
Oat-Protein-Based Diet Lowers Blood Pressure and Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Rats

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Ethics
2.2. Oat Protein Extraction and Oat-Protein-Feeding Study Design
2.3. Transthoracic Echocardiography
2.4. BP Measurements
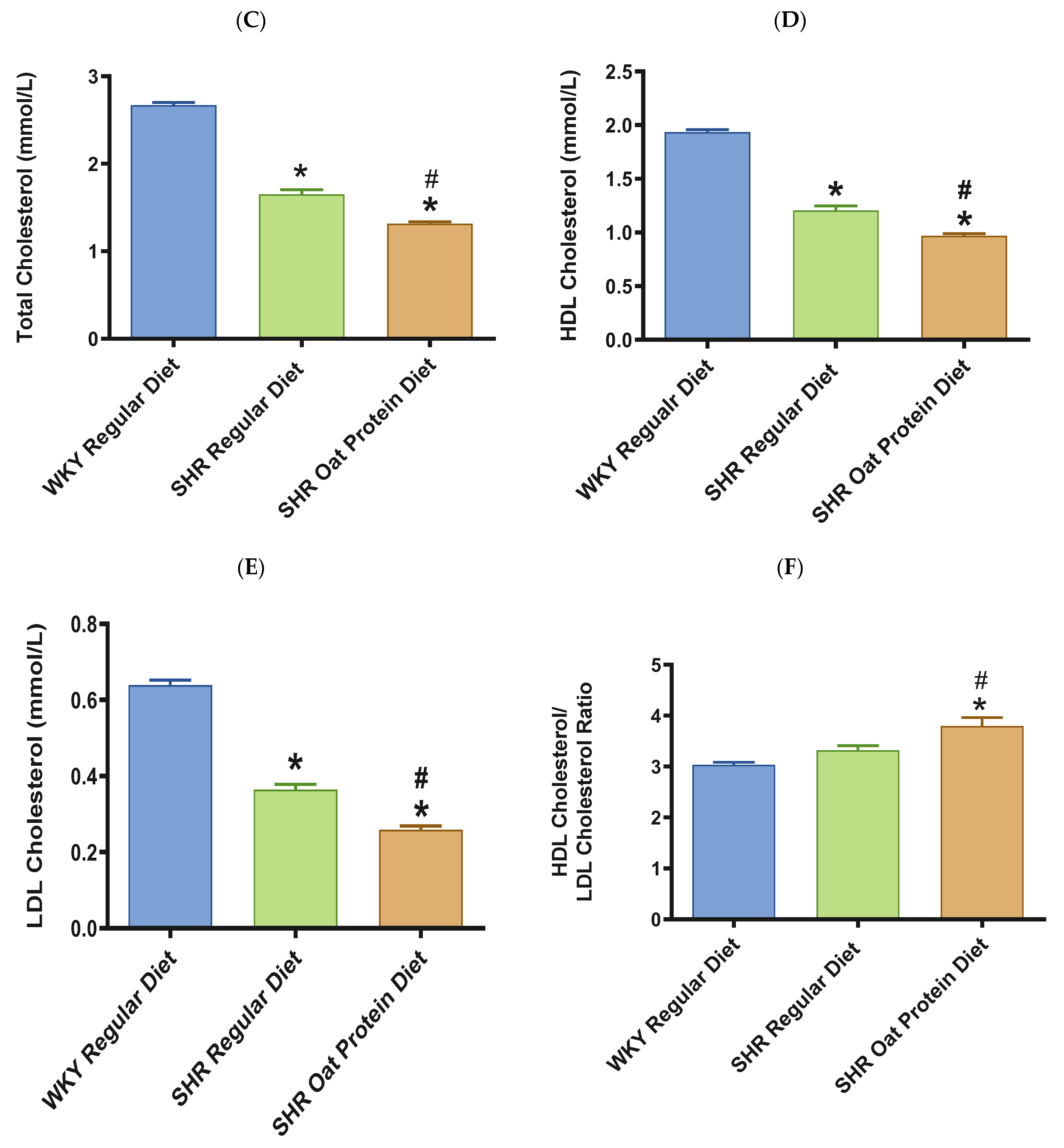
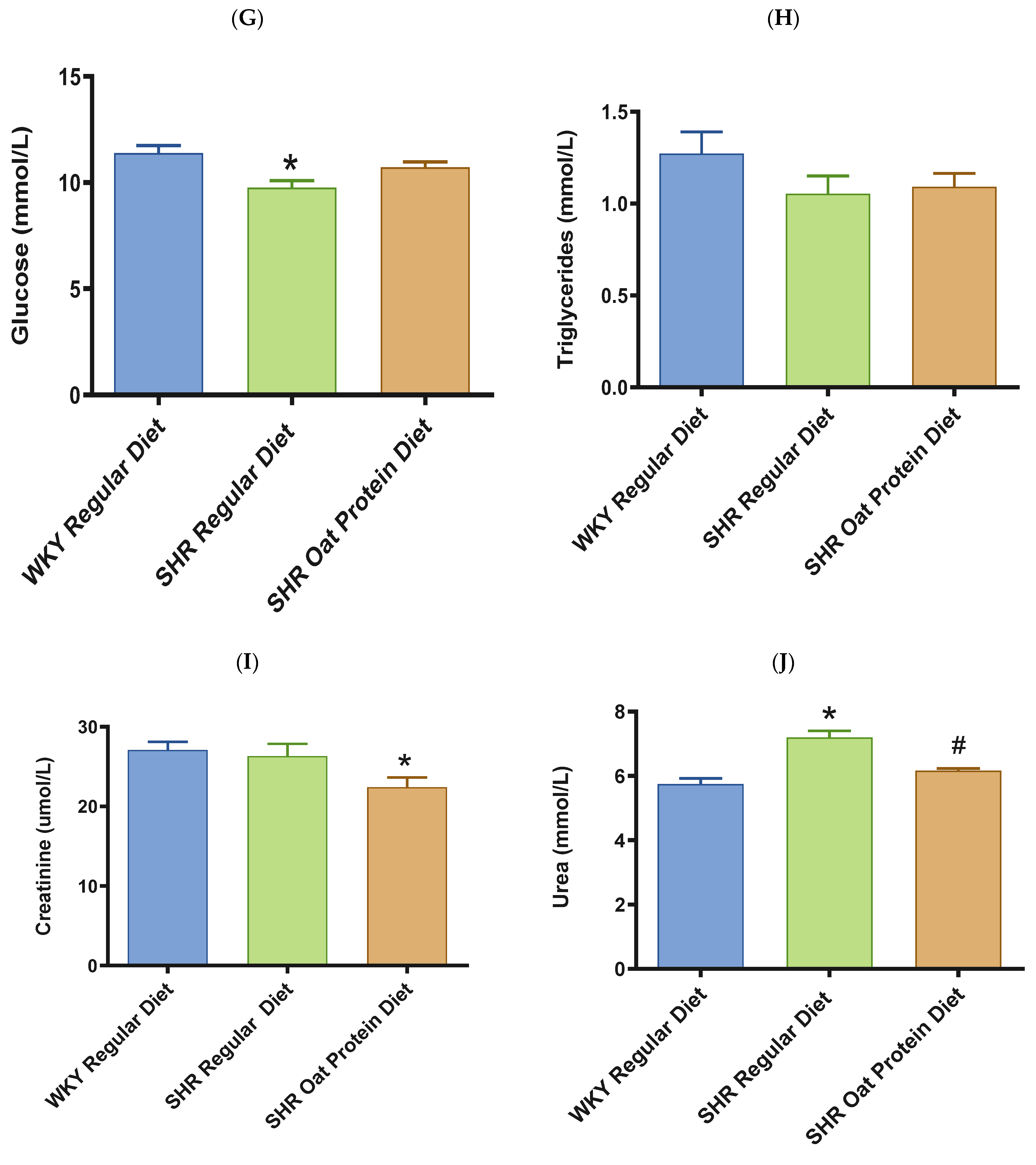
2.5. Biochemical Assessments
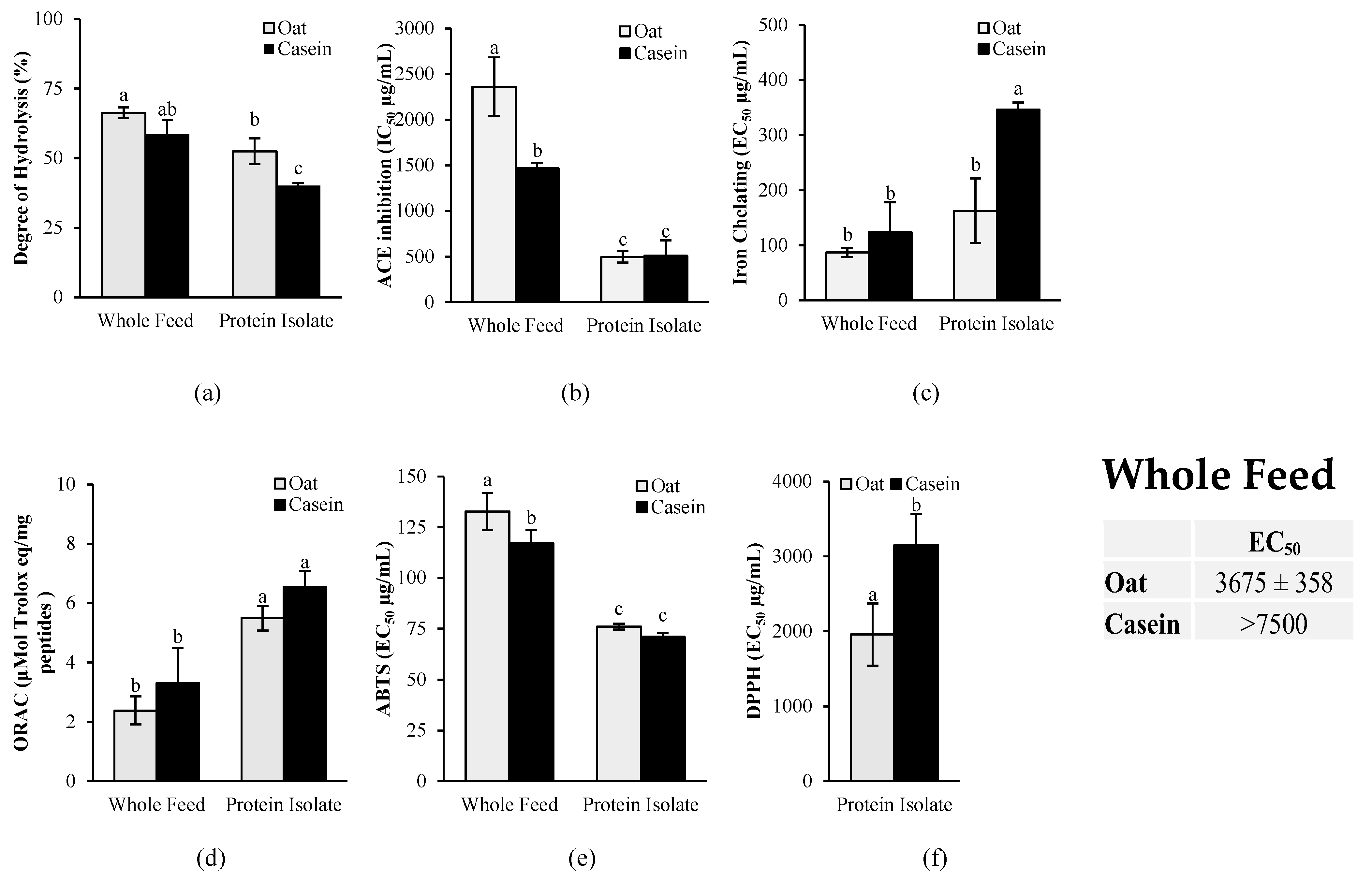
2.6. In Vitro Antioxidant and ACE Inhibition Activities of Oat Protein in Comparison to Casein After In Vitro Gastrointestinal Digestion
2.6.1. In Vitro Gastrointestinal Digestion Protocol
2.6.2. Assessment of Protein Degree of Hydrolysis
2.6.3. In Vitro ACE Inhibition Activity
2.6.4. In Vitro Antioxidant Activity
2.6.5. In Silico Prediction of Antihypertensive, Antioxidant, and Anti-Inflammatory Activities of Casein and Oat Proteins After Gastrointestinal Digestion
2.7. Statistical Analysis
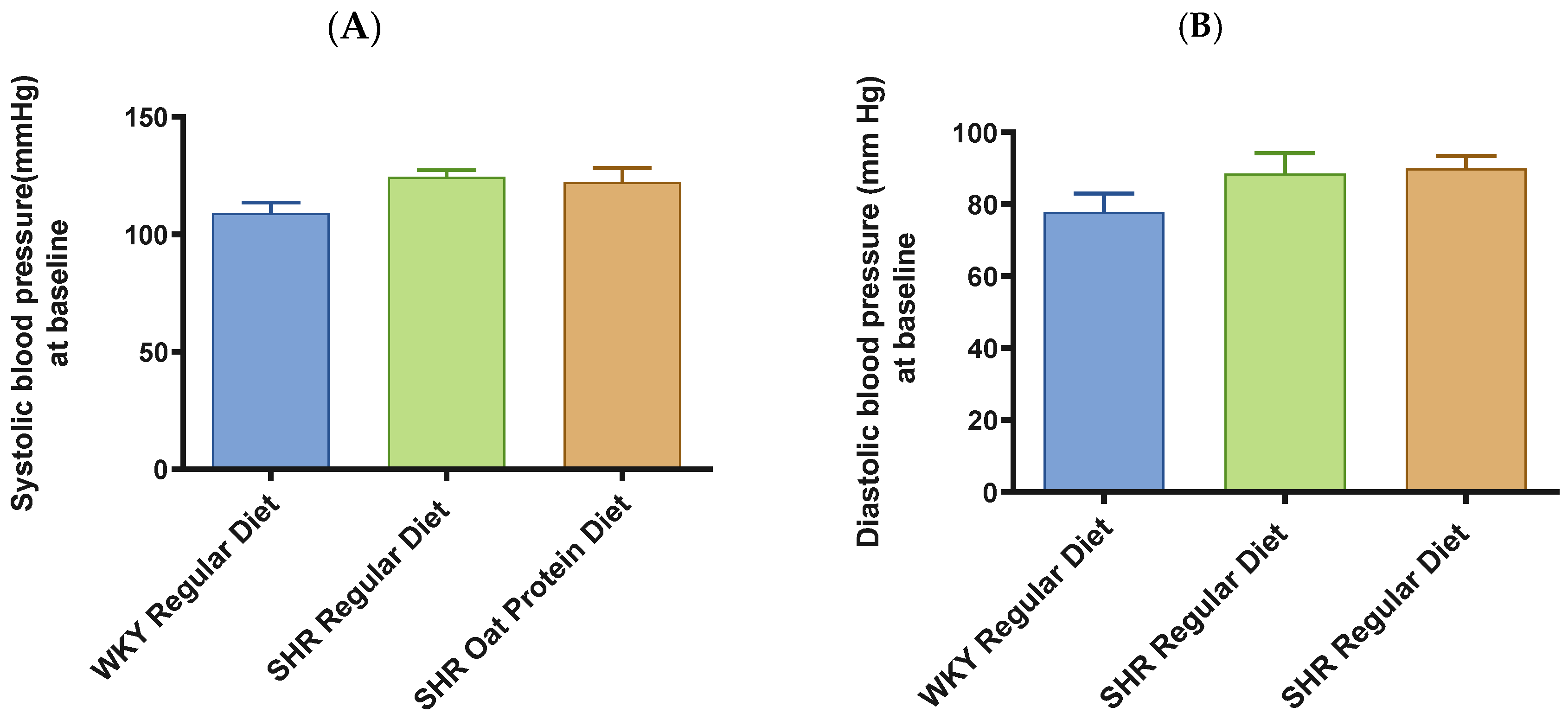
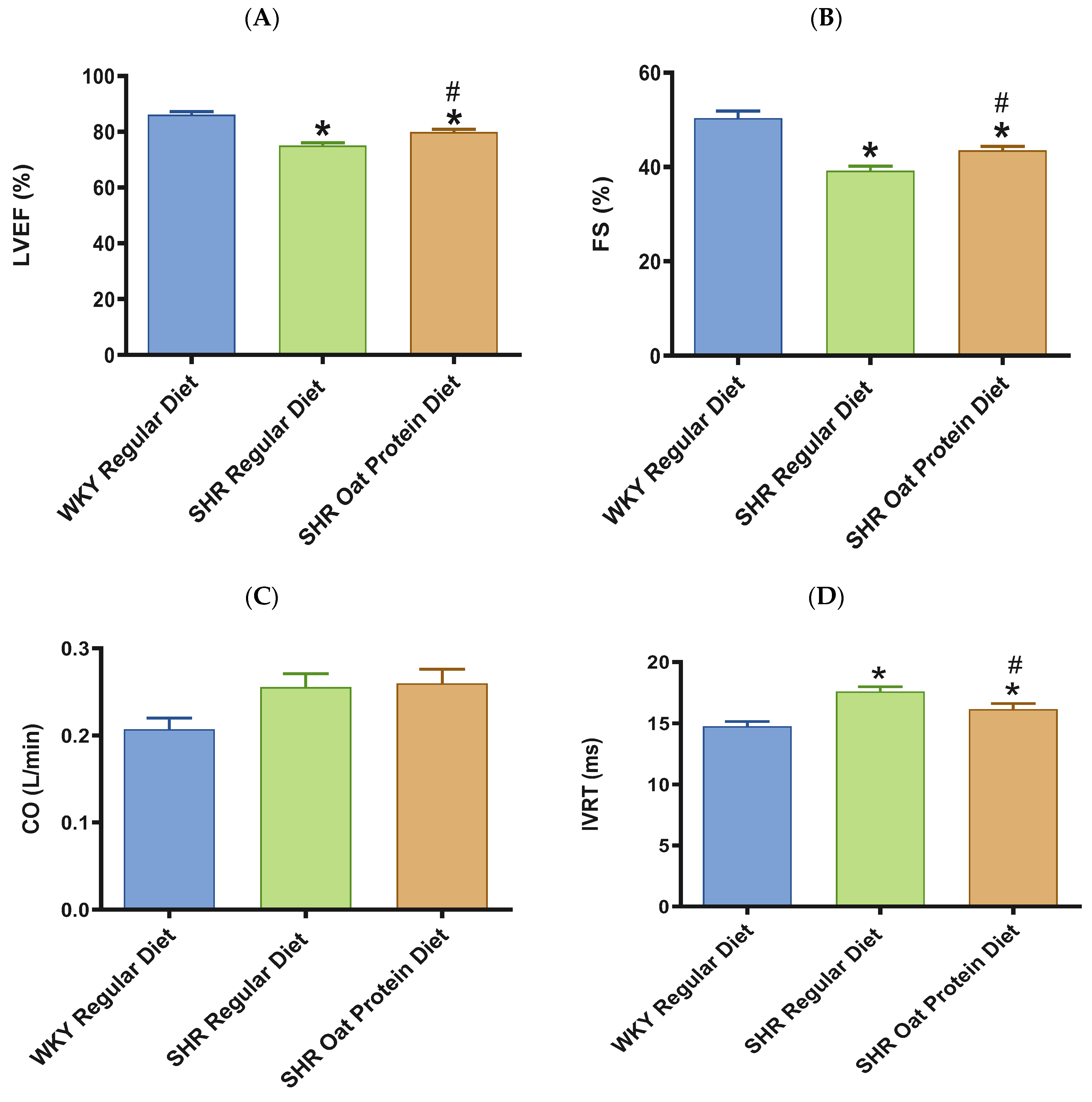
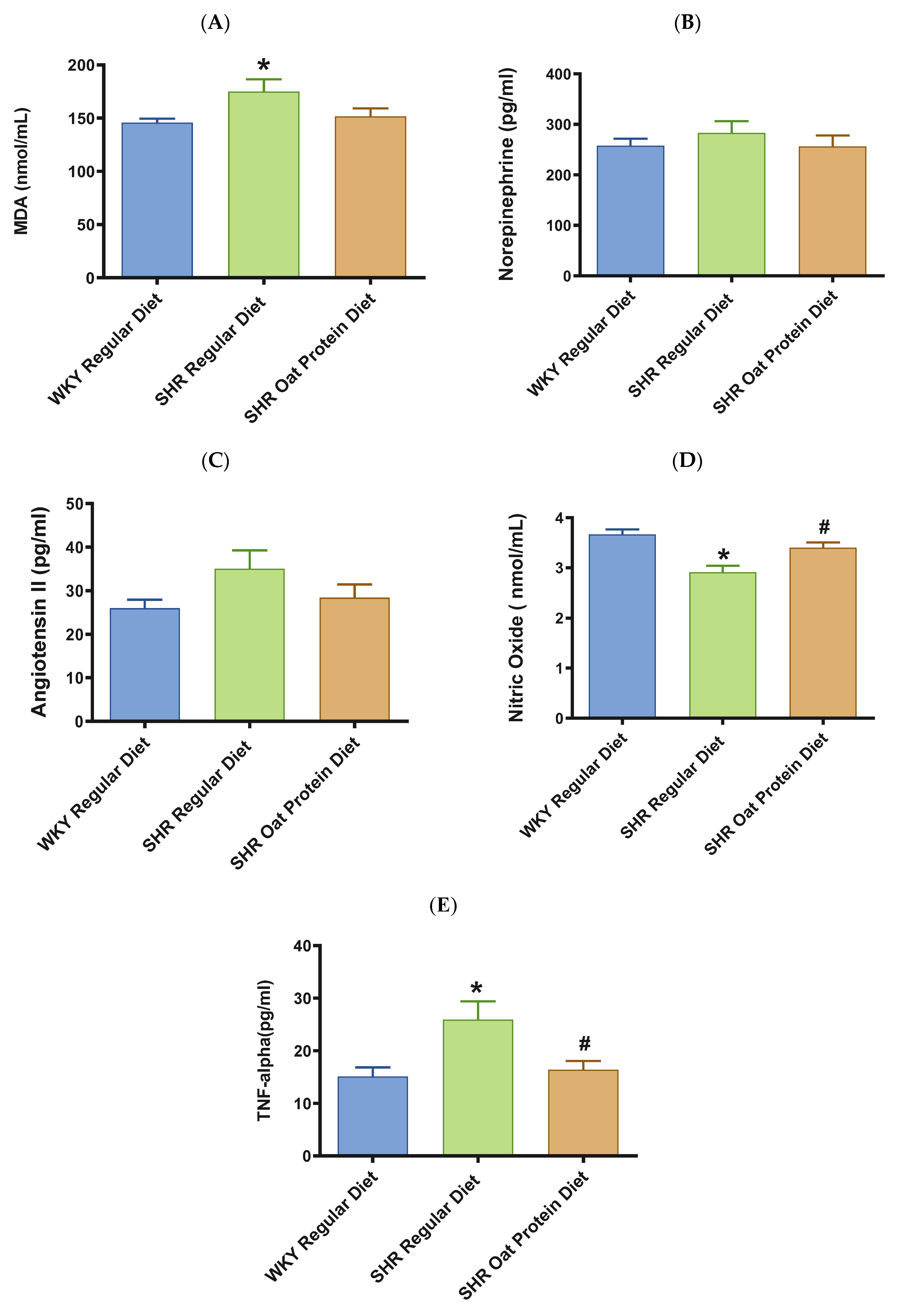
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, G.A.; Mensah, G.A.; Fuster, V. The Global Burden of Cardiovascular Diseases and Risks: A Compass for Global Action. J. Am. Coll. Cardiol. 2020, 76, 2980–2981. [Google Scholar] [CrossRef] [PubMed]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef]
- Bouchard, J.; Valookaran, A.F.; Aloud, B.M.; Raj, P.; Malunga, L.N.; Thandapilly, S.J.; Netticadan, T. Impact of oats in the prevention/management of hypertension. Food Chem. 2022, 381, 132198. [Google Scholar] [CrossRef]
- Valookaran, A.F.; Bouchard, J.; Aloud, B.M.; Thandapilly, S.J.; Netticadan, T. Therapeutic Potential of Select Dietary Compounds in the Management of Hypertension and its Cardiovascular Complications. Molecules 2022, 27, 7222. [Google Scholar] [CrossRef]
- Svetkey, L.P.; Simons-Morton, D.; Vollmer, W.M.; Appel, L.J.; Conlin, P.R.; Ryan, D.H.; Ard, J.; Kennedy, B.M. Effects of dietary patterns on blood pressure: Subgroup analysis of the Dietary Approaches to Stop Hypertension (DASH) randomized clinical trial. Arch. Intern. Med. 1999, 159, 285–293. [Google Scholar] [CrossRef]
- Alexander, S.; Ostfeld, R.J.; Allen, K.; Williams, K.A. A plant-based diet and hypertension. J. Geriatr. Cardiol. 2017, 14, 327–330. [Google Scholar]
- Lloyd-Jones, D.M.; Lewis, C.E.; Schreiner, P.J.; Shikany, J.M.; Sidney, S.; Reis, J.P. The Coronary Artery Risk Development In Young Adults (CARDIA) Study: JACC Focus Seminar 8/8. J. Am. Coll. Cardiol. 2021, 78, 260–277. [Google Scholar] [CrossRef] [PubMed]
- Elliott, P.; Stamler, J.; Dyer, A.R.; Appel, L.; Dennis, B.; Kesteloot, H.; Ueshima, H.; Okayama, A.; Chan, Q.; Garside, D.B.; et al. Association between protein intake and blood pressure: The INTERMAP Study. Arch. Intern. Med. 2006, 166, 79–87. [Google Scholar] [CrossRef]
- Stamler, J.; Liu, K.; Ruth, K.J.; Pryer, J.; Greenland, P. Eight-year blood pressure change in middle-aged men: Relationship to multiple nutrients. Hypertension 2002, 39, 1000–1006. [Google Scholar] [CrossRef]
- Shobako, N. Hypotensive peptides derived from plant proteins. Peptides 2021, 142, 170573. [Google Scholar] [CrossRef]
- Llanaj, E.; Dejanovic, G.M.; Valido, E.; Bano, A.; Gamba, M.; Kastrati, L.; Minder, B.; Stojic, S.; Voortman, T.; Marques-Vidal, P.; et al. Effect of oat supplementation interventions on cardiovascular disease risk markers: A systematic review and meta-analysis of randomized controlled trials. Eur. J. Nutr. 2022, 61, 1749–1778. [Google Scholar] [CrossRef] [PubMed]
- Rafique, H.; Dong, R.; Wang, X.; Alim, A.; Aadil, R.M.; Li, L.; Zou, L.; Hu, X. Dietary-Nutraceutical Properties of Oat Protein and Peptides. Front. Nutr. 2022, 9, 950400. [Google Scholar] [CrossRef]
- Boukid, F. Oat proteins as emerging ingredients for food formulation: Where we stand? Eur. Food Res. Technol. 2021, 247, 535–544. [Google Scholar] [CrossRef]
- Jing, X.; Yang, C.; Zhang, L. Characterization and Analysis of Protein Structures in Oat Bran. J. Food Sci. 2016, 81, C2337–C2343. [Google Scholar] [CrossRef] [PubMed]
- Georgiopoulos, G.; Kollia, Z.; Katsi, V.; Oikonomou, D.; Tsioufis, C.; Tousoulis, D. Nurse’s Contribution to Alleviate Non-adherence to Hypertension Treatment. Curr. Hypertens. Rep. 2018, 20, 65. [Google Scholar] [CrossRef]
- Esfandi, R.; Seidu, I.; Willmore, W.; Tsopmo, A. Antioxidant, pancreatic lipase, and alpha-amylase inhibitory properties of oat bran hydrolyzed proteins and peptides. J. Food Biochem. 2022, 46, e13762. [Google Scholar] [CrossRef]
- Wang, W.; Liu, X.; Li, Y.; You, H.; Yu, Z.; Wang, L.; Liu, X.; Ding, L. Identification and Characterization of Dipeptidyl Peptidase-IV Inhibitory Peptides from Oat Proteins. Foods 2022, 11, 1406. [Google Scholar] [CrossRef]
- Tong, L.T.; Guo, L.; Zhou, X.; Qiu, J.; Liu, L.; Zhong, K.; Zhou, S. Effects of dietary oat proteins on cholesterol metabolism of hypercholesterolaemic hamsters. J. Sci. Food Agric. 2016, 96, 1396–1401. [Google Scholar] [CrossRef]
- Guo, L.; Tong, L.T.; Liu, L.; Zhong, K.; Qiu, J.; Zhou, S. The cholesterol-lowering effects of oat varieties based on their difference in the composition of proteins and lipids. Lipids Health Dis. 2014, 13, 182. [Google Scholar] [CrossRef]
- Bouchard, J.; Raj, P.; Yu, L.; Sobhi, B.; Malalgoda, M.; Malunga, L.; Netticadan, T.; Joseph Thandapilly, S. Oat protein modulates cholesterol metabolism and improves cardiac systolic function in high fat, high sucrose fed rats. Appl. Physiol. Nutr. Metab. 2024, 49, 738–750. [Google Scholar] [CrossRef]
- Bing, O.H.; Conrad, C.H.; Boluyt, M.O.; Robinson, K.G.; Brooks, W.W. Studies of prevention, treatment and mechanisms of heart failure in the aging spontaneously hypertensive rat. Heart Fail. Rev. 2002, 7, 71–88. [Google Scholar] [CrossRef] [PubMed]
- Doggrell, S.A.; Brown, L. Rat models of hypertension, cardiac hypertrophy and failure. Cardiovasc. Res. 1998, 39, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Raj, P.; Ames, N.; Joseph Thandapilly, S.; Yu, L.; Netticadan, T. The effects of oat ingredients on blood pressure in spontaneously hypertensive rats. J. Food Biochem. 2020, 44, e13402. [Google Scholar] [CrossRef] [PubMed]
- Brodkorb, A.; Egger, L.; Alminger, M.; Alvito, P.; Assunção, R.; Ballance, S.; Bohn, T.; Bourlieu-Lacanal, C.; Boutrou, R.; Carrière, F.; et al. INFOGEST static in vitro simulation of gastrointestinal food digestion. Nat. Protoc. 2019, 14, 991–1014. [Google Scholar] [CrossRef]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; FitzGerald, R.J. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Barbana, C.; Boye, J.I. Angiotensin I-converting enzyme inhibitory properties of lentil protein hydrolysates: Determination of the kinetics of inhibition. Food Chem. 2011, 127, 94–101. [Google Scholar] [CrossRef]
- Martineau-Côté, D.; Achouri, A.; Wanasundara, J.; Karboune, S.; L’Hocine, L. Health Beneficial Bioactivities of Faba Bean Gastrointestinal (In Vitro) Digestate in Comparison to Soybean and Pea. Int. J. Mol. Sci. 2022, 23, 9210. [Google Scholar] [CrossRef]
- Orona-Tamayo, D.; Valverde, M.E.; Nieto-Rendón, B.; Paredes-López, O. Inhibitory activity of chia (Salvia hispanica L.) protein fractions against angiotensin I-converting enzyme and antioxidant capacity. LWT—Food Sci. Technol. 2015, 64, 236–242. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Tomer, D.P.; McLeman, L.D.; Ohmine, S.; Scherer, P.M.; Murray, B.K.; O’Neill, K.L. Comparison of the Total Oxyradical Scavenging Capacity and Oxygen Radical Absorbance Capacity Antioxidant Assays. J. Med. Food 2007, 10, 337–344. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2022, 51, D523–D531. [Google Scholar]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed]
- Aronow, W.S. Treatment of hypertensive emergencies. Ann. Transl. Med. 2017, 5, S5. [Google Scholar] [CrossRef] [PubMed]
- Appel, L.J.; Sacks, F.M.; Carey, V.J.; Obarzanek, E.; Swain, J.F.; Miller, E.R.; Conlin, P.R.; Erlinger, T.P.; Rosner, B.A.; Laranjo, N.M.; et al. Effects of protein, monounsaturated fat, and carbohydrate intake on blood pressure and serum lipids: Results of the OmniHeart randomized trial. JAMA 2005, 294, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Rouse, I.L.; Beilin, L.J.; Armstrong, B.K.; Vandongen, R. Blood-pressure-lowering effect of a vegetarian diet: Controlled trial in normotensive subjects. Lancet 1983, 1, 5–10. [Google Scholar] [CrossRef]
- Vasdev, S.; Stuckless, J. Antihypertensive effects of dietary protein and its mechanism. Int. J. Angiol. 2010, 19, e7–e20. [Google Scholar] [CrossRef]
- Darewicz, M.; Pliszka, M.; Borawska-Dziadkiewicz, J.; Minkiewicz, P.; Iwaniak, A. Multi-Bioactivity of Protein Digests and Peptides from Oat (Avena sativa L.) Kernels in the Prevention of the Cardiometabolic Syndrome. Molecules 2022, 27, 7907. [Google Scholar] [CrossRef]
- Bleakley, S.; Hayes, M.; O’Shea, N.; Gallagher, E.; Lafarga, T. Predicted Release and Analysis of Novel ACE-I, Renin, and DPP-IV Inhibitory Peptides from Common Oat (Avena sativa) Protein Hydrolysates Using In Silico Analysis. Foods 2017, 6, 108. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, X.; Zhuang, Y.; Li, Y.; Shi, P.; Tian, H.; Li, X.; Chen, X. Isolation of novel ACE-inhibitory peptide from naked oat globulin hydrolysates in silico approach: Molecular docking, in vivo antihypertension and effects on renin and intracellular endothelin-1. J. Food Sci. 2020, 85, 1328–1337. [Google Scholar] [CrossRef]
- Guo, H.; Hao, Y.; Fan, X.; Richel, A.; Everaert, N.; Yang, X.; Ren, G. Administration with Quinoa Protein Reduces the Blood Pressure in Spontaneously Hypertensive Rats and Modifies the Fecal Microbiota. Nutrients 2021, 13, 2446. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, Y.; Tian, T.; Xu, Y. Antihypertensive and antioxidant effects of food-derived bioactive peptides in spontaneously hypertensive rats. Food Sci. Nutr. 2024, 12, 8200–8210. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Zubcevic, J.; Vijay-Kumar, M.; Yang, T.; Manandhar, I.; Aryal, S.; Muralitharan, R.R.; Li, H.-B.; Li, Y.; Abais-Battad, J.M.; et al. Prospects for Leveraging the Microbiota as Medicine for Hypertension. Hypertension 2024, 81, 951–963. [Google Scholar] [CrossRef] [PubMed]
- Tong, L.T.; Xiao, T.; Wang, L.; Lu, C.; Liu, L.; Zhou, X.; Wang, A.; Qin, W.; Wang, F. Plant protein reduces serum cholesterol levels in hypercholesterolemia hamsters by modulating the compositions of gut microbiota and metabolites. iScience 2021, 24, 103435. [Google Scholar] [CrossRef]
- Zhen, J.; Zhou, Z.; He, M.; Han, H.X.; Lv, E.H.; Wen, P.B.; Liu, X.; Wang, Y.T.; Cai, X.C.; Tian, J.Q.; et al. The gut microbial metabolite trimethylamine N-oxide and cardiovascular diseases. Front. Endocrinol. 2023, 14, 1085041. [Google Scholar] [CrossRef]
- Crimarco, A.; Springfield, S.; Petlura, C.; Streaty, T.; Cunanan, K.; Lee, J.; Fielding-Singh, P.; Carter, M.M.; Topf, M.A.; Wastyk, H.C.; et al. A randomized crossover trial on the effect of plant-based compared with animal-based meat on trimethylamine-N-oxide and cardiovascular disease risk factors in generally healthy adults: Study with Appetizing Plantfood-Meat Eating Alternative Trial (SWAP-MEAT). Am. J. Clin. Nutr. 2020, 112, 1188–1199. [Google Scholar]
- Kim, H.M.; Hwang, I.C.; Choi, H.M.; Yoon, Y.E.; Cho, G.Y. Prognostic implication of left ventricular hypertrophy regression after antihypertensive therapy in patients with hypertension. Front. Cardiovasc. Med. 2022, 9, 1082008. [Google Scholar] [CrossRef]
- Kawasoe, S.; Ohishi, M. Regression of left ventricular hypertrophy. Hypertens. Res. 2024, 47, 1225–1226. [Google Scholar] [CrossRef]
- Verdecchia, P.; Schillaci, G.; Borgioni, C.; Ciucci, A.; Gattobigio, R.; Zampi, I.; Reboldi, G.; Porcellati, C. Prognostic significance of serial changes in left ventricular mass in essential hypertension. Circulation 1998, 97, 48–54. [Google Scholar] [CrossRef]
- Wong, M.; Staszewsky, L.; Latini, R.; Barlera, S.; Glazer, R.; Aknay, N.; Hester, A.; Anand, I.; Cohn, J.N. Severity of left ventricular remodeling defines outcomes and response to therapy in heart failure: Valsartan heart failure trial (Val-HeFT) echocardiographic data. J. Am. Coll. Cardiol. 2004, 43, 2022–2027. [Google Scholar] [CrossRef]
- Raj, P.; Sayfee, K.; Yu, L.; Sabra, A.; Wijekoon, C.; Malunga, L.; Thandapilly, S.J.; Netticadan, T. Oat Beta-Glucan Alone and in Combination with Hydrochlorothiazide Lowers High Blood Pressure in Male but Not Female Spontaneously Hypertensive Rats. Nutrients 2023, 15, 3180. [Google Scholar] [CrossRef]
- Mureddu, G.F.; de Simone, G.; Greco, R.; Rosato, G.F.; Contaldo, F. Left ventricular filling in arterial hypertension. Influence of obesity and hemodynamic and structural confounders. Hypertension 1997, 29, 544–550. [Google Scholar] [CrossRef] [PubMed]
- de Simone, G.; Greco, R.; Mureddu, G.; Romano, C.; Guida, R.; Celentano, A.; Contaldo, F. Relation of left ventricular diastolic properties to systolic function in arterial hypertension. Circulation 2000, 101, 152–157. [Google Scholar] [CrossRef]
- Mattioli, A.V.; Zennaro, M.; Bonatti, S.; Bonetti, L.; Mattioli, G. Regression of left ventricular hypertrophy and improvement of diastolic function in hypertensive patients treated with telmisartan. Int. J. Cardiol. 2004, 97, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Griendling, K.K.; Camargo, L.L.; Rios, F.J.; Alves-Lopes, R.; Montezano, A.C.; Touyz, R.M. Oxidative Stress and Hypertension. Circ. Res. 2021, 128, 993–1020. [Google Scholar] [CrossRef]
- Anderson, H.D.; Rahmutula, D.; Gardner, D.G. Tumor necrosis factor-alpha inhibits endothelial nitric-oxide synthase gene promoter activity in bovine aortic endothelial cells. J. Biol. Chem. 2004, 279, 963–969. [Google Scholar] [CrossRef]
- Forstermann, U.; Sessa, W.C. Nitric oxide synthases: Regulation and function. Eur. Heart J. 2012, 33, 829–837. [Google Scholar] [CrossRef]
- Tran, N.; Garcia, T.; Aniqa, M.; Ali, S.; Ally, A.; Nauli, S.M. Endothelial Nitric Oxide Synthase (eNOS) and the Cardiovascular System: In Physiology and in Disease States. Am. J. Biomed. Sci. Res. 2022, 15, 153–177. [Google Scholar] [PubMed]
- Li, Q.; Youn, J.Y.; Cai, H. Mechanisms and consequences of endothelial nitric oxide synthase dysfunction in hypertension. J. Hypertens. 2015, 33, 1128–1136. [Google Scholar] [CrossRef] [PubMed]
- van Loo, G.; Bertrand, M.J.M. Death by TNF: A road to inflammation. Nat. Rev. Immunol. 2023, 23, 289–303. [Google Scholar] [CrossRef]
- Snyder, E.C.; Abdelbary, M.; El-Marakby, A.; Sullivan, J.C. Treatment of male and female spontaneously hypertensive rats with TNF-alpha inhibitor etanercept increases markers of renal injury independent of an effect on blood pressure. Biol. Sex Differ. 2022, 13, 17. [Google Scholar] [CrossRef]
- Zhang, J.; Patel, J.M.; Li, Y.D.; Block, E.R. Proinflammatory cytokines downregulate gene expression and activity of constitutive nitric oxide synthase in porcine pulmonary artery endothelial cells. Res. Commun. Mol. Pathol. Pharmacol. 1997, 96, 71–87. [Google Scholar] [PubMed]
- Goodwin, B.L.; Pendleton, L.C.; Levy, M.M.; Solomonson, L.P.; Eichler, D.C. Tumor necrosis factor-alpha reduces argininosuccinate synthase expression and nitric oxide production in aortic endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H1115–H1121. [Google Scholar] [CrossRef] [PubMed]
- Xia, Z.; Liu, M.; Wu, Y.; Sharma, V.; Luo, T.; Ouyang, J.; McNeill, J.H. N-acetylcysteine attenuates TNF-alpha-induced human vascular endothelial cell apoptosis and restores eNOS expression. Eur. J. Pharmacol. 2006, 550, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Crorkin, P.; Hao, S.; Ferreri, N.R. Responses to Ang II (Angiotensin II), Salt Intake, and Lipopolysaccharide Reveal the Diverse Actions of TNF-alpha (Tumor Necrosis Factor-alpha) on Blood Pressure and Renal Function. Hypertension 2022, 79, 2656–2670. [Google Scholar] [CrossRef] [PubMed]
- Mehaffey, E.; Majid, D.S.A. Tumor necrosis factor-alpha, kidney function, and hypertension. Am. J. Physiol. Renal. Physiol. 2017, 313, F1005–F1008. [Google Scholar] [CrossRef]
- Yuzbashian, E.; Asghari, G.; Mirmiran, P.; Hosseini, F.S.; Azizi, F. Associations of dietary macronutrients with glomerular filtration rate and kidney dysfunction: Tehran lipid and glucose study. J. Nephrol. 2015, 28, 173–180. [Google Scholar] [CrossRef]
- Chen, X.; Wei, G.; Jalili, T.; Metos, J.; Giri, A.; Cho, M.E.; Boucher, R.; Greene, T.; Beddhu, S. The Associations of Plant Protein Intake With All-Cause Mortality in CKD. Am. J. Kidney Dis. 2016, 67, 423–430. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, L.; Wu, S.; Zhao, W.; Ding, L.; Liu, J. In Vivo anti-hypertensive effect of peptides from egg white and its molecular mechanism with ACE. Int. J. Food Sci. Technol. 2021, 56, 1030–1039. [Google Scholar] [CrossRef]
- Zhao, T.; Liu, B.; Yuan, L.; Sun, L.; Zhuang, Y. ACE inhibitory activity in vitro and antihypertensive effect in vivo of LSGYGP and its transepithelial transport by Caco-2 cell monolayer. J. Funct. Foods 2019, 61, 103488. [Google Scholar] [CrossRef]
- Gao, Q.; Xu, L.; Cai, J. New drug targets for hypertension: A literature review. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166037. [Google Scholar] [CrossRef]
- Rodrigo, R.; González, J.; Paoletto, F. The role of oxidative stress in the pathophysiology of hypertension. Hypertens. Res. 2011, 34, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Omosebi, M.O.; Osundahunsi, O.F.; Fagbemi, T.N. Effect of extrusion on protein quality, antinutritional factors, and digestibility of complementary diet from quality protein maize and soybean protein concentrate. J. Food Biochem. 2018, 42, e12508. [Google Scholar] [CrossRef]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef]
- Lund, M.N. Reactions of plant polyphenols in foods: Impact of molecular structure. Trends Food Sci. Technol. 2021, 112, 241–251. [Google Scholar] [CrossRef]
- Kutzli, I.; Weiss, J.; Gibis, M. Glycation of Plant Proteins Via Maillard Reaction: Reaction Chemistry, Technofunctional Properties, and Potential Food Application. Foods 2021, 10, 376. [Google Scholar] [CrossRef]
- Udenigwe, C.C.; Fogliano, V. Food matrix interaction and bioavailability of bioactive peptides: Two faces of the same coin? J. Funct. Foods 2017, 35, 9–12. [Google Scholar] [CrossRef]

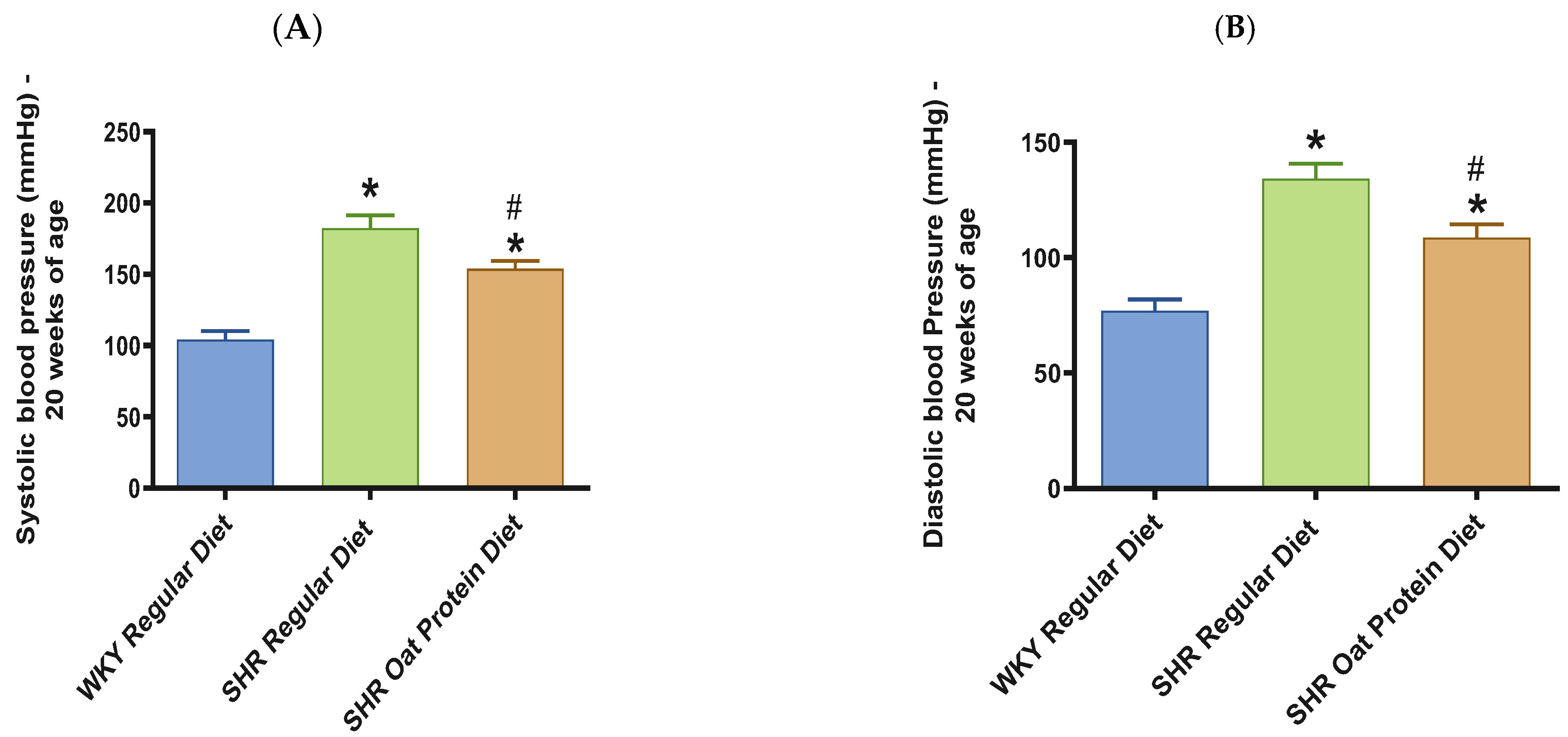


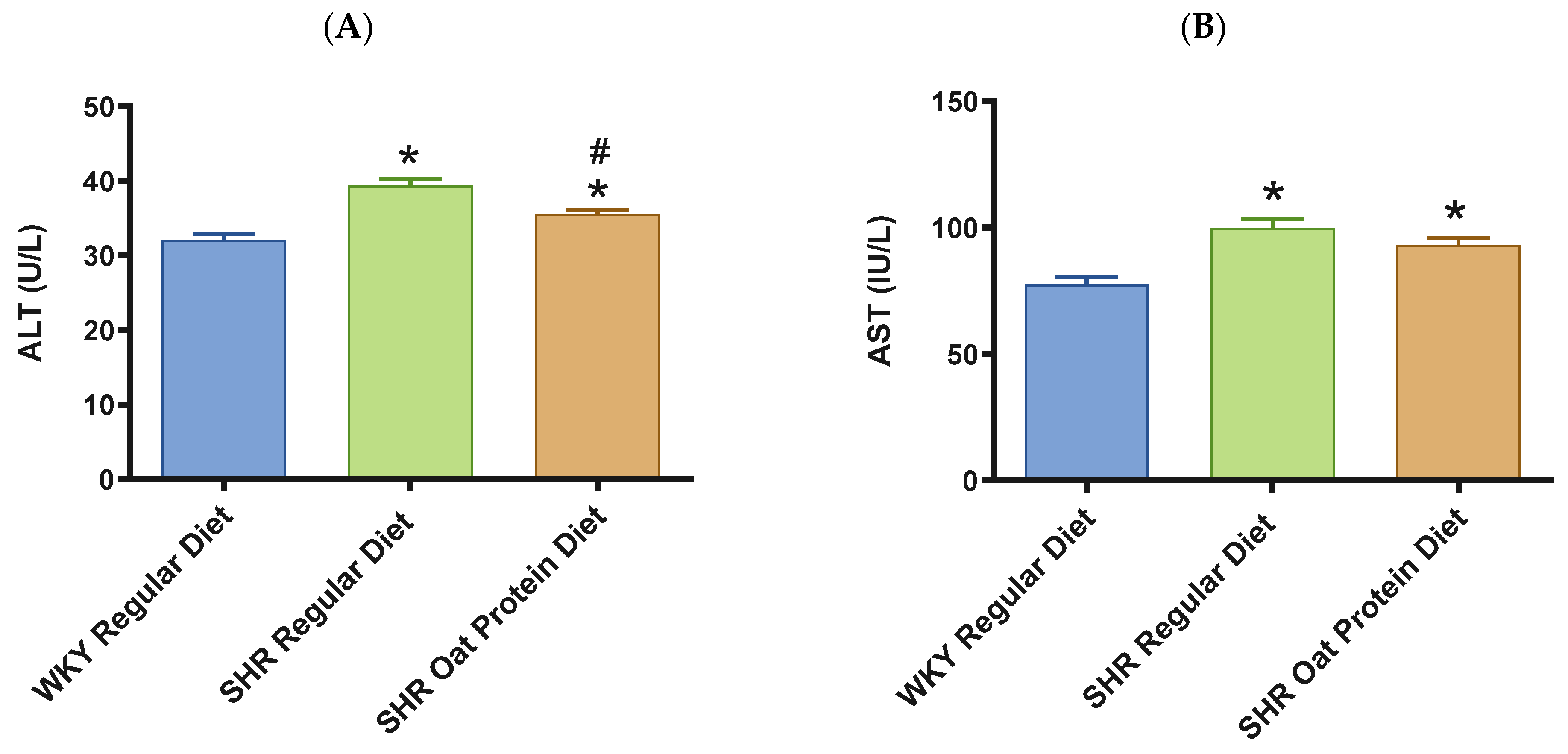

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raj, P.; Bouchard, J.; Martineau-Côté, D.; Malunga, L.; L’Hocine, L.; Yu, L.; Sobhi, B.; Achouri, A.; Pitre, M.; Thandapilly, S.J.; et al. Oat-Protein-Based Diet Lowers Blood Pressure and Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Rats. Nutrients 2024, 16, 3870. https://doi.org/10.3390/nu16223870
Raj P, Bouchard J, Martineau-Côté D, Malunga L, L’Hocine L, Yu L, Sobhi B, Achouri A, Pitre M, Thandapilly SJ, et al. Oat-Protein-Based Diet Lowers Blood Pressure and Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Rats. Nutrients. 2024; 16(22):3870. https://doi.org/10.3390/nu16223870
Chicago/Turabian StyleRaj, Pema, Jenny Bouchard, Delphine Martineau-Côté, Lovemore Malunga, Lamia L’Hocine, Liping Yu, Babak Sobhi, Allaoua Achouri, Mélanie Pitre, Sijo Joseph Thandapilly, and et al. 2024. "Oat-Protein-Based Diet Lowers Blood Pressure and Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Rats" Nutrients 16, no. 22: 3870. https://doi.org/10.3390/nu16223870
APA StyleRaj, P., Bouchard, J., Martineau-Côté, D., Malunga, L., L’Hocine, L., Yu, L., Sobhi, B., Achouri, A., Pitre, M., Thandapilly, S. J., & Netticadan, T. (2024). Oat-Protein-Based Diet Lowers Blood Pressure and Prevents Cardiac Remodeling and Dysfunction in Spontaneously Hypertensive Rats. Nutrients, 16(22), 3870. https://doi.org/10.3390/nu16223870