
Supplementation with a Whey Protein Concentrate Enriched in Bovine Milk Exosomes Improves Longitudinal Growth and Supports Bone Health During Catch-Up Growth in Rats


 ,
,  , , , , , and
, , , , , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bovine Milk Exosome Enrichment Methods and Supplementation of the Experimental Diet
2.2. In Vitro Research and Experimental Design
2.3. Cell Growth Test
2.4. Cell Cycle Analysis
2.5. Clonogenic Assay
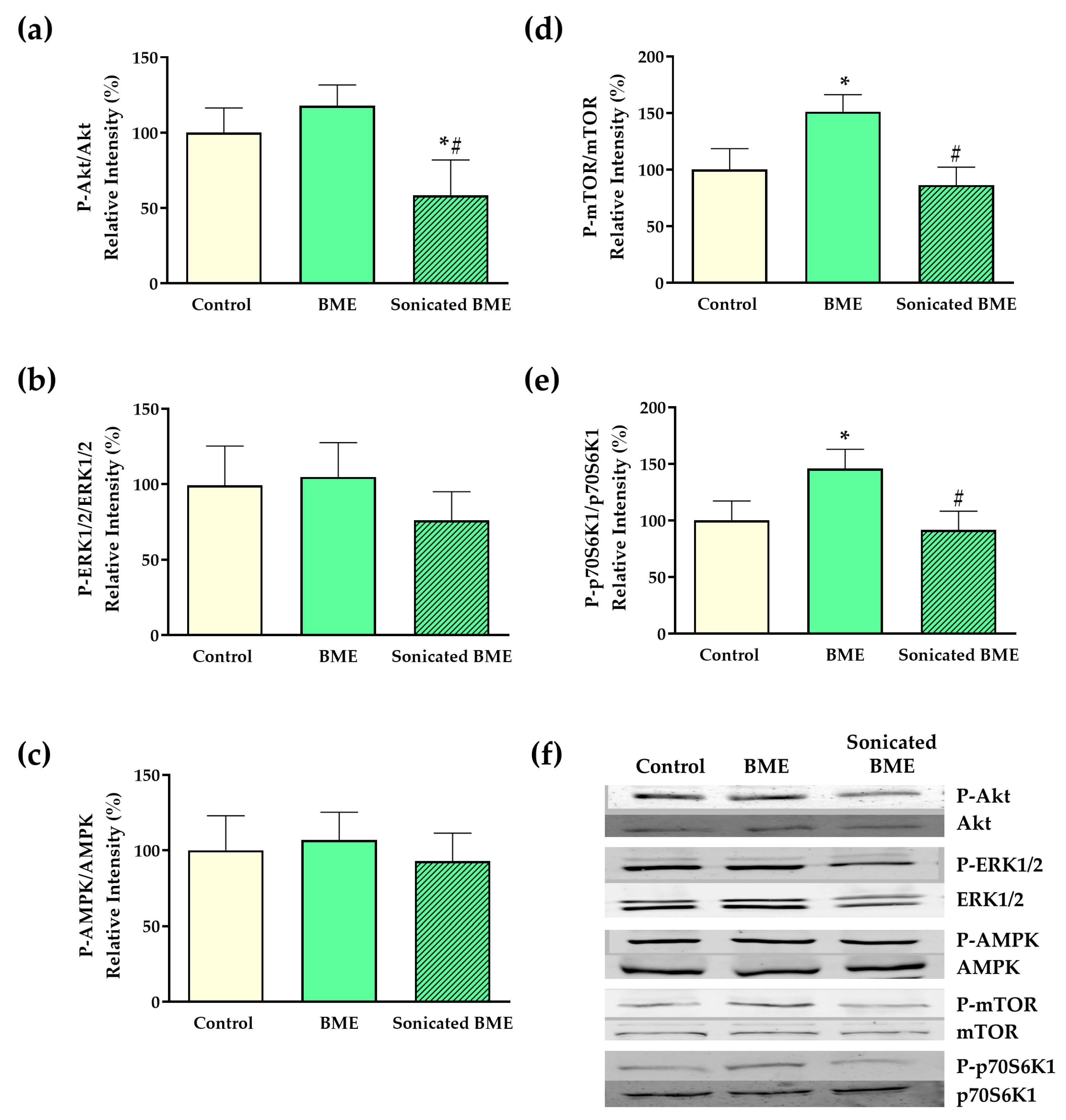
2.6. Cell Signaling Assessment
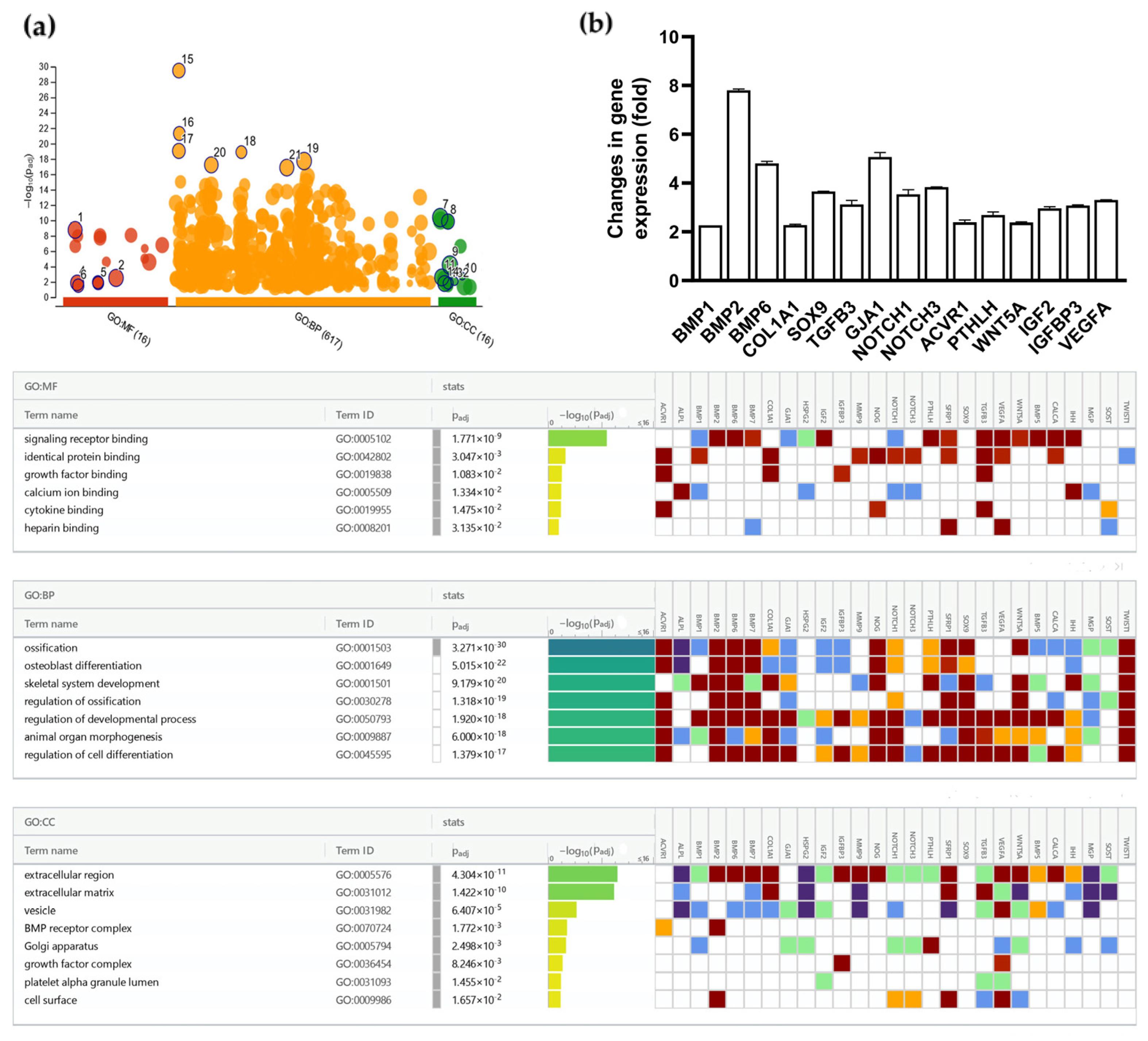
2.7. Gene Profiler PCR Array
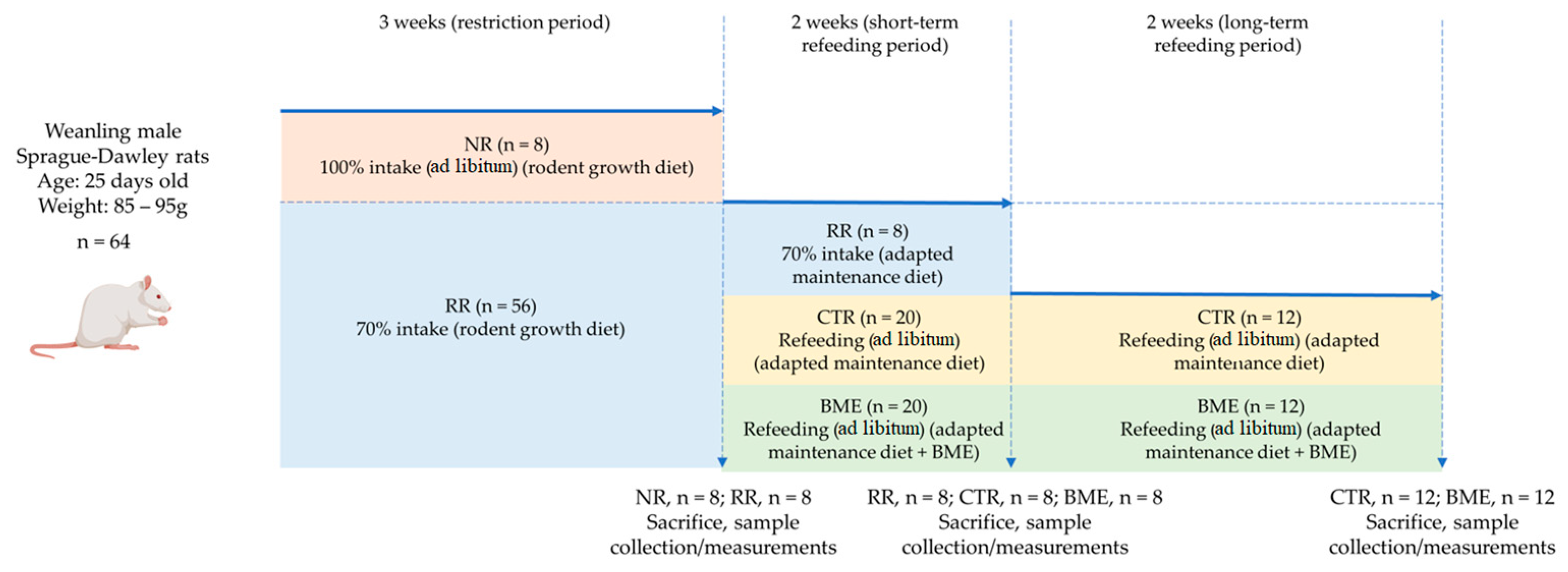
2.8. In Vivo Research and Experimental Design
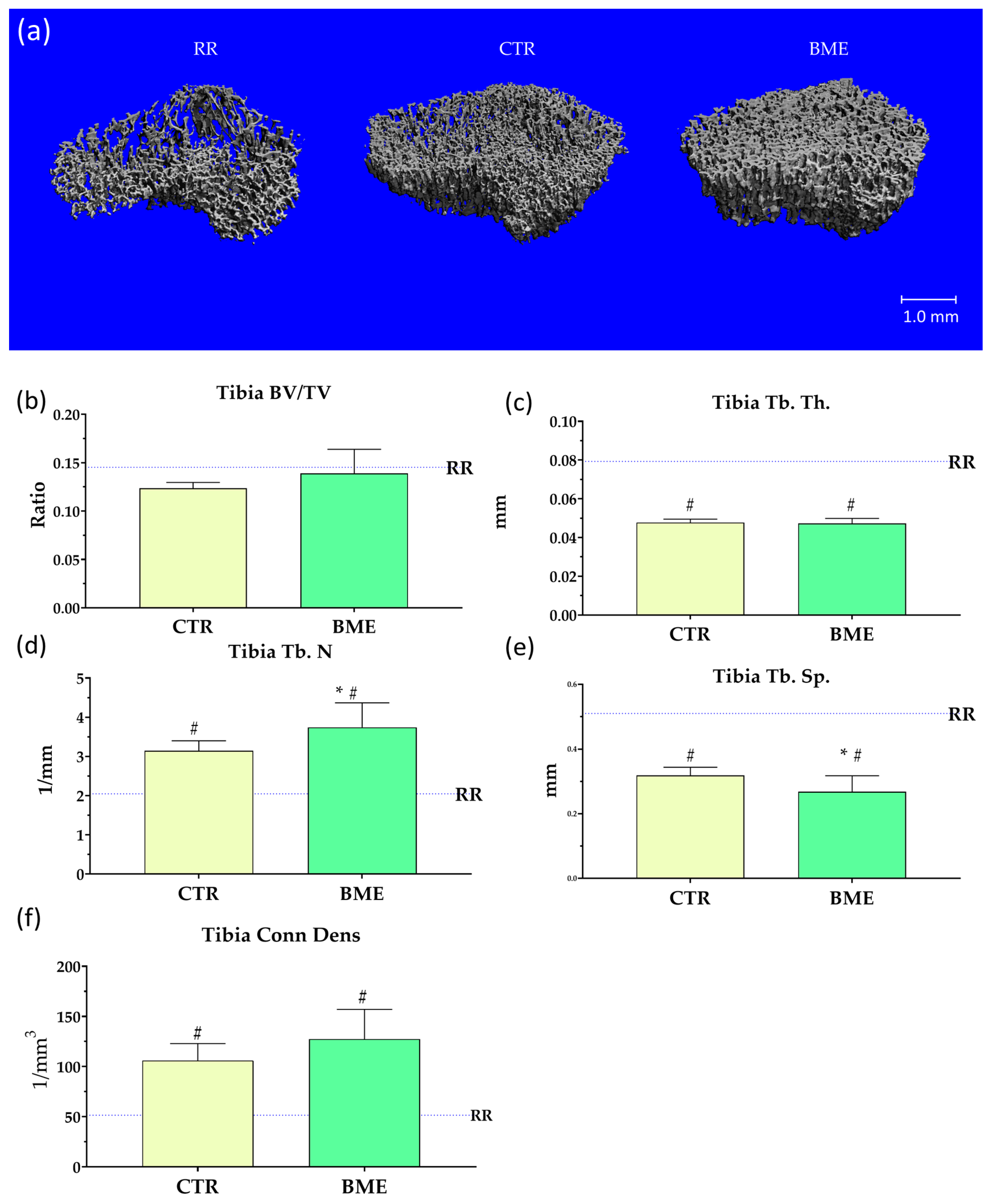
2.9. Ex Vivo Measurements, Densitometry, and Micro-CT Analyses
2.10. Statistical Analysis
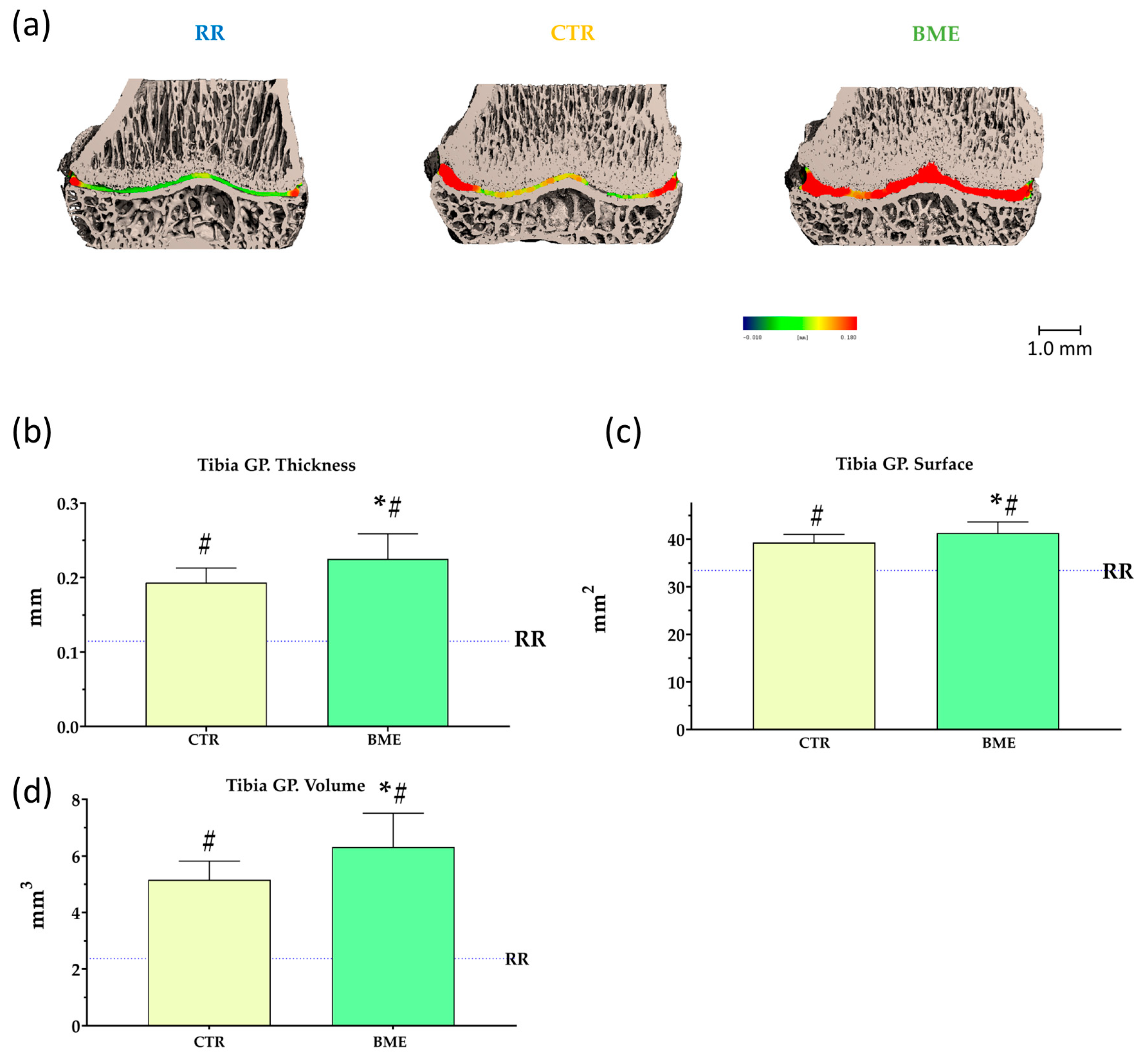
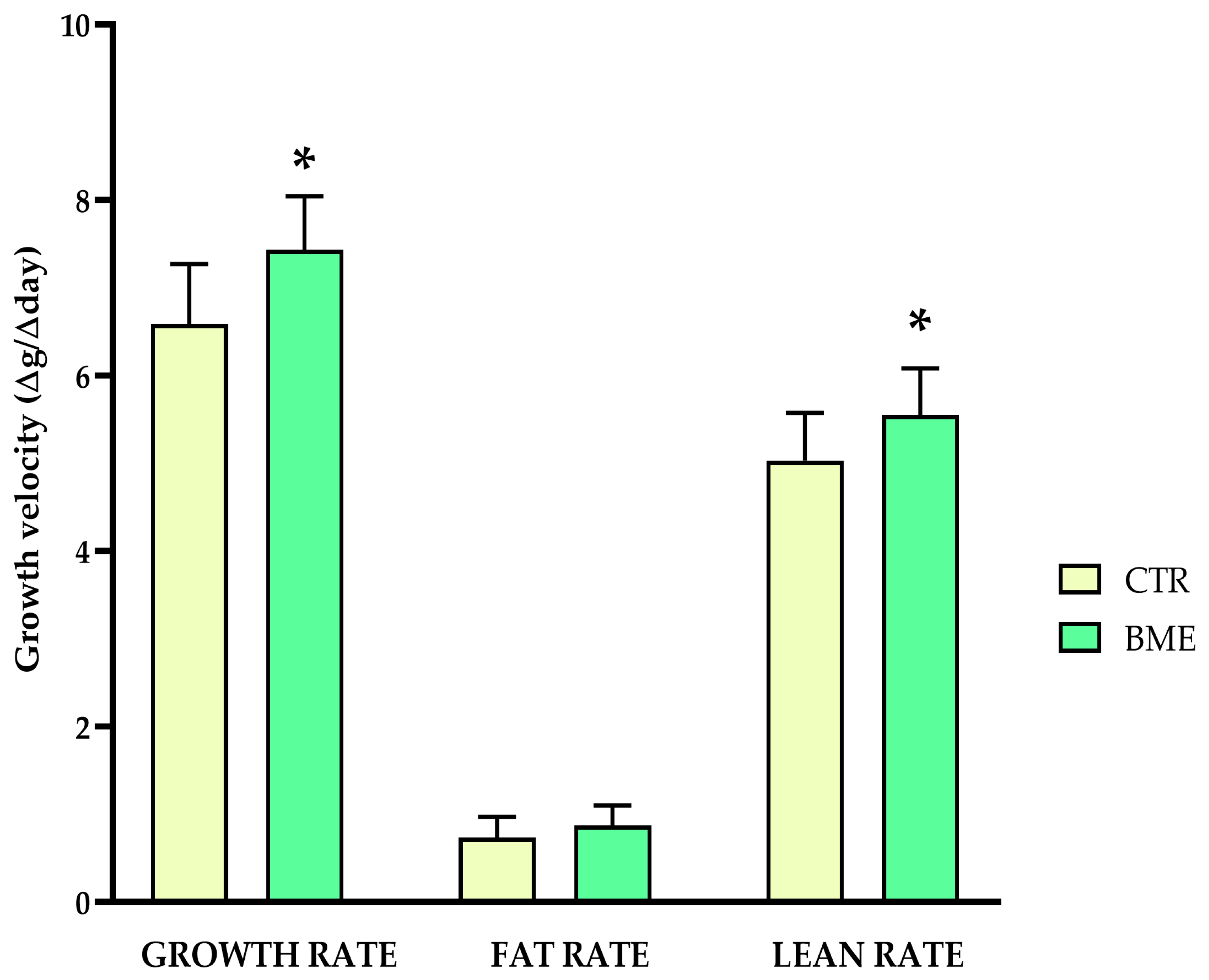
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- WHO. WHO Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Campisi, S.C.; Carducci, B.; Söder, O.; Bhutta, Z.A. The intricate relationship between chronic undernutrition, impaired linear growth and delayed puberty: Is ‘catch-up’growth possible during adolescence? Innocenti Work. Pap. 2018. [Google Scholar] [CrossRef]
- Ssentongo, P.; Ssentongo, A.E.; Ba, D.M.; Ericson, J.E.; Na, M.; Gao, X.; Fronterre, C.; Chinchilli, V.M.; Schiff, S.J. Global, regional and national epidemiology and prevalence of child stunting, wasting and underweight in low- and middle-income countries, 2006–2018. Sci. Rep. 2021, 11, 5204. [Google Scholar] [CrossRef] [PubMed]
- Olofin, I.; McDonald, C.M.; Ezzati, M.; Flaxman, S.; Black, R.E.; Fawzi, W.W.; Caulfield, L.E.; Danaei, G. Associations of suboptimal growth with all-cause and cause-specific mortality in children under five years: A pooled analysis of ten prospective studies. PLoS ONE 2013, 8, e64636. [Google Scholar] [CrossRef]
- Benjamin-Chung, J.; Mertens, A.; Colford, J.M.; Hubbard, A.E.; van der Laan, M.J.; Coyle, J.; Sofrygin, O.; Cai, W.; Nguyen, A.; Pokpongkiat, N.N.; et al. Early-childhood linear growth faltering in low- and middle-income countries. Nature 2023, 621, 550–557. [Google Scholar] [CrossRef]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in ∼700,000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef]
- Inzaghi, E.; Pampanini, V.; Deodati, A.; Cianfarani, S. The Effects of Nutrition on Linear Growth. Nutrients 2022, 14, 1752. [Google Scholar] [CrossRef]
- Huizinga, C.T.; Engelbregt, M.J.T.; Rekers-Mombarg, L.T.M.; Vaessen, S.F.C.; Delemarre-van de Waal, H.A.; Fodor, M. Ligation of the Uterine Artery and Early Postnatal Food Restriction—Animal Models for Growth Retardation. Horm. Res. 2004, 62, 233–240. [Google Scholar] [CrossRef]
- Wit, J.M.; Boersma, B. Catch-up growth: Definition, mechanisms, and models. J. Pediatr. Endocrinol. Metab. JPEM 2002, 15 (Suppl. S5), 1229–1241. [Google Scholar] [PubMed]
- Singhal, A. Long-Term Adverse Effects of Early Growth Acceleration or Catch-Up Growth. Ann. Nutr. Metab. 2017, 70, 236–240. [Google Scholar] [CrossRef]
- Wells, J.C. The thrifty phenotype: An adaptation in growth or metabolism? Am. J. Hum. Biol. 2011, 23, 65–75. [Google Scholar] [CrossRef]
- Stevens, D.G.; Boyer, M.I.; Bowen, C.V.A. Transplantation of Epiphyseal Plate Allografts Between Animals of Different Ages. J. Pediatr. Orthop. 1999, 19, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Ağırdil, Y. The growth plate: A physiologic overview. EFORT Open Rev. 2020, 5, 498–507. [Google Scholar] [CrossRef]
- Hallett, S.A.; Ono, W.; Ono, N. Growth Plate Chondrocytes: Skeletal Development, Growth and Beyond. Int. J. Mol. Sci. 2019, 20, 6009. [Google Scholar] [CrossRef]
- Gat-Yablonski, G.; Shtaif, B.; Abraham, E.; Phillip, M. Nutrition-induced catch-up growth at the growth plate. J. Pediatr. Endocrinol. Metab. 2008, 21, 879–894. [Google Scholar] [CrossRef]
- Lui, J.C.; Nilsson, O.; Baron, J. Growth plate senescence and catch-up growth. Cartil. Bone Dev. Its Disord. 2011, 21, 23–29. [Google Scholar]
- Chagin, A.S.; Karimian, E.; Sundström, K.; Eriksson, E.; Sävendahl, L. Catch-up growth after dexamethasone withdrawal occurs in cultured postnatal rat metatarsal bones. J. Endocrinol. 2010, 22, 21. [Google Scholar] [CrossRef] [PubMed]
- Pando, R.; Masarwi, M.; Shtaif, B.; Idelevich, A.; Monsonego-Ornan, E.; Shahar, R.; Phillip, M.; Gat-Yablonski, G. Bone quality is affected by food restriction and by nutrition-induced catch-up growth. J. Endocrinol. 2014, 223, 227–239. [Google Scholar] [CrossRef]
- Golden, N.H.; Abrams, S.A. Optimizing bone health in children and adolescents. Pediatrics 2014, 134, e1229–e1243. [Google Scholar] [CrossRef]
- Yoshimura, T.; Tohya, T.; Onoda, C.; Okamura, H. Poor nutrition in prepubertal Japanese children at the end of World War II suppressed bone development. Maturitas 2005, 52, 32–34. [Google Scholar] [CrossRef]
- Masarwi, M.; Gabet, Y.; Dolkart, O.; Brosh, T.; Shamir, R.; Phillip, M.; Gat-Yablonski, G. Skeletal effect of casein and whey protein intake during catch-up growth in young male Sprague–Dawley rats. Br. J. Nutr. 2016, 116, 59–69. [Google Scholar] [CrossRef]
- Bar-Maisels, M.; Gabet, Y.; Shamir, R.; Hiram-Bab, S.; Pasmanik-Chor, M.; Phillip, M.; Bar-Yoseph, F.; Gat-Yablonski, G. Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats. Nutrients 2017, 9, 764. [Google Scholar] [CrossRef] [PubMed]
- Bueno-Vargas, P.; Manzano, M.; Pérez-Castillo, Í.M.; Rueda, R.; López-Pedrosa, J.M. Dietary complex and slow digestive carbohydrates promote bone mass and improve bone microarchitecture during catch-up growth in rats. Nutrients 2022, 14, 1303. [Google Scholar] [CrossRef] [PubMed]
- Oichi, T.; Kodama, J.; Wilson, K.; Tian, H.; Imamura Kawasawa, Y.; Usami, Y.; Oshima, Y.; Saito, T.; Tanaka, S.; Iwamoto, M.; et al. Nutrient-regulated dynamics of chondroprogenitors in the postnatal murine growth plate. Bone Res. 2023, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- García-Martínez, J.; Pérez-Castillo, Í.M.; Salto, R.; López-Pedrosa, J.M.; Rueda, R.; Girón, M.D. Beneficial effects of bovine milk exosomes in metabolic interorgan cross-talk. Nutrients 2022, 14, 1442. [Google Scholar] [CrossRef]
- Kim, J.; Guan, K.-L. mTOR as a central hub of nutrient signalling and cell growth. Nat. Cell Biol. 2019, 21, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Long, F. mTOR signaling in skeletal development and disease. Bone Res. 2018, 6, 1. [Google Scholar] [CrossRef]
- Melnik, B.C.; John, S.M.; Schmitz, G. Milk is not just food but most likely a genetic transfection system activating mTORC1 signaling for postnatal growth. Nutr. J. 2013, 12, 103. [Google Scholar] [CrossRef]
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-derived microRNAs of human milk and their effects on infant health and development. Biomolecules 2021, 11, 851. [Google Scholar] [CrossRef]
- Yan, B.; Zhang, Z.; Jin, D.; Cai, C.; Jia, C.; Liu, W.; Wang, T.; Li, S.; Zhang, H.; Huang, B.; et al. mTORC1 regulates PTHrP to coordinate chondrocyte growth, proliferation and differentiation. Nat. Commun. 2016, 7, 11151. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Arntz, O.J.; Davidson, E.N.B.; van Lent, P.L.; Koenders, M.I.; van der Kraan, P.M.; van den Berg, W.B.; Ferreira, A.V.; van de Loo, F.A. Milk extracellular vesicles accelerate osteoblastogenesis but impair bone matrix formation. J. Nutr. Biochem. 2016, 30, 74–84. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Di Ceglie, I.; Arntz, O.J.; Van den Berg, W.B.; Van den Hoogen, F.H.; Ferreira, A.V.; Van Lent, P.L.; Van De Loo, F.A. Milk-derived nanoparticle fraction promotes the formation of small osteoclasts but reduces bone resorption. J. Cell. Physiol. 2017, 232, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Liu, Q.; Zheng, T.; Li, J.; Zhang, T.; Yao, Y.; Liu, Y.; Lin, K.; Liu, T.; Gong, P. Oral milk-derived extracellular vesicles inhibit osteoclastogenesis and ameliorate bone loss in ovariectomized mice by improving gut microbiota. J. Agric. Food Chem. 2024, 72, 4726–4736. [Google Scholar] [CrossRef]
- Oliveira, M.C.; Pieters, B.C.; Guimarães, P.B.; Duffles, L.F.; Heredia, J.E.; Silveira, A.L.; Oliveira, A.C.; Teixeira, M.M.; Ferreira, A.V.; Silva, T.A. Bovine milk extracellular vesicles are osteoprotective by increasing osteocyte numbers and targeting RANKL/OPG system in experimental models of bone loss. Front. Bioeng. Biotechnol. 2020, 8, 891. [Google Scholar] [CrossRef]
- Yun, B.; Maburutse, B.; Kang, M.; Park, M.; Park, D.; Kim, Y.; Oh, S. Dietary bovine milk–derived exosomes improve bone health in an osteoporosis-induced mouse model. J. Dairy Sci. 2020, 103, 7752–7760. [Google Scholar] [CrossRef]
- Go, G.; Jeon, J.; Lee, G.; Lee, J.H.; Lee, S.H. Bovine milk extracellular vesicles induce the proliferation and differentiation of osteoblasts and promote osteogenesis in rats. J. Food Biochem. 2021, 45, e13705. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, O.; Baron, J. Fundamental limits on longitudinal bone growth: Growth plate senescence and epiphyseal fusion. Trends Endocrinol. Metab. 2004, 15, 370–374. [Google Scholar] [CrossRef]
- Andreu, Z.; Yáñez-Mó, M. Tetraspanins in Extracellular Vesicle Formation and Function. Front. Immunol. 2014, 5, 442. [Google Scholar] [CrossRef]
- Reinhardt, T.A.; Lippolis, J.D.; Nonnecke, B.J.; Sacco, R.E. Bovine milk exosome proteome. J. Proteom. 2012, 75, 1486–1492. [Google Scholar] [CrossRef]
- Sukreet, S.; Silva, B.V.R.E.; Adamec, J.; Cui, J.; Zempleni, J. Sonication and Short-term Incubation Alter the Content of Bovine Milk Exosome Cargos and Exosome Bioavailability (OR26-08-19). Curr. Dev. Nutr. 2019, 3, nzz033.OR026-008-019. [Google Scholar] [CrossRef]
- Ormerod, M. Analysis of DNA-general methods. In Flow Cytometry: A Practical Approach; IRI Press: Washington, DC, USA, 1996; pp. 119–135. [Google Scholar]
- Girón, M.D.; Sevillano, N.; Vargas, A.M.; Domínguez, J.; Guinovart, J.J.; Salto, R. The glucose-lowering agent sodium tungstate increases the levels and translocation of GLUT4 in L6 myotubes through a mechanism associated with ERK1/2 and MEF2D. Diabetologia 2008, 51, 1285–1295. [Google Scholar] [CrossRef]
- Raudvere, U.; Kolberg, L.; Kuzmin, I.; Arak, T.; Adler, P.; Peterson, H.; Vilo, J. g: Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019, 47, W191–W198. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, T.; Rüegsegger, P. A new method for the model-independent assessment of thickness in three-dimensional images. J. Microsc. 1997, 185, 67–75. [Google Scholar] [CrossRef]
- Shaikh, A.; Wesner, A.A.; Abuhattab, M.; Kutty, R.G.; Premnath, P. Cell cycle regulators and bone: Development and regeneration. Cell Biosci. 2023, 13, 35. [Google Scholar] [CrossRef]
- Dy, P.; Wang, W.; Bhattaram, P.; Wang, Q.; Wang, L.; Ballock, R.T.; Lefebvre, V. Sox9 directs hypertrophic maturation and blocks osteoblast differentiation of growth plate chondrocytes. Dev. Cell 2012, 22, 597–609. [Google Scholar] [CrossRef]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124 Pt 20, 3428–3440. [Google Scholar] [CrossRef]
- Duyar, I.; Pelin, C. Body height estimation based on tibia length in different stature groups. Am. J. Phys. Anthropol. 2003, 122, 23–27. [Google Scholar] [CrossRef]
- Tsai, A.; Stamoulis, C.; Barber, I.; Kleinman, P.K. Infant lower extremity long bone growth rates: Comparison of contemporary with early 20th century data using mixed effect models. Am. J. Hum. Biol. 2017, 29, e22905. [Google Scholar] [CrossRef]
- Gat-Yablonski, G.; Yackobovitch-Gavan, M.; Phillip, M. Which dietary components modulate longitudinal growth? Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 211–216. [Google Scholar] [CrossRef]
- Bar-Maisels, M.; Menahem, C.; Gabet, Y.; Hiram-Bab, S.; Phillip, M.; Gat-Yablonski, G. Different effects of soy and whey on linear bone growth and growth pattern in young male sprague-dawley rats. Front. Nutr. 2021, 8, 739607. [Google Scholar] [CrossRef]
- Yackobovitch-Gavan, M.; Phillip, M.; Gat-Yablonski, G. How milk and its proteins affect growth, bone health, and weight. Horm. Res. Paediatr. 2017, 88, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Sukreet, S.; Pereira Braga, C.; An, T.T.; Adamec, J.; Cui, J.; Zempleni, J. Ultrasonication of Milk Decreases the Content of Exosomes and MicroRNAs in an Exosome-Defined Rodent Diet. J. Nutr. 2022, 152, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.S.; Wu, K.Y.; Auyeung, V.; Chen, Q.; Gruppuso, P.A.; Phornphutkul, C. Leucine restriction inhibits chondrocyte proliferation and differentiation through mechanisms both dependent and independent of mTOR signaling. Am. J. Physiol.-Endocrinol. Metab. 2009, 296, E1374–E1382. [Google Scholar] [CrossRef]
- Phornphutkul, C.; Wu, K.-Y.; Auyeung, V.; Chen, Q.; Gruppuso, P.A. mTOR signaling contributes to chondrocyte differentiation. Dev. Dyn. 2008, 237, 702–712. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, O.; Carbajo-Pérez, E.; Garcia, E.; Gil, H.; Molinos, I.; Rodriguez, J.; Ordoñez, F.A.; Santos, F. Rapamycin retards growth and causes marked alterations in the growth plate of young rats. Pediatr. Nephrol. 2007, 22, 954–961. [Google Scholar] [CrossRef]
- Álvarez-García, Ó.; García-López, E.; Loredo, V.; Gil-Peña, H.; Rodríguez-Suárez, J.; Ordóñez, F.Á.; Carbajo-Pérez, E.; Santos, F. Rapamycin induces growth retardation by disrupting angiogenesis in the growth plate. Kidney Int. 2010, 78, 561–568. [Google Scholar] [CrossRef] [PubMed]
- Even-Zohar, N.; Jacob, J.; Amariglio, N.; Rechavi, G.; Potievsky, O.; Phillip, M.; Gat-Yablonski, G. Nutrition-induced catch-up growth increases hypoxia inducible factor 1α RNA levels in the growth plate. Bone 2008, 42, 505–515. [Google Scholar] [CrossRef]
- Farnum, C.E.; Lee, A.O.; O’Hara, K.; Wilsman, N.J. Effect of Short-Term Fasting on Bone Elongation Rates: An Analysis of Catch-up Growth in Young Male Rats. Pediatr. Res. 2003, 53, 33–41. [Google Scholar] [CrossRef]
- Guedes, P.M.S.G.; Zamarioli, A.; Botega, I.I.; Silva, R.A.B.d.; Issa, J.P.M.; Butezloff, M.M.; Sousa, Y.T.C.S.; Ximenez, J.P.B.; Volpon, J.B. Undernutrition impairs the quality of growth plate and trabecular and cortical bones in growing rats. Acta Cir. Bras. 2019, 34, e201900301. [Google Scholar] [CrossRef]
- Devlin, M.J.; Cloutier, A.M.; Thomas, N.A.; Panus, D.A.; Lotinun, S.; Pinz, I.; Baron, R.; Rosen, C.J.; Bouxsein, M.L. Caloric restriction leads to high marrow adiposity and low bone mass in growing mice. J. Bone Miner. Res. 2010, 25, 2078–2088. [Google Scholar] [CrossRef]
- Ghasemi, A.; Jeddi, S.; Kashfi, K. The laboratory rat: Age and body weight matter. EXCLI J. 2021, 20, 1431–1445. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624. [Google Scholar] [PubMed]
- Khanam, A.; Ngu, A.; Zempleni, J. Bioavailability of orally administered small extracellular vesicles from bovine milk in C57BL/6J mice. Int. J. Pharm. 2023, 639, 122974. [Google Scholar] [CrossRef] [PubMed]
- Manca, S.; Upadhyaya, B.; Mutai, E.; Desaulniers, A.T.; Cederberg, R.A.; White, B.R.; Zempleni, J. Milk exosomes are bioavailable and distinct microRNA cargos have unique tissue distribution patterns. Sci. Rep. 2018, 8, 11321. [Google Scholar] [CrossRef]
- Golan-Gerstl, R.; Elbaum Shiff, Y.; Moshayoff, V.; Schecter, D.; Leshkowitz, D.; Reif, S. Characterization and biological function of milk-derived miRNAs. Mol. Nutr. Food Res. 2017, 61, 1700009. [Google Scholar] [CrossRef]
- Izumi, H.; Tsuda, M.; Sato, Y.; Kosaka, N.; Ochiya, T.; Iwamoto, H.; Namba, K.; Takeda, Y. Bovine milk exosomes contain microRNA and mRNA and are taken up by human macrophages. J. Dairy Sci. 2015, 98, 2920–2933. [Google Scholar] [CrossRef]
- Lian, W.-S.; Ko, J.-Y.; Chen, Y.-S.; Ke, H.-J.; Hsieh, C.-K.; Kuo, C.-W.; Wang, S.-Y.; Huang, B.-W.; Tseng, J.-G.; Wang, F.-S. MicroRNA-29a represses osteoclast formation and protects against osteoporosis by regulating PCAF-mediated RANKL and CXCL12. Cell Death Dis. 2019, 10, 705. [Google Scholar] [CrossRef]
- Liu, H.; Su, H.; Wang, X.; Hao, W. MiR-148a regulates bone marrow mesenchymal stem cells-mediated fracture healing by targeting insulin-like growth factor 1. J. Cell. Biochem. 2019, 120, 1350–1361. [Google Scholar] [CrossRef]
- Yang, C.; Liu, X.; Zhao, K.; Zhu, Y.; Hu, B.; Zhou, Y.; Wang, M.; Wu, Y.; Zhang, C.; Xu, J.; et al. miRNA-21 promotes osteogenesis via the PTEN/PI3K/Akt/HIF-1α pathway and enhances bone regeneration in critical size defects. Stem Cell Res. Ther. 2019, 10, 65. [Google Scholar] [CrossRef]
- Dai, J.; Dong, R.; Han, X.; Li, J.; Gong, X.; Bai, Y.; Kang, F.; Liang, M.; Zeng, F.; Hou, Z.; et al. Osteoclast-derived exosomal let-7a-5p targets Smad2 to promote the hypertrophic differentiation of chondrocytes. Am. J. Physiol.-Cell Physiol. 2020, 319, C21–C33. [Google Scholar] [CrossRef]
- Jee, Y.H.; Wang, J.; Yue, S.; Jennings, M.; Clokie, S.J.; Nilsson, O.; Lui, J.C.; Baron, J. mir-374-5p, mir-379-5p, and mir-503-5p Regulate Proliferation and Hypertrophic Differentiation of Growth Plate Chondrocytes in Male Rats. Endocrinology 2018, 159, 1469–1478. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.; Zhang, W.; Zhou, C.; Zhao, G.; Zhong, H.; Hang, K.; Xu, J.; Zhang, W.; Chen, E.; Wu, J.; et al. MFG-E8 promotes osteogenic differentiation of human bone marrow mesenchymal stem cells through GSK3β/β-catenin signaling pathway. FASEB J. 2023, 37, e22950. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, L.; Pan, J.; Luo, B.; Zeng, H.; Shao, Y.; Zhang, H.; Guan, H.; Guo, D.; Zeng, C.; et al. MFG-E8 regulated by miR-99b-5p protects against osteoarthritis by targeting chondrocyte senescence and macrophage reprogramming via the NF-κB pathway. Cell Death Dis. 2021, 12, 533. [Google Scholar] [CrossRef]
- Pieters, B.C.; Arntz, O.J.; Bennink, M.B.; Broeren, M.G.; van Caam, A.P.; Koenders, M.I.; van Lent, P.L.; van den Berg, W.B.; de Vries, M.; van der Kraan, P.M. Commercial cow milk contains physically stable extracellular vesicles expressing immunoregulatory TGF-β. PLoS ONE 2015, 10, e0121123. [Google Scholar] [CrossRef]
- Dangelo, M.; Sarment, D.P.; Billings, P.C.; Pacifici, M. Activation of transforming growth factor beta in chondrocytes undergoing endochondral ossification. J. Bone Miner. Res. Off. J. Am. Soc. Bone Miner. Res. 2001, 16, 2339–2347. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.; Zhang, S.; Liu, Q.; Huang, C.; Hao, H.; Tan, M.S.; Yu, X.; Lou, C.K.L.; Huang, R.; Zhang, Z. Milk-derived extracellular vesicles protect intestinal barrier integrity in the gut-liver axis. Sci. Adv. 2023, 9, eade5041. [Google Scholar] [CrossRef]
- Liu, Q.; Hao, H.; Li, J.; Zheng, T.; Yao, Y.; Tian, X.; Zhang, Z.; Yi, H. Oral Administration of Bovine Milk-Derived Extracellular Vesicles Attenuates Cartilage Degeneration via Modulating Gut Microbiota in DMM-Induced Mice. Nutrients 2023, 15, 747. [Google Scholar] [CrossRef]
- Luo, Y.; Bi, J.; Lin, Y.; He, J.; Wu, S.; Zhang, Y.; Wang, Y.; Song, S.; Guo, H. Milk-derived small extracellular vesicles promote bifidobacteria growth by accelerating carbohydrate metabolism. LWT 2023, 182, 114866. [Google Scholar] [CrossRef]
- Zhou, F.; Paz, H.A.; Sadri, M.; Fernando, S.C.; Zempleni, J. A diet defined by its content of bovine milk exosomes alters the composition of the intestinal microbiome in C57BL/6 mice. FASEB J. 2017, 31, 965.924. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Measurements | Animal Group | ||
|---|---|---|---|
| RR | CTR | BME | |
| Total Body Length (mm) | 188.8 ± 8.78 | 208.4 ± 3.52 # | 207.8 ± 5.22 # |
| Tibia Length (mm) | 36.37 ± 1.26 | 38.64 ± 0.66 # | 38.77 ± 0.59 # |
| BMD (mg/cm2) | 124 ± 6.16 | 138 ± 3.39 # | 143.4 ± 5.24 #,* |
| BMC (g) | 0.192 ± 0.02 | 0.261 ± 0.007 # | 0.271 ± 0.02 # |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Martínez, J.; Salto, R.; Girón, M.D.; Pérez-Castillo, Í.M.; Bueno Vargas, P.; Vílchez, J.D.; Linares-Pérez, A.; Manzano, M.; García-Córcoles, M.T.; Rueda, R.; et al. Supplementation with a Whey Protein Concentrate Enriched in Bovine Milk Exosomes Improves Longitudinal Growth and Supports Bone Health During Catch-Up Growth in Rats. Nutrients 2024, 16, 3814. https://doi.org/10.3390/nu16223814
García-Martínez J, Salto R, Girón MD, Pérez-Castillo ÍM, Bueno Vargas P, Vílchez JD, Linares-Pérez A, Manzano M, García-Córcoles MT, Rueda R, et al. Supplementation with a Whey Protein Concentrate Enriched in Bovine Milk Exosomes Improves Longitudinal Growth and Supports Bone Health During Catch-Up Growth in Rats. Nutrients. 2024; 16(22):3814. https://doi.org/10.3390/nu16223814
Chicago/Turabian StyleGarcía-Martínez, Jorge, Rafael Salto, María D. Girón, Íñigo M. Pérez-Castillo, Pilar Bueno Vargas, Jose D. Vílchez, Azahara Linares-Pérez, Manuel Manzano, María T. García-Córcoles, Ricardo Rueda, and et al. 2024. "Supplementation with a Whey Protein Concentrate Enriched in Bovine Milk Exosomes Improves Longitudinal Growth and Supports Bone Health During Catch-Up Growth in Rats" Nutrients 16, no. 22: 3814. https://doi.org/10.3390/nu16223814
APA StyleGarcía-Martínez, J., Salto, R., Girón, M. D., Pérez-Castillo, Í. M., Bueno Vargas, P., Vílchez, J. D., Linares-Pérez, A., Manzano, M., García-Córcoles, M. T., Rueda, R., & López-Pedrosa, J. M. (2024). Supplementation with a Whey Protein Concentrate Enriched in Bovine Milk Exosomes Improves Longitudinal Growth and Supports Bone Health During Catch-Up Growth in Rats. Nutrients, 16(22), 3814. https://doi.org/10.3390/nu16223814