
Effects of Dietary Quality on Vaginal Microbiome Composition Throughout Pregnancy in a Multi-Ethnic Cohort

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Subjects and Recruitment
2.2. Data Collections
2.2.1. Quantitative Food Frequency Questionnaire
2.2.2. Vaginal Swab Collection
2.2.3. Demographic Data and Pregnancy Outcome Data Extraction
2.3. Microbiome Sequencing
2.4. Data Analysis
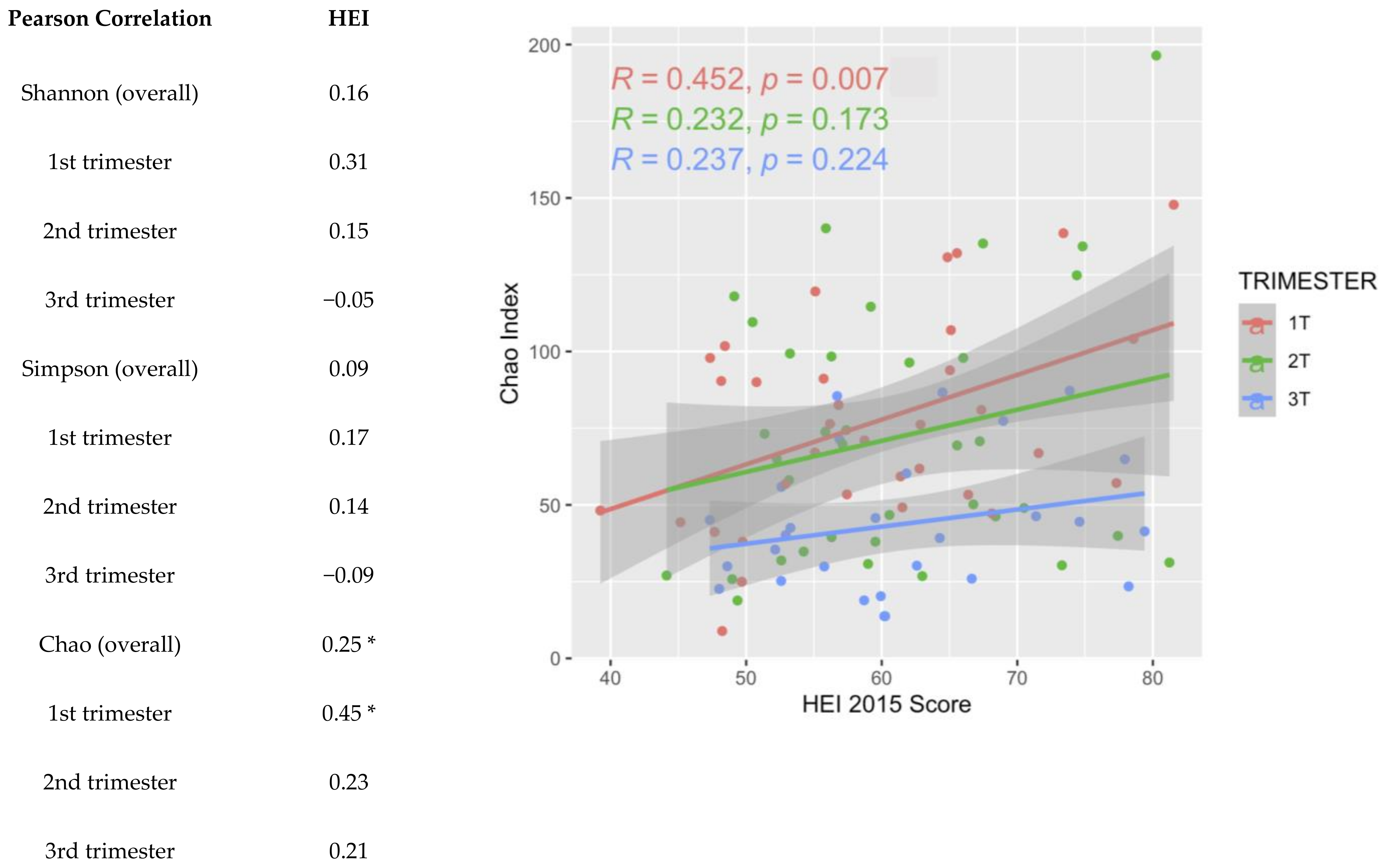
3. Results
3.1. Study Group Demographics
3.2. 16S Metagenomic Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doerflinger, S.Y.; Throop, A.L.; Herbst-Kralovetz, M.M. Bacteria in the vaginal microbiome alter the innate immune response and barrier properties of the human vaginal epithelia in a species-specific manner. J. Infect. Dis. 2014, 209, 1989–1999. [Google Scholar] [CrossRef] [PubMed]
- Spear, G.T.; Gilbert, D.; Landay, A.L.; Zariffard, R.; French, A.L.; Patel, P.; Gillevet, P.M. Pyrosequencing of the genital microbiotas of HIV-seropositive and -seronegative women reveals Lactobacillus iners as the predominant Lactobacillus Species. Appl. Environ. Microbiol. 2011, 77, 378–381. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, J.; Zhou, L.; Shi, H.; Mai, H.; Su, J.; Ma, X.; Zhong, J. The First Lanthipeptide from Lactobacillus iners, Inecin L, Exerts High Antimicrobial Activity against Human Vaginal Pathogens. Appl. Environ. Microbiol. 2023, 89, e0212322. [Google Scholar] [CrossRef] [PubMed]
- Boskey, E.R.; Cone, R.A.; Whaley, K.J.; Moench, T.R. Origins of vaginal acidity: High D/L lactate ratio is consistent with bacteria being the primary source. Hum. Reprod. 2001, 16, 1809–1813. [Google Scholar] [CrossRef]
- Voravuthikunchai, S.P.; Bilasoi, S.; Supamala, O. Antagonistic activity against pathogenic bacteria by human vaginal lactobacilli. Anaerobe 2006, 12, 221–226. [Google Scholar] [CrossRef]
- Brown, R.G.; Al-Memar, M.; Marchesi, J.R.; Lee, Y.S.; Smith, A.; Chan, D.; Lewis, H.; Kindinger, L.; Terzidou, V.; Bourne, T.; et al. Establishment of vaginal microbiota composition in early pregnancy and its association with subsequent preterm prelabor rupture of the fetal membranes. Transl. Res. 2019, 207, 30–43. [Google Scholar] [CrossRef]
- Geldenhuys, J.; Redelinghuys, M.J.; Lombaard, H.A.; Ehlers, M.M.; Cowan, D.; Kock, M.M. Diversity of the gut, vaginal and oral microbiome among pregnant women in South Africa with and without pre-eclampsia. Front. Glob. Womens Health 2022, 3, 810673. [Google Scholar] [CrossRef]
- Lebeer, S.; Ahannach, S.; Gehrmann, T.; Wittouck, S.; Eilers, T.; Oerlemans, E.; Condori, S.; Dillen, J.; Spacova, I.; Vander Donck, L.; et al. A citizen-science-enabled catalogue of the vaginal microbiome and associated factors. Nat. Microbiol. 2023, 8, 2183–2195. [Google Scholar] [CrossRef]
- Song, S.D.; Acharya, K.D.; Zhu, J.E.; Deveney, C.M.; Walther-Antonio, M.R.S.; Tetel, M.J.; Chia, N. Daily Vaginal Microbiota Fluctuations Associated with Natural Hormonal Cycle, Contraceptives, Diet, and Exercise. mSphere 2020, 5, e00593-20. [Google Scholar] [CrossRef] [PubMed]
- Krog, M.C.; Hugerth, L.W.; Fransson, E.; Bashir, Z.; Nyboe Andersen, A.; Edfeldt, G.; Engstrand, L.; Schuppe-Koistinen, I.; Nielsen, H.S. The healthy female microbiome across body sites: Effect of hormonal contraceptives and the menstrual cycle. Hum. Reprod. 2022, 37, 1525–1543. [Google Scholar] [CrossRef]
- Ravel, J.; Gajer, P.; Abdo, Z.; Schneider, G.M.; Koenig, S.S.; McCulle, S.L.; Karlebach, S.; Gorle, R.; Russell, J.; Tacket, C.O.; et al. Vaginal microbiome of reproductive-age women. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4680–4687. [Google Scholar] [CrossRef] [PubMed]
- Aagaard, K.; Riehle, K.; Ma, J.; Segata, N.; Mistretta, T.A.; Coarfa, C.; Raza, S.; Rosenbaum, S.; Van den Veyver, I.; Milosavljevic, A.; et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS ONE 2012, 7, e36466. [Google Scholar] [CrossRef]
- Romero, R.; Hassan, S.S.; Gajer, P.; Tarca, A.L.; Fadrosh, D.W.; Nikita, L.; Galuppi, M.; Lamont, R.F.; Chaemsaithong, P.; Miranda, J.; et al. The composition and stability of the vaginal microbiota of normal pregnant women is different from that of non-pregnant women. Microbiome 2014, 2, 4. [Google Scholar] [CrossRef]
- Mitsou, E.K.; Kakali, A.; Antonopoulou, S.; Mountzouris, K.C.; Yannakoulia, M.; Panagiotakos, D.B.; Kyriacou, A. Adherence to the Mediterranean diet is associated with the gut microbiota pattern and gastrointestinal characteristics in an adult population. Br. J. Nutr. 2017, 117, 1645–1655. [Google Scholar] [CrossRef]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef]
- Garcia-Mantrana, I.; Selma-Royo, M.; Alcantara, C.; Collado, M.C. Shifts on Gut Microbiota Associated to Mediterranean Diet Adherence and Specific Dietary Intakes on General Adult Population. Front. Microbiol. 2018, 9, 890. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.M.; Martin, C.L.; Siega-Riz, A.M.; Dole, N.; Basta, P.V.; Serrano, M.; Fettweis, J.; Wu, M.; Sun, S.; Thorp, J.M., Jr.; et al. Is prenatal diet associated with the composition of the vaginal microbiome? Paediatr. Perinat. Epidemiol. 2022, 36, 243–253. [Google Scholar] [CrossRef]
- Sun, H.; Yamada, P.; Paetow, A.; Chan, M.; Arslan, A.; Landberg, R.; Dominguez-Bello, M.G.; Young, B.K. A randomized controlled trial of the effects of whole grains versus refined grains diets on the microbiome in pregnancy. Sci. Rep. 2022, 12, 7509. [Google Scholar] [CrossRef] [PubMed]
- Whyte, J.J.; Alexenko, A.P.; Davis, A.M.; Ellersieck, M.R.; Fountain, E.D.; Rosenfeld, C.S. Maternal diet composition alters serum steroid and free fatty acid concentrations and vaginal pH in mice. J. Endocrinol. 2007, 192, 75–81. [Google Scholar] [CrossRef]
- Spear, G.T.; French, A.L.; Gilbert, D.; Zariffard, M.R.; Mirmonsef, P.; Sullivan, T.H.; Spear, W.W.; Landay, A.; Micci, S.; Lee, B.H.; et al. Human alpha-amylase present in lower-genital-tract mucosal fluid processes glycogen to support vaginal colonization by Lactobacillus. J. Infect. Dis. 2014, 210, 1019–1028. [Google Scholar] [CrossRef]
- Dall’Asta, M.; Laghi, L.; Morselli, S.; Re, M.C.; Zagonari, S.; Patuelli, G.; Foschi, C.; Pedna, M.F.; Sambri, V.; Marangoni, A.; et al. Pre-Pregnancy Diet and Vaginal Environment in Caucasian Pregnant Women: An Exploratory Study. Front. Mol. Biosci. 2021, 8, 702370. [Google Scholar] [CrossRef] [PubMed]
- National Cancer Institute; National Institutes of Health. Overview and Background of the Healthy Eating Index. Available online: https://epi.grants.cancer.gov/hei/ (accessed on 15 April 2017).
- Bowyer, R.C.E.; Jackson, M.A.; Pallister, T.; Skinner, J.; Spector, T.D.; Welch, A.A.; Steves, C.J. Use of dietary indices to control for diet in human gut microbiota studies. Microbiome 2018, 6, 77. [Google Scholar] [CrossRef] [PubMed]
- Tsigga, M.; Filis, V.; Hatzopoulou, K.; Kotzamanidis, C.; Grammatikopoulou, M.G. Healthy Eating Index during pregnancy according to pre-gravid and gravid weight status. Public Health Nutr. 2010, 14, 290–296. [Google Scholar] [CrossRef]
- Fettweis, J.M.; Brooks, J.P.; Serrano, M.G.; Sheth, N.U.; Girerd, P.H.; Edwards, D.J.; Strauss, J.F.; The Vaginal Microbiome Consortium; Jefferson, K.K.; Buck, G.A. Differences in vaginal microbiome in African American women versus women of European ancestry. Microbiology 2014, 160, 2272–2282. [Google Scholar] [CrossRef] [PubMed]
- MacIntyre, D.A.; Chandiramani, M.; Lee, Y.S.; Kindinger, L.; Smith, A.; Angelopoulos, N.; Lehne, B.; Arulkumaran, S.; Brown, R.; Teoh, T.G.; et al. The vaginal microbiome during pregnancy and the postpartum period in a European population. Sci. Rep. 2015, 5, 8988. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Yamagishi, Y.; Miyamoto, K.; Oka, K.; Takahashi, M.; Mikamo, H. Characterization of the vaginal microbiota of Japanese women. Anaerobe 2018, 54, 172–177. [Google Scholar] [CrossRef]
- Marangoni, A.; Laghi, L.; Zagonari, S.; Patuelli, G.; Zhu, C.; Foschi, C.; Morselli, S.; Pedna, M.F.; Sambri, V. New Insights into Vaginal Environment During Pregnancy. Front. Mol. Biosci. 2021, 8, 656844. [Google Scholar] [CrossRef]
- Price, J.T.; Vwalika, B.; France, M.; Ravel, J.; Ma, B.; Mwape, H.; Rittenhouse, K.J.; De Paris, K.; Hobbs, M.; Nelson, J.A.; et al. HIV-associated vaginal microbiome and inflammation predict spontaneous preterm birth in Zambia. Sci. Rep. 2022, 12, 8573. [Google Scholar] [CrossRef]
- Stout, M.J.; Zhou, Y.; Wylie, K.M.; Tarr, P.I.; Macones, G.A.; Tuuli, M.G. Early pregnancy vaginal microbiome trends and preterm birth. Am. J. Obs. Gynecol. 2017, 217, 356.e1–356.e18. [Google Scholar] [CrossRef]
- Vinturache, A.E.; Gyamfi-Bannerman, C.; Hwang, J.; Mysorekar, I.U.; Jacobsson, B.; Preterm Birth International, C. Maternal microbiome—A pathway to preterm birth. Semin. Fetal Neonatal Med. 2016, 21, 94–99. [Google Scholar] [CrossRef]
- Strout, N.; Pasic, L.; Hicks, C.; Chua, X.Y.; Tashvighi, N.; Butler, P.; Liu, Z.; El-Assaad, F.; Holmes, E.; Susic, D.; et al. The MothersBabies Study, an Australian Prospective Cohort Study Analyzing the Microbiome in the Preconception and Perinatal Period to Determine Risk of Adverse Pregnancy, Postpartum, and Child-Related Health Outcomes: Study Protocol. Int. J. Environ. Res. Public Health 2023, 20, 6736. [Google Scholar] [CrossRef] [PubMed]
- Alves-Santos, N.H.; Cocate, P.G.; Eshriqui, I.; Benaim, C.; Barros, E.G.; Emmett, P.M.; Kac, G. Dietary patterns and their association with adiponectin and leptin concentrations throughout pregnancy: A prospective cohort. Br. J. Nutr. 2018, 119, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Miller, C.B.; Benny, P.; Riel, J.; Boushey, C.; Perez, R.; Khadka, V.; Qin, Y.; Maunakea, A.K.; Lee, M.J. Adherence to Mediterranean diet impacts gastrointestinal microbial diversity throughout pregnancy. BMC Pregnancy Childbirth 2021, 21, 558. [Google Scholar] [CrossRef] [PubMed]
- Wilkens, L.R.; Kang, M.; Park, S.Y.; Sawyer, A.; Boushey, C.J.; Au, D.L.M.; Paik, H.Y.; Le Marchand, L.; Haiman, C.A.; Kolonel, L.N.; et al. Validation and Comparison of Nutrient Intakes from Two Dietary Questionnaires Developed for the Multiethnic Cohort Study. J. Acad. Nutr. Diet. 2023, 123, 655–663.e651. [Google Scholar] [CrossRef]
- Kolonel, L.N.; Henderson, B.E.; Hankin, J.H.; Nomura, A.M.; Wilkens, L.R.; Pike, M.C.; Stram, D.O.; Monroe, K.R.; Earle, M.E.; Nagamine, F.S. A multiethnic cohort in Hawaii and Los Angeles: Baseline characteristics. Am. J. Epidemiol. 2000, 151, 346–357. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Boushey, C.J.; Wilkens, L.R.; Haiman, C.A.; Le Marchand, L. High-Quality Diets Associate With Reduced Risk of Colorectal Cancer: Analyses of Diet Quality Indexes in the Multiethnic Cohort. Gastroenterology 2017, 153, 386–394.e2. [Google Scholar] [CrossRef]
- Harmon, B.E.; Boushey, C.J.; Shvetsov, Y.B.; Ettienne, R.; Reedy, J.; Wilkens, L.R.; Le Marchand, L.; Henderson, B.E.; Kolonel, L.N. Associations of key diet-quality indexes with mortality in the Multiethnic Cohort: The Dietary Patterns Methods Project. Am. J. Clin. Nutr. 2015, 101, 587–597. [Google Scholar] [CrossRef]
- Stram, D.O.; Hankin, J.H.; Wilkens, L.R.; Pike, M.C.; Monroe, K.R.; Park, S.; Henderson, B.E.; Nomura, A.M.; Earle, M.E.; Nagamine, F.S.; et al. Calibration of the dietary questionnaire for a multiethnic cohort in Hawaii and Los Angeles. Am. J. Epidemiol. 2000, 151, 358–370. [Google Scholar] [CrossRef]
- Krebs-Smith, S.M.; Pannucci, T.E.; Subar, A.F.; Kirkpatrick, S.I.; Lerman, J.L.; Tooze, J.A.; Wilson, M.M.; Reedy, J. Update of the Healthy Eating Index-2015. J. Acad. Nutr. Diet. 2018, 118, 1591–1602. [Google Scholar] [CrossRef]
- Rifas-Shiman, S.L.; Rich-Edwards, J.W.; Kleinman, K.P.; Oken, E.; Gillman, M.W. Dietary quality during pregnancy varies by maternal characteristics in Project Viva: A US cohort. J. Am. Diet. Assoc. 2009, 109, 1004–1011. [Google Scholar] [CrossRef]
- Yee, L.M.; Silver, R.M.; Haas, D.M.; Parry, S.; Mercer, B.M.; Iams, J.; Wing, D.; Parker, C.B.; Reddy, U.M.; Wapner, R.J.; et al. Quality of periconceptional dietary intake and maternal and neonatal outcomes. Am. J. Obs. Gynecol. 2020, 223, 121.e1–121.e8. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, A.L.; Kaar, J.L.; Crume, T.L.; Starling, A.P.; Siega-Riz, A.M.; Ringham, B.M.; Glueck, D.H.; Norris, J.M.; Barbour, L.A.; Friedman, J.E.; et al. Maternal diet quality in pregnancy and neonatal adiposity: The Healthy Start Study. Int. J. Obes. 2016, 40, 1056–1062. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhu, Y.; Chavarro, J.E.; Bao, W.; Tobias, D.K.; Ley, S.H.; Forman, J.P.; Liu, A.; Mills, J.; Bowers, K.; et al. Healthful Dietary Patterns and the Risk of Hypertension Among Women With a History of Gestational Diabetes Mellitus: A Prospective Cohort Study. Hypertension 2016, 67, 1157–1165. [Google Scholar] [CrossRef]
- Institute of Medicine. Weight Gain during Pregnancy: Reexamining the Guidelines; National Academies Press: Washington, DC, USA, 2009. [Google Scholar]
- Balle, C.; Konstantinus, I.N.; Jaumdally, S.Z.; Havyarimana, E.; Lennard, K.; Esra, R.; Barnabas, S.L.; Happel, A.U.; Moodie, Z.; Gill, K.; et al. Hormonal contraception alters vaginal microbiota and cytokines in South African adolescents in a randomized trial. Nat. Commun. 2020, 11, 5578. [Google Scholar] [CrossRef]
- Reid, G. Modulating the vaginal microbiome: The need for a bridge between science and practice. Semin. Reprod. Med. 2014, 32, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Reid, G.; Challis, J.R.G.; Gloor, G.B.; Asztalos, E.; Money, D.; Seney, S.; Bocking, A.D. Effect of Oral Probiotic Lactobacillus rhamnosus GR-1 and Lactobacillus reuteri RC-14 on the Vaginal Microbiota, Cytokines and Chemokines in Pregnant Women. Nutrients 2020, 12, 368. [Google Scholar] [CrossRef]
- Bloom, S.M.; Mafunda, N.A.; Woolston, B.M.; Hayward, M.R.; Frempong, J.F.; Abai, A.B.; Xu, J.; Mitchell, A.J.; Westergaard, X.; Hussain, F.A.; et al. Cysteine dependence of Lactobacillus iners is a potential therapeutic target for vaginal microbiota modulation. Nat. Microbiol. 2022, 7, 434–450. [Google Scholar] [CrossRef]
- Faucher, M.A.; Greathouse, K.L.; Hastings-Tolsma, M. Exploration of the Vaginal and Gut Microbiome, Dietary Intake, and Metabolomics in African American Women by Body Mass Index and Gestational Weight Gain. J. Midwifery Women’s Health 2019, 64, 674–675. [Google Scholar] [CrossRef]
- Sha, B.E.; Zariffard, M.R.; Wang, Q.J.; Chen, H.Y.; Bremer, J.; Cohen, M.H.; Spear, G.T. Female genital-tract HIV load correlates inversely with Lactobacillus species but positively with bacterial vaginosis and Mycoplasma hominis. J. Infect. Dis. 2005, 191, 25–32. [Google Scholar] [CrossRef]
- Chen, S.; Xue, X.; Zhang, Y.; Zhang, H.; Huang, X.; Chen, X.; Deng, G.; Luo, S.; Gao, J. Vaginal Atopobium is Associated with Spontaneous Abortion in the First Trimester: A Prospective Cohort Study in China. Microbiol. Spectr. 2022, 10, e0203921. [Google Scholar] [CrossRef]
- Chen, X.; Lu, Y.; Chen, T.; Li, R. The Female Vaginal Microbiome in Health and Bacterial Vaginosis. Front. Cell. Infect. Microbiol. 2021, 11, 631972. [Google Scholar] [CrossRef] [PubMed]
- Wiesenfeld, H.C.; Hillier, S.L.; Krohn, M.A.; Landers, D.V.; Sweet, R.L. Bacterial vaginosis is a strong predictor of Neisseria gonorrhoeae and Chlamydia trachomatis infection. Clin. Infect. Dis. 2003, 36, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Vitali, B.; Cruciani, F.; Baldassarre, M.E.; Capursi, T.; Spisni, E.; Valerii, M.C.; Candela, M.; Turroni, S.; Brigidi, P. Dietary supplementation with probiotics during late pregnancy: Outcome on vaginal microbiota and cytokine secretion. BMC Microbiol. 2012, 12, 236. [Google Scholar] [CrossRef]
- Rani, V.U.; Rao, S.R.; Grisha, P.; Sharma, A.V.; Usha, M.S. The efficacy of probiotic b. coagulans (snz-1969) tablets in the treatment of recurrent bacterial vaginosis. Int. J. Probiotics Prebiotics 2017, 12, 175–182. [Google Scholar]
- Hantoushzadeh, S.; Golshahi, F.; Javadian, P.; Khazardoost, S.; Aram, S.; Hashemi, S.; Mirarmandehi, B.; Borna, S. Comparative efficacy of probiotic yoghurt and clindamycin in treatment of bacterial vaginosis in pregnant women: A randomized clinical trial. J. Matern. Fetal Neonatal Med. 2012, 25, 1021–1024. [Google Scholar] [CrossRef]
- Laue, C.; Papazova, E.; Liesegang, A.; Pannenbeckers, A.; Arendarski, P.; Linnerth, B.; Domig, K.J.; Kneifel, W.; Petricevic, L.; Schrezenmeir, J. Effect of a yoghurt drink containing Lactobacillus strains on bacterial vaginosis in women—A double-blind, randomised, controlled clinical pilot trial. Benef. Microbes 2018, 9, 35–50. [Google Scholar] [CrossRef]
- Miller, E.A.; Beasley, D.E.; Dunn, R.R.; Archie, E.A. Lactobacilli Dominance and Vaginal pH: Why Is the Human Vaginal Microbiome Unique? Front. Microbiol. 2016, 7, 1936. [Google Scholar] [CrossRef]
- Maskarinec, G.; Hullar, M.A.J.; Monroe, K.R.; Shepherd, J.A.; Hunt, J.; Randolph, T.W.; Wilkens, L.R.; Boushey, C.J.; Le Marchand, L.; Lim, U.; et al. Fecal Microbial Diversity and Structure Are Associated with Diet Quality in the Multiethnic Cohort Adiposity Phenotype Study. J. Nutr. 2019, 149, 1575–1584. [Google Scholar] [CrossRef]
- Noormohammadi, M.; Eslamian, G.; Kazemi, S.N.; Rashidkhani, B. Dietary acid load, alternative healthy eating index score, and bacterial vaginosis: Is there any association? A case-control study. BMC Infect. Dis. 2022, 22, 803. [Google Scholar] [CrossRef]
- Dunlop, A.L.; Satten, G.A.; Hu, Y.J.; Knight, A.K.; Hill, C.C.; Wright, M.L.; Smith, A.K.; Read, T.D.; Pearce, B.D.; Corwin, E.J. Vaginal Microbiome Composition in Early Pregnancy and Risk of Spontaneous Preterm and Early Term Birth Among African American Women. Front. Cell. Infect. Microbiol. 2021, 11, 641005. [Google Scholar] [CrossRef]
- Wen, A.; Srinivasan, U.; Goldberg, D.; Owen, J.; Marrs, C.F.; Misra, D.; Wing, D.A.; Ponnaluri, S.; Miles-Jay, A.; Bucholz, B.; et al. Selected vaginal bacteria and risk of preterm birth: An ecological perspective. J. Infect. Dis. 2014, 209, 1087–1094. [Google Scholar] [CrossRef]
- Cha, L.; Le, T.; Ve’e, T.; Ah Soon, N.T.; Tseng, W. Pacific Islanders in the Era of COVID-19: An Overlooked Community in Need. J. Racial Ethn. Health Disparities 2022, 9, 1347–1356. [Google Scholar] [CrossRef]
- Heyrana, K.J.; Kaneshiro, B.; Soon, R.; Nguyen, B.T.; Natavio, M.F. Data Equity for Asian American and Native Hawaiian and Other Pacific Islander People in Reproductive Health Research. Obs. Gynecol. 2023, 142, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Fialkowski, M.K.; Aflague, T.F.; Esquivel, M.K. A Reminder to Nutrition Researchers: Disaggregate Data on Native Hawaiians and Other Pacific Islanders From Asian Americans. J. Nutr. Educ. Behav. 2023, 55, 691–692. [Google Scholar] [CrossRef]
- Looijer-van Langen, M.; Hotte, N.; Dieleman, L.A.; Albert, E.; Mulder, C.; Madsen, K.L. Estrogen receptor-beta signaling modulates epithelial barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G621–G626. [Google Scholar] [CrossRef] [PubMed]
- Nuriel-Ohayon, M.; Neuman, H.; Ziv, O.; Belogolovski, A.; Barsheshet, Y.; Bloch, N.; Uzan, A.; Lahav, R.; Peretz, A.; Frishman, S.; et al. Progesterone Increases Bifidobacterium Relative Abundance during Late Pregnancy. Cell Rep. 2019, 27, 730–736.e733. [Google Scholar] [CrossRef] [PubMed]
- Walther-Antonio, M.R.; Jeraldo, P.; Berg Miller, M.E.; Yeoman, C.J.; Nelson, K.E.; Wilson, B.A.; White, B.A.; Chia, N.; Creedon, D.J. Pregnancy’s stronghold on the vaginal microbiome. PLoS ONE 2014, 9, e98514. [Google Scholar] [CrossRef]
- Gupta, P.; Singh, M.P.; Goyal, K. Diversity of Vaginal Microbiome in Pregnancy: Deciphering the Obscurity. Front. Public Health 2020, 8, 326. [Google Scholar] [CrossRef]
- Jenkins, D.J.; Woolston, B.M.; Hood-Pishchany, M.I.; Pelayo, P.; Konopaski, A.N.; Quinn Peters, M.; France, M.T.; Ravel, J.; Mitchell, C.M.; Rakoff-Nahoum, S.; et al. Bacterial amylases enable glycogen degradation by the vaginal microbiome. Nat. Microbiol. 2023, 8, 1641–1652. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Maximum Points | Standard for Maximum Score | Standard for Minimum Score |
|---|---|---|---|
| Points given for adequacy of consumption | |||
| Total Fruits | 5 | ≥0.8 c equivalents/1000 kcal | No fruit |
| Whole Fruits | 5 | ≥0.4 c equivalents/1000 kcal | No whole fruit |
| Total Vegetables | 5 | ≥1.1 c equivalents/1000 kcal | No vegetables |
| Greens and Beans | 5 | ≥0.2 c equivalents/1000 kcal | No dark green vegetables or beans and peas |
| Whole Grains | 10 | ≥1.5 oz equivalents/1000 kcal | No whole grains |
| Dairy | 10 | ≥1.3 c equivalents/1000 kcal | No dairy |
| Total Protein Foods | 5 | ≥2.5 oz equivalents/1000 kcal | No protein foods |
| Seafood and Plant Proteins | 5 | ≥0.8 oz equivalents/1000 kcal | No seafood or plant proteins |
| Fatty Acids | 10 | (PUFAs a + MUFAs b)/SFAs c ≥ 2.5 | (PUFAs + MUFAs)/SFAs ≤ 1.2 |
| Points given for moderation of consumption | |||
| Refined Grains | 10 | ≤1.8 oz equivalents/1000 kcal | ≥4.3 oz equivalents/1000 kcal |
| Sodium | 10 | ≤1.1 g/1000 kcal | ≥2.0 g/1000 kcal |
| Added Sugars | 10 | ≤6.5% of energy | ≥26% of energy |
| Saturated Fats | 10 | ≤8% of energy | ≥16% of energy |
| Filipino n = 10 | Japanese n = 10 | Native Hawaiian n =10 | Non-Hispanic White n = 10 | p-Value | |
|---|---|---|---|---|---|
| Age (median, [SD]) | 25 [5.03] | 34 [5.02] | 31 [6.03] | 32 [5.55] | NS |
| Parity (median) | 0 | 0.5 | 0 | 1 | |
| Obesity 1 | 1 | 3 | 6 | 3 | |
| Excess Gestational Weight Gain 2 | 0 | 0 | 6 | 3 | |
| Mode of Delivery | |||||
| Cesarean Delivery | 3 | 3 | 4 | 0 | |
| Vaginal Delivery | 7 | 7 | 6 | 10 | |
| Pregnancy Complications | |||||
| Gestational Diabetes | 0 | 1 | 2 | 1 | |
| Preeclampsia | 1 | 5 | 2 | 1 | |
| Spontaneous Preterm Labor | 1 | 0 | 0 | 0 | |
| Healthy Eating Index Score (median) [Interquartile Range] | 55.71 [52.59, 57.41] | 60.38 [53.08, 67.41] | 65.38 [53.70, 75.85] | 60.68 [55.38, 65.38] | 0.032 a |
| Macronutrient Intake | |||||
| Percent of Energy from Protein | 16.24 (2.11) | 16.29 (2.32) | 16.37 (2.90) | 15.85 (2.01) | NS |
| Percent Energy from Carbohydrate | 50.15 (6.35) | 45.60 (6.29) | 52.33 (6.94) b | 47.19 (5.19) | 0.017 |
| Percent Energy from Fat | 34.78 (4.88) | 37.67 (5.63) | 33.11 (5.32) | 36.47 (3.88) | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, C.; Morikawa, K.; Benny, P.; Riel, J.; Fialkowski, M.K.; Qin, Y.; Khadka, V.; Lee, M.-J. Effects of Dietary Quality on Vaginal Microbiome Composition Throughout Pregnancy in a Multi-Ethnic Cohort. Nutrients 2024, 16, 3405. https://doi.org/10.3390/nu16193405
Miller C, Morikawa K, Benny P, Riel J, Fialkowski MK, Qin Y, Khadka V, Lee M-J. Effects of Dietary Quality on Vaginal Microbiome Composition Throughout Pregnancy in a Multi-Ethnic Cohort. Nutrients. 2024; 16(19):3405. https://doi.org/10.3390/nu16193405
Chicago/Turabian StyleMiller, Corrie, Kira Morikawa, Paula Benny, Jonathan Riel, Marie K. Fialkowski, Yujia Qin, Vedbar Khadka, and Men-Jean Lee. 2024. "Effects of Dietary Quality on Vaginal Microbiome Composition Throughout Pregnancy in a Multi-Ethnic Cohort" Nutrients 16, no. 19: 3405. https://doi.org/10.3390/nu16193405
APA StyleMiller, C., Morikawa, K., Benny, P., Riel, J., Fialkowski, M. K., Qin, Y., Khadka, V., & Lee, M.-J. (2024). Effects of Dietary Quality on Vaginal Microbiome Composition Throughout Pregnancy in a Multi-Ethnic Cohort. Nutrients, 16(19), 3405. https://doi.org/10.3390/nu16193405