercetin supplementation on metabolic and hormonal parameters as well as plasma concentration and gene expression of resistin in overweight or obese women with polycystic ovary syndrome. Phytother. Res. 2018, 32, 2282–2289. [Google Scholar] [CrossRef]
- Stenkula, K.G.; Erlanson-Albertsson, C. Adipose cell size: Importance in health and disease. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, 284–295. [Google Scholar] [CrossRef]
- Skurk, T.; Alberti-Huber, C.; Herder, C.; Hauner, H. Relationship between adipocyte size and adipokine expression and secretion. J. Clin. Endocrinol. Metab. 2007, 92, 1023–1033. [Google Scholar] [CrossRef]
- Bastard, J.P.; Jardel, C.; Bruckert, E.; Blondy, P.; Capeau, J.; Laville, M.; Vidal, H.; Hainque, B. Elevated levels of interleukin 6 are reduced in serum and subcutaneous adipose tissue of obese women after weight loss. J. Clin. Endocrinol. Metab. 2000, 85, 3338–3342. [Google Scholar] [CrossRef] [PubMed]
- Kanda, H.; Tateya, S.; Tamori, Y.; Kotani, K.; Hiasa, K.; Kitazawa, R.; Kitazawa, S.; Miyachi, H.; Maeda, S.; Egashira, K.; et al. MCP-1 contributes to macrophage infiltration into adipose tissue, insulin resistance, and hepatic steatosis in obesity. J. Clin. Investig. 2006, 116, 1494–1505. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh Pahlavani, H. Exercise Therapy for People with Sarcopenic Obesity: Myokines and Adipokines as Effective Actors. Front. Endocrinol. 2022, 13, 811751. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Xu, D.; Yin, C.; Wang, S.; Wang, M.; Xiao, Y. IL-10/STAT3 is reduced in childhood obesity with hypertriglyceridemia and is related to triglyceride level in diet-induced obese rats. BMC Endocr. Disord. 2018, 18, 39. [Google Scholar] [CrossRef]
- de Farias, T.; Cruz, M.M.; de Sa, R.; Severi, I.; Perugini, J.; Senzacqua, M.; Cerutti, S.M.; Giordano, A.; Cinti, S.; lonso-Vale, M.I.C. Melatonin supplementation decreases hypertrophic obesity and inflammation induced by high-fat diet in mice. Front. Endocrinol. 2019, 10, 750. [Google Scholar] [CrossRef]
- Kawser Hossain, M.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Kumar Saha, S.; Yang, G.M.; Choi, H.Y.; Cho, S.G. Molecular Mechanisms of the Anti-Obesity and Anti-Diabetic Properties of Flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef]
- Schwager, J.; Hoeller, U.; Wolfram, S.; Richard, N. Rose hip and its constituent galactolipids confer cartilage protection by modulating cytokine, and chemokine expression. BMC Complement. Altern. Med. 2011, 11, 105. [Google Scholar] [CrossRef] [PubMed]
- Schwager, J.; Richard, N.; Schoop, R.; Wolfram, S. A novel rose hip preparation with enhanced anti-inflammatory and chondroprotective effects. Mediat. Inflamm. 2014, 2014, 105710. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, K.; Olejnik, A. Rosehip Extract Decreases Reactive Oxygen Species Production and Lipid Accumulation in Hypertrophic 3T3-L1 Adipocytes with the Modulation of Inflammatory State. Nutrients 2024, 16, 3269. https://doi.org/10.3390/nu16193269
Kowalska K, Olejnik A. Rosehip Extract Decreases Reactive Oxygen Species Production and Lipid Accumulation in Hypertrophic 3T3-L1 Adipocytes with the Modulation of Inflammatory State. Nutrients. 2024; 16(19):3269. https://doi.org/10.3390/nu16193269
Chicago/Turabian StyleKowalska, Katarzyna, and Anna Olejnik. 2024. "Rosehip Extract Decreases Reactive Oxygen Species Production and Lipid Accumulation in Hypertrophic 3T3-L1 Adipocytes with the Modulation of Inflammatory State" Nutrients 16, no. 19: 3269. https://doi.org/10.3390/nu16193269
APA StyleKowalska, K., & Olejnik, A. (2024). Rosehip Extract Decreases Reactive Oxygen Species Production and Lipid Accumulation in Hypertrophic 3T3-L1 Adipocytes with the Modulation of Inflammatory State. Nutrients, 16(19), 3269. https://doi.org/10.3390/nu16193269