
Vitamin C Inhibits Lipopolysaccharide-Induced Hyperinflammatory State of Chronic Myeloid Leukemia Cells through Purinergic Signaling and Autophagy
 , ,
, ,  , , , , , ,
, , , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Human Cell Culture Study
2.2. Cytotoxicity Evaluation Using MTT Assay
2.3. Adenosine Triphosphate (ATP) Measurement
2.4. Cytokines Measurement
2.5. Reverse Transcriptase–Polymerase Chain Reaction (RT-PCR)
2.6. Protein Expression of Autophagy Biomarker LC3-I and LC3-II
2.7. Statistical Analysis
3. Results
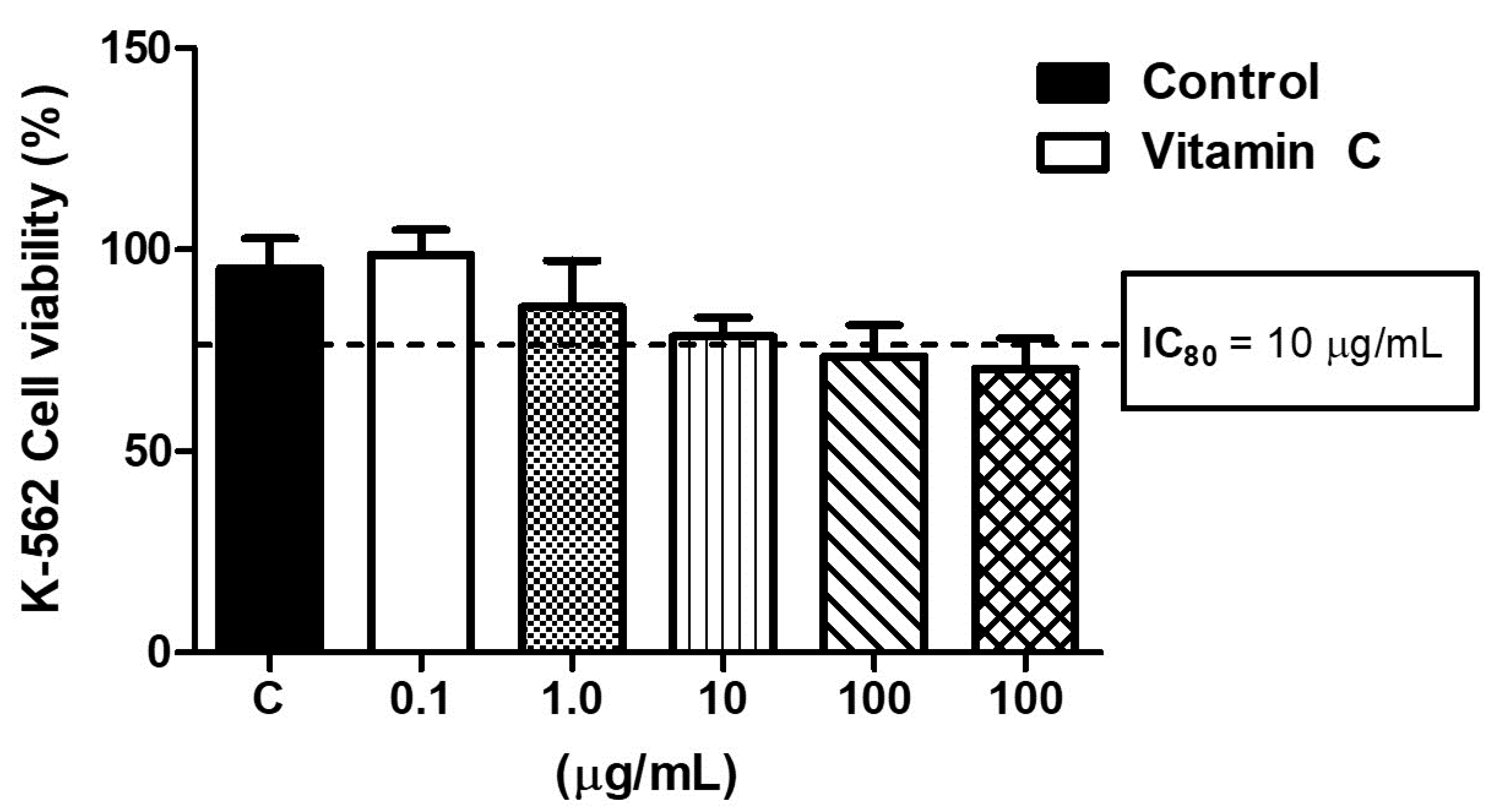
3.1. High Levels of Vitamin C Induced K-562 Cell Toxicity
3.2. Vitamin C Modulates Pro- and Anti-Inflammatory Cytokines in K-562 Cells
3.3. Vitamin C Modulates Components of the Purinergic Signaling Pathway in K-562 Cells
3.4. Vitamin C Reduces Autophagy Pathway in K-562 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, P.; Kumar, V.; Gupta, S.K.; Kumari, G.; Verma, M. Combating TKI resistance in CML by inhibiting the PI3K/Akt/mTOR pathway in combination with TKIs: A review. Med. Oncol. 2021, 38, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Cortes, J.; Kantarjian, H. Estimations of the increasing prevalence and plateau prevalence of chronic myeloid leukemia in the era of tyrosine kinase inhibitor therapy. Cancer 2012, 118, 3123–3127. [Google Scholar] [CrossRef] [PubMed]
- Rinaldi, I.; Winston, K. Chronic Myeloid Leukemia, from Pathophysiology to Treatment-Free Remission: A Narrative Literature Review. J. Blood Med. 2023, 14, 261–277. [Google Scholar] [CrossRef]
- Aldapt, M.B.; Al-Mashdali, A.F.; Obeidat, K.; Chandra, P.; Yassin, M. Viral Infections and Incidence of Reactivations in Chronic Myeloid Leukemia Patients. Oncology 2023. Online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Donadieu, J.; Leblanc, T.; Bader Meunier, B.; Barkaoui, M.; Fenneteau, O.; Bertrand, Y.; Maier-Redelsperger, M.; Micheau, M.; Stephan, J.L.; Phillipe, N.; et al. Analysis of risk factors for myelodysplasias, leukemias and death from infection among patients with congenital neutropenia. Experience of the French Severe Chronic Neutropenia Study Group. Haematologica 2005, 90, 45–53. [Google Scholar]
- Bestach, Y.; Toloza, M.J.; Ferri, C.; Larripa, I.; Belli, C. The dynamic of TNF and IL6 gene expression in chronic myeloid leukemia patients reveals early responders to imatinib. Leuk. Res. 2019, 86, 106221. [Google Scholar] [CrossRef]
- Ćojbašić, I.; Golubović, I.; Ćojbašić, Ž. Clinical Outcomes of Patients with Chronic Myeloid Leukemia and COVID-19 Infection—A Single Center Survey. Medicina 2023, 59, 1564. [Google Scholar] [CrossRef]
- Sun, L.-R.; Wang, L.-Z.; Zhong, R.; Zhao, Y.-X.; Sun, Y. Tyrosine Kinase Inhibitors for Pediatric Leukemia: History and Current Status. Discov. Med. 2022, 33, 93–99. [Google Scholar]
- Willig, J.B.; Vianna, D.R.B.; Beckenkamp, A.; Beckenkamp, L.R.; Sévigny, J.; Wink, M.R.; Buffon, A.; Pilger, D.A. Imatinib mesylate affects extracellular ATP catabolism and expression of NTPDases in a chronic myeloid leukemia cell line. Purinergic Signal. 2020, 16, 29–40. [Google Scholar] [CrossRef]
- Bella, Y.F.; Oliveira, C.R.; Mateus-Silva, J.R.; Brandao-Rangel, M.A.R.; Silva-Reis, A.; Santos, J.d.M.B.; Albertini, R.; Lopes-Martins, R.A.B.; de Oliveira, L.V.F.; Vieira, R.P. A phytotherapic blend immunity-6™ inhibits myeloid leukemic cells 2 activation involving purinergic signaling. Biomed. Pharmacother. 2023, 159, 114263. [Google Scholar] [CrossRef]
- Bellodi, C.; Lidonnici, M.R.; Hamilton, A.; Helgason, G.V.; Soliera, A.R.; Ronchetti, M.; Galavotti, S.; Young, K.W.; Selmi, T.; Yacobi, R.; et al. Targeting autophagy potentiates tyrosine kinase inhibitor–induced cell death in Philadelphia chromosome–positive cells, including primary CML stem cells. J. Clin. Investig. 2009, 119, 1109–1123. [Google Scholar] [CrossRef]
- Crowley, L.C.; Elzinga, B.M.; O'Sullivan, G.C.; McKenna, S.L. Autophagy induction by Bcr-Abl-expressing cells facilitates their recovery from a targeted or nontargeted treatment. Am. J. Hematol. 2010, 86, 38–47. [Google Scholar] [CrossRef]
- Liu, X.; Rothe, K.; Yen, R.; Fruhstorfer, C.; Maetzig, T.; Chen, M.; Forrest, D.L.; Humphries, R.K.; Jiang, X. A novel AHI-1–BCR-ABL–DNM2 complex regulates leukemic properties of primitive CML cells through enhanced cellular endocytosis and ROS-mediated autophagy. Leukemia 2017, 31, 2376–2387. [Google Scholar] [CrossRef]
- Yu, Y.; Yang, L.; Zhao, M.; Zhu, S.; Kang, R.; Vernon, P.; Tang, D.; Cao, L. Targeting microRNA-30a-mediated autophagy enhances imatinib activity against human chronic myeloid leukemia cells. Leukemia 2012, 26, 1752–1760. [Google Scholar] [CrossRef]
- Drullion, C.; Trégoat, C.; Lagarde, V.; Tan, S.; Gioia, R.; Priault, M.; Djavaheri-Mergny, M.; Brisson, A.; Auberger, P.; Mahon, F.-X.; et al. Apoptosis and autophagy have opposite roles on imatinib-induced K562 leukemia cell senescence. Cell Death Dis. 2012, 3, e373. [Google Scholar] [CrossRef] [PubMed]
- Agathocleous, M.; Meacham, C.E.; Burgess, R.J.; Piskounova, E.; Zhao, Z.; Crane, G.M.; Cowin, B.L.; Bruner, E.; Murphy, M.M.; Chen, W.; et al. Ascorbate regulates haematopoietic stem cell function and leukaemogenesis. Nature 2017, 549, 476–481. [Google Scholar] [CrossRef] [PubMed]
- Gillberg, L.; Ørskov, A.D.; Liu, M.; Harsløf, L.B.; Jones, P.A.; Grønbæk, K. Vitamin C—A new player in regulation of the cancer epigenome. Semin. Cancer Biol. 2018, 51, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Cimmino, L.; Dolgalev, I.; Wang, Y.; Yoshimi, A.; Martin, G.H.; Wang, J.; Ng, V.; Xia, B.; Witkowski, M.T.; Mitchell-Flack, M.; et al. Restoration of TET2 Function Blocks Aberrant Self-Renewal and Leukemia Progression. Cell 2017, 170, 1079–1095.e20. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.N.; Carr, A.C.; Antony, A.; Peng, S.; Fitzpatrick, M.G. Intravenous Vitamin C Administration Improved Blood Cell Counts and Health-Related Quality of Life of Patient with History of Relapsed Acute Myeloid Leukaemia. Antioxidants 2018, 7, 92. [Google Scholar] [CrossRef]
- Portugal, C.C.; da Encarnação, T.G.; Sagrillo, M.A.; Pereira, M.R.; Relvas, J.B.; Socodato, R.; Paes-De-Carvalho, R. Activation of adenosine A3 receptors regulates vitamin C transport and redox balance in neurons. Free Radic. Biol. Med. 2021, 163, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Karim, R.; Kadowaki, M. Effect and proposed mechanism of vitamin C modulating amino acid regulation of autophagic proteolysis. Biochimie 2017, 142, 51–62. [Google Scholar] [CrossRef]
- Rothe, K.; Porter, V.; Jiang, X. Current Outlook on Autophagy in Human Leukemia: Foe in Cancer Stem Cells and Drug Resistance, Friend in New Therapeutic Interventions. Int. J. Mol. Sci. 2019, 20, 461. [Google Scholar] [CrossRef]
- Barbosa, C.M.; Oliveira, C.R.; Nascimento, F.D.; Smith, M.C.; Fausto, D.M.; Soufen, M.A.; Sena, E.; Araújo, R.C.; Tersariol, I.L.; Bincoletto, C.; et al. Biphosphinic palladacycle complex mediates lysosomal-membrane permeabilization and cell death in K562 leukaemia cells. Eur. J. Pharmacol. 2006, 542, 37–47. [Google Scholar] [CrossRef]
- Garcia, M.; Santos-Dias, A.; Bachi, A.L.L.; Oliveira-Junior, M.C.; Andrade-Souza, A.S.; Ferreira, S.C.; Aquino-Junior, J.C.J.; Almeida, F.M.; Rigonato-Oliveira, N.C.; Oliveira, A.P.L.; et al. Creatine supplementation impairs airway inflammation in an experimental model of asthma involving P2 × 7 receptor. Eur. J. Immunol. 2019, 49, 928–939. [Google Scholar] [CrossRef] [PubMed]
- Alonso, S.; Minty, A.; Bourlet, Y.; Buckingham, M. Buckingham. Comparison of three actin-coding sequences in the mouse; evolutionary relationships between the actin genes of warm-blooded vertebrates. J. Mol. Evol. 1986, 23, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, D.; Idzko, M.; Dichmann, S.; Purlis, D.; Virchow, C.; Norgauer, J.; Chiozzi, P.; Di Virgilio, F.; Luttmann, W. P2 purinergic receptors of human eosinophils: Characterization and coupling to oxygen radical production. FEBS Lett. 2000, 486, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.G.; da Silva, C.A.A.; da Silva, A.; Vieira, R.P.; Mesquita-Ferrari, R.A.; Cogo, J.C.; Zamuner, S.R. Photobiomodulation Protects and Promotes Differentiation of C2C12 Myoblast Cells Exposed to Snake Venom. PLoS ONE 2016, 11, e0152890. [Google Scholar] [CrossRef]
- Wang, X.S.; Shi, Q.; Shah, N.D.; Heijnen, C.J.; Cohen, E.N.; Reuben, J.M.; Orlowski, R.Z.; Qazilbash, M.H.; Johnson, V.E.; Williams, L.A.; et al. Inflammatory Markers and Development of Symptom Burden in Patients with Multiple Myeloma during Autologous Stem Cell Transplantation. Clin. Cancer Res. 2014, 20, 1366–1374. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, M.; Dou, H.; Lin, W.; Zou, L. Sirtuin 1 inhibits lipopolysaccharide-induced inflammation in chronic myelogenous leukemia k562 cells through interacting with the Toll-like receptor 4-nuclear factor κ B-reactive oxygen species signaling axis. Cancer Cell Int. 2020, 20, 73. [Google Scholar] [CrossRef]
- Yao, X.; Huang, J.; Zhong, H.; Shen, N.; Faggioni, R.; Fung, M.; Yao, Y. Targeting interleukin-6 in inflammatory autoimmune diseases and cancers. Pharmacol. Ther. 2014, 141, 125–139. [Google Scholar] [CrossRef]
- Holbrook, J.; Lara-Reyna, S.; Jarosz-Griffiths, H.; McDermott, M.F. Tumour necrosis factor signalling in health and disease. F1000Research 2019, 8, F1000. [Google Scholar] [CrossRef] [PubMed]
- Iwase, S.; Nakada, T.-A.; Hattori, N.; Takahashi, W.; Takahashi, N.; Aizimu, T.; Yoshida, M.; Morizane, T.; Oda, S. Interleukin-6 as a diagnostic marker for infection in critically ill patients: A systematic review and meta-analysis. Am. J. Emerg. Med. 2018, 37, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Thompson, D.K.; Huffman, K.M.; Kraus, W.E.; Kraus, V.B. Critical Appraisal of Four IL-6 Immunoassays. PLoS ONE 2012, 7, e30659. [Google Scholar] [CrossRef] [PubMed]
- Aneja, R.; Carcillo, J. Differences between adult and pediatric septic shock. Minerva Anestesiol. 2011, 77, 986–992. [Google Scholar]
- Franco, D.M.; Arevalo-Rodriguez, I.; i Figuls, M.R.; Oleas, N.G.M.; Nuvials, X.; Zamora, J. Plasma interleukin-6 concentration for the diagnosis of sepsis in critically ill adults. Emergencias 2019, 4, CD011811. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, J.; He, H.; Ling, X.; Li, F.; Yang, Z.; Zhao, J.; Li, H.; Yang, T.; Zhao, S.; et al. Increased Cytokine Levels Assist in the Diagnosis of Respiratory Bacterial Infections or Concurrent Bacteremia in Patients With Non-Hodgkin’s Lymphoma. Front. Cell Infect. Microbiol. 2022, 12, 860526. [Google Scholar] [CrossRef]
- Pavlovsky, C.; Cordoba, B.V.; Sanchez, M.B.; Moiraghi, B.; Varela, A.; Custidiano, R.; Fernandez, I.; Freitas, M.J.; Ventriglia, M.V.; Bendek, G.; et al. Elevated plasma levels of IL-6 and MCP-1 selectively identify CML patients who better sustain molecular remission after TKI withdrawal. J. Hematol. Oncol. 2023, 16, 43. [Google Scholar] [CrossRef]
- Shen, N.; Liu, S.; Cui, J.; Li, Q.; You, Y.; Zhong, Z.; Cheng, F.; Guo, A.-Y.; Zou, P.; Yuan, G.; et al. Tumor necrosis factor α knockout impaired tumorigenesis in chronic myeloid leukemia cells partly by metabolism modification and miRNA regulation. OncoTargets Ther. 2019, 12, 2355–2364. [Google Scholar] [CrossRef] [PubMed]
- Balkwill, F. Tumour necrosis factor and cancer. Nat. Rev. Cancer 2009, 9, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The master regulator of immunity to infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Alberca-Custódio, R.W.; Greiffo, F.R.; MacKenzie, B.; Oliveira-Junior, M.C.; Andrade-Sousa, A.S.; Graudenz, G.S.; Santos, A.B.G.; Damaceno-Rodrigues, N.R.; Castro-Faria-Neto, H.C.; Arantes-Costa, F.M.; et al. Aerobic Exercise Reduces Asthma Phenotype by Modulation of the Leukotriene Pathway. Front. Immunol. 2016, 7, 237. [Google Scholar] [CrossRef]
- Mackenzie, B.; Andrade-Sousa, A.S.; Oliveira-Junior, M.C.; Assumpção-Neto, E.; Brandão-Rangel, M.A.R.; Silva-Renno, A.; Santos-Dias, A.; Cicko, S.; Grimm, M.; Müller, T.; et al. Dendritic Cells Are Involved in the Effects of Exercise in a Model of Asthma. Med. Sci. Sports Exerc. 2016, 48, 1459–1467. [Google Scholar] [CrossRef]
- Fitch, B.A.; Zhou, M.; Situ, J.; Surianarayanan, S.; Reeves, M.Q.; Hermiston, M.L.; Wiemels, J.L.; Kogan, S.C. Decreased IL-10 accelerates B-cell leukemia/lymphoma in a mouse model of pediatric lymphoid leukemia. Blood Adv. 2022, 6, 854–865. [Google Scholar] [CrossRef]
- Sanchez-Correa, B.; Bergua, J.M.; Campos, C.; Gayoso, I.; Arcos, M.J.; Bañas, H.; Morgado, S.; Casado, J.G.; Solana, R.; Tarazona, R. Cytokine profiles in acute myeloid leukemia patients at diagnosis: Survival is inversely correlated with IL-6 and directly correlated with IL-10 levels. Cytokine 2013, 61, 885–891. [Google Scholar] [CrossRef]
- Sachdeva, A.; Gouge, J.; Kontovounisios, C.; Nikolaou, S.; Ashworth, A.; Lim, K.; Chong, I. Klotho and the Treatment of Human Malignancies. Cancers 2020, 12, 1665. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Wu, W.; He, Y.; Ma, S.; Gao, J. The role of klotho in chronic kidney disease. BMC Nephrol. 2018, 19, 285. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, M. Emerging role of α-Klotho in energy metabolism and cardiometabolic diseases. Diabetes Metab. Syndr. Clin. Res. Rev. 2023, 17, 102854. [Google Scholar] [CrossRef] [PubMed]
- Prud’homme, G.J.; Kurt, M.; Wang, Q. Pathobiology of the Klotho Antiaging Protein and Therapeutic Considerations. Front. Aging 2022, 3, 931331. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Wang, Y.; Xiong, Y.; Lin, X.; Zhou, P.; Chen, Z. Reduced Klotho expression contributes to poor survival rates in human patients with ovarian cancer, and overexpression of Klotho inhibits the progression of ovarian cancer partly via the inhibition of systemic inflammation in nude mice. Mol. Med. Rep. 2017, 15, 1777–1785. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Oliveira, A.R.; Aquino-Junior, J.; Abbasi, A.; Santos-Dias, A.; Oliveira-Junior, M.C.; Alberca-Custodio, R.W.; Rigonato-Oliveira, N.C.; Salles-Dias, L.P.; Damaceno-Rodrigues, N.R.; Caldini, E.; et al. Effects of aerobic exercise on molecular aspects of asthma: Involvement of SOCS-JAK-STAT. Exerc. Immunol. Rev. 2019, 25, 50–62. [Google Scholar]
- Idzko, M.; Ferrari, D.; Eltzschig, H.K. Nucleotide signalling during inflammation. Nature 2014, 509, 310–317. [Google Scholar] [CrossRef]
- Schmid, S.; Kübler, M.; Ayata, C.K.; Lazar, Z.; Haager, B.; Hoßfeld, M.; Meyer, A.; Cicko, S.; Elze, M.; Wiesemann, S.; et al. Altered purinergic signaling in the tumor associated immunologic microenvironment in metastasized non-small-cell lung cancer. Lung Cancer 2015, 90, 516–521. [Google Scholar] [CrossRef]
- Drill, M.; Jones, N.C.; Hunn, M.; O’brien, T.J.; Monif, M. Antagonism of the ATP-gated P2X7 receptor: A potential therapeutic strategy for cancer. Purinergic Signal. 2021, 17, 215–227. [Google Scholar] [CrossRef]
- Shieh, C.; Heinrich, A.; Serchov, T.; van Calker, D.; Biber, K. P2X7-dependent, but differentially regulated release of IL-6, CCL2, and TNF-α in cultured mouse microglia. Glia 2014, 62, 592–607. [Google Scholar] [CrossRef]
- Larrouyet-Sarto, M.L.; Tamura, A.S.; Alves, V.S.; Santana, P.T.; Ciarlini-Magalhães, R.; Rangel, T.P.; Siebert, C.; Hartwig, J.R.; dos Santos, T.M.; Wyse, A.T.; et al. P2X7 receptor deletion attenuates oxidative stress and liver damage in sepsis. Purinergic Signal. 2020, 16, 561–572. [Google Scholar] [CrossRef]
- Sáez, P.J.; Vargas, P.; Shoji, K.F.; Harcha, P.A.; Lennon-Duménil, A.-M.; Sáez, J.C. ATP promotes the fast migration of dendritic cells through the activity of pannexin 1 channels and P2X7 receptors. Sci. Signal. 2017, 10, eaah7107. [Google Scholar] [CrossRef]
- Tarumoto, T.; Nagai, T.; Ohmine, K.; Miyoshi, T.; Nakamura, M.; Kondo, T.; Mitsugi, K.; Nakano, S.; Muroi, K.; Komatsu, N.; et al. Ascorbic acid restores sensitivity to imatinib via suppression of Nrf2-dependent gene expression in the imatinib-resistant cell line. Exp. Hematol. 2004, 32, 375–381. [Google Scholar] [CrossRef]
- Sharifzadeh, S.; Lak, E.R.; Tamaddon, G.; Ramzi, M.; Ranjbaran, R.; Abedi, E. The mRNA Expression of PTEN, LEF1, JAK3, LC3 and p62/SQSTM1 Genes in Patients with Chronic Myeloid Leukemia. Cardiovasc. Hematol. Agents Med. Chem. 2023, 21, 67–76. [Google Scholar] [CrossRef]
- Li, M.-Q.; Liu, Z.-G. Dual role of autophagy in chronic myeloid leukemia. Zhongguo Shi Yan Xue Ye Xue Za Zhi 2015, 23, 583–586. (In Chinese) [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pires, D.A.; Brandão-Rangel, M.A.R.; Silva-Reis, A.; Olímpio, F.R.S.; Aimbire, F.; Oliveira, C.R.; Mateus-Silva, J.R.; Zamarioli, L.S.; Bachi, A.L.L.; Bella, Y.F.; et al. Vitamin C Inhibits Lipopolysaccharide-Induced Hyperinflammatory State of Chronic Myeloid Leukemia Cells through Purinergic Signaling and Autophagy. Nutrients 2024, 16, 383. https://doi.org/10.3390/nu16030383
Pires DA, Brandão-Rangel MAR, Silva-Reis A, Olímpio FRS, Aimbire F, Oliveira CR, Mateus-Silva JR, Zamarioli LS, Bachi ALL, Bella YF, et al. Vitamin C Inhibits Lipopolysaccharide-Induced Hyperinflammatory State of Chronic Myeloid Leukemia Cells through Purinergic Signaling and Autophagy. Nutrients. 2024; 16(3):383. https://doi.org/10.3390/nu16030383
Chicago/Turabian StylePires, Daniela A., Maysa A. R. Brandão-Rangel, Anamei Silva-Reis, Fabiana R. S. Olímpio, Flavio Aimbire, Carlos R. Oliveira, José R. Mateus-Silva, Lucas S. Zamarioli, André L. L. Bachi, Yanesko F. Bella, and et al. 2024. "Vitamin C Inhibits Lipopolysaccharide-Induced Hyperinflammatory State of Chronic Myeloid Leukemia Cells through Purinergic Signaling and Autophagy" Nutrients 16, no. 3: 383. https://doi.org/10.3390/nu16030383
APA StylePires, D. A., Brandão-Rangel, M. A. R., Silva-Reis, A., Olímpio, F. R. S., Aimbire, F., Oliveira, C. R., Mateus-Silva, J. R., Zamarioli, L. S., Bachi, A. L. L., Bella, Y. F., Santos, J. M. B., Bincoletto, C., Lancha, A. H., Jr., & Vieira, R. P. (2024). Vitamin C Inhibits Lipopolysaccharide-Induced Hyperinflammatory State of Chronic Myeloid Leukemia Cells through Purinergic Signaling and Autophagy. Nutrients, 16(3), 383. https://doi.org/10.3390/nu16030383