

Erigeron annuus Extract Improves DNCB-Induced Atopic Dermatitis in a Mouse Model via the Nrf2/HO-1 Pathway

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Erigeron annuus Extracts (EAE)
2.2. Cell Culture
2.3. Cell Viability
2.4. In Vitro Antioxidant Activity
2.5. Establishment of the H2O2-Induced Oxidative Stress Model
2.6. Protective Effect of EAE on HaCaT Cells by H2O2-Induced Oxidative Stress
2.7. Analysis of MDA and SOD Contents in HaCaT Cells
2.8. Measurement of ROS Production
2.9. Animals
2.10. Sub-Acute Toxicity (28-Day Repeat Dose Toxicity)
2.11. Body Weight and Organ Weight Measurement and Serum Biochemistry Analysis
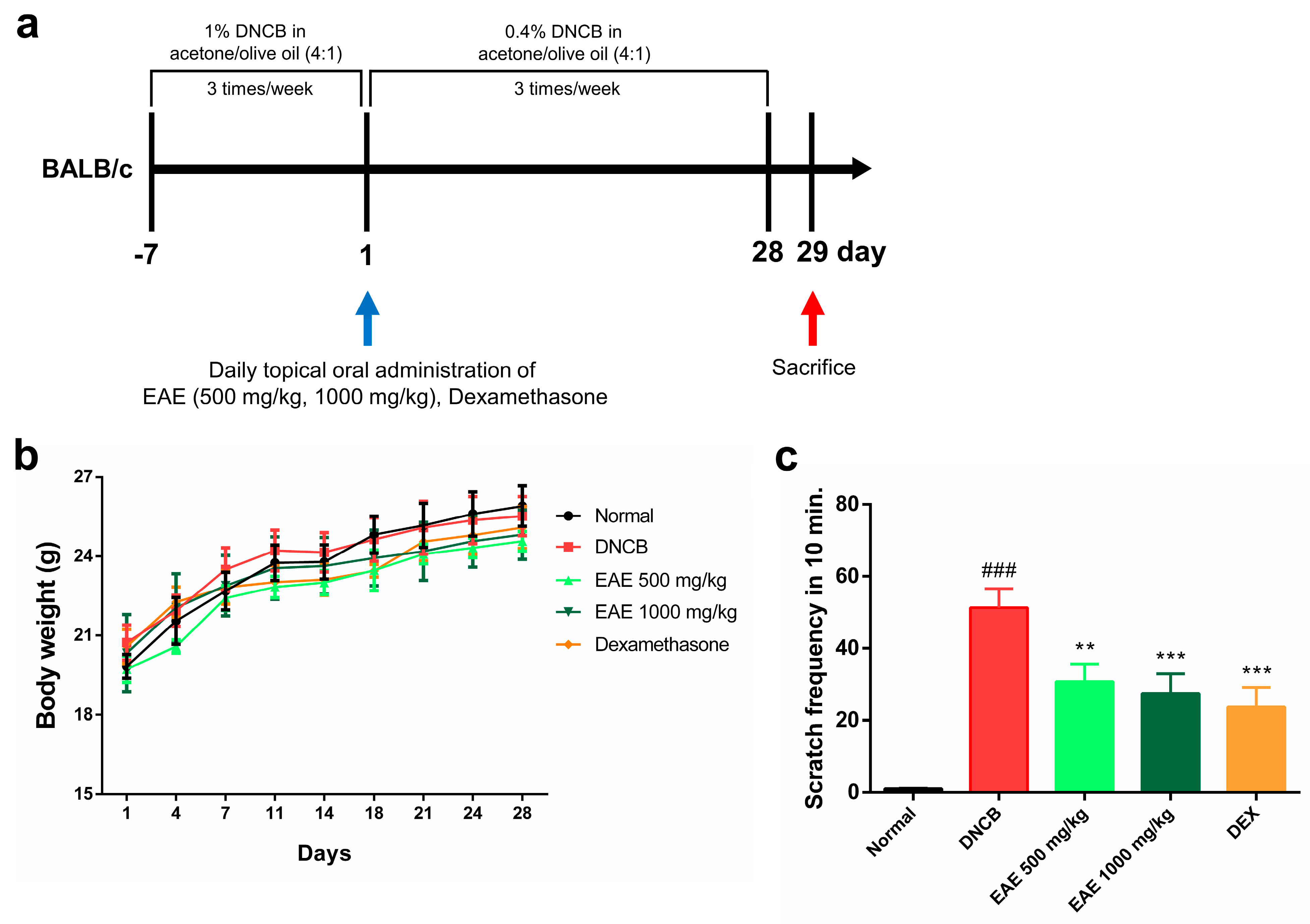
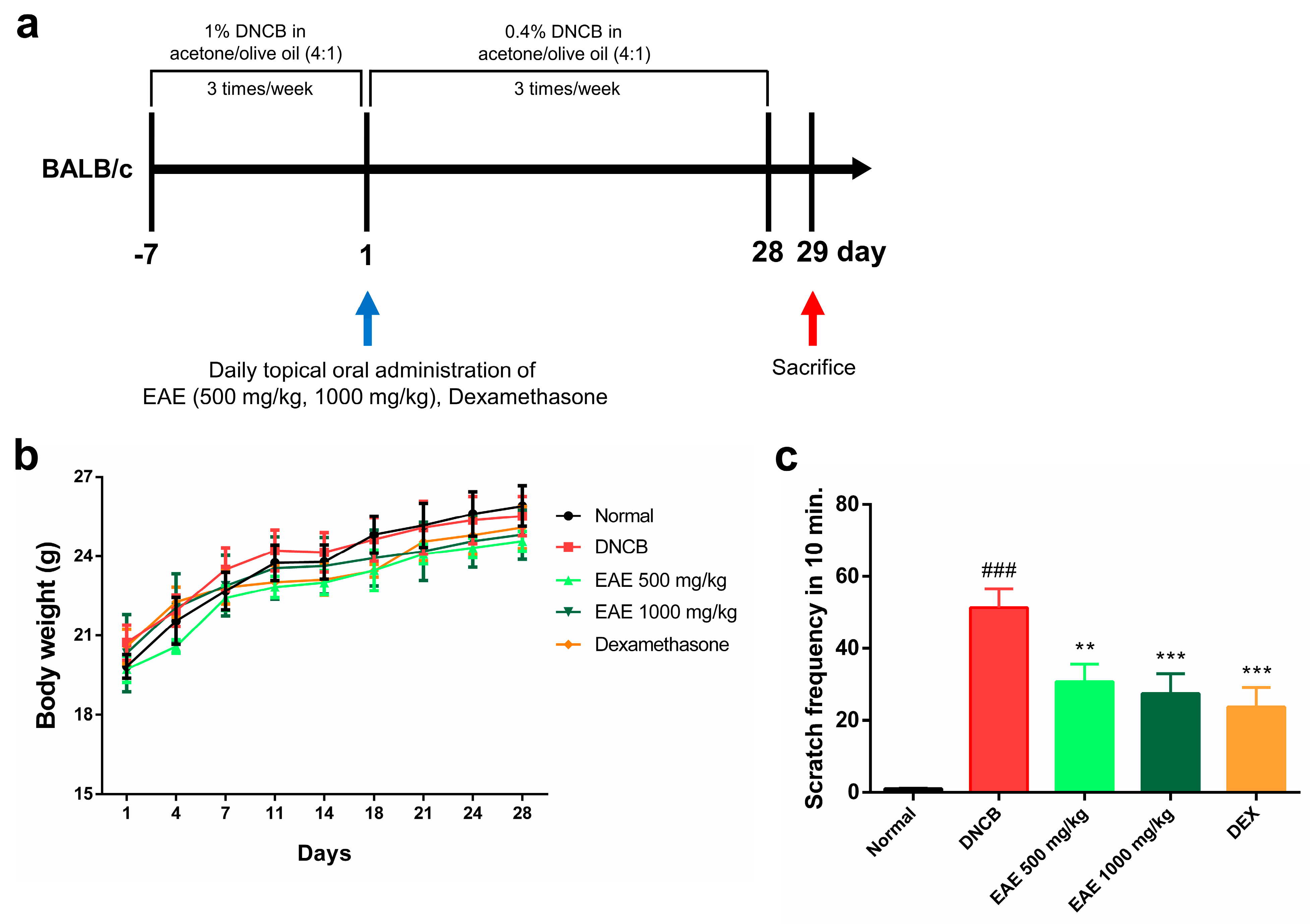
2.12. Atopic Dermatitis Model Induction and Treatment
2.13. Measures Body Weight and Scratching Frequency
2.14. Histological Analysis
2.15. Western Blotting
2.16. Statistical Analysis
3. Results
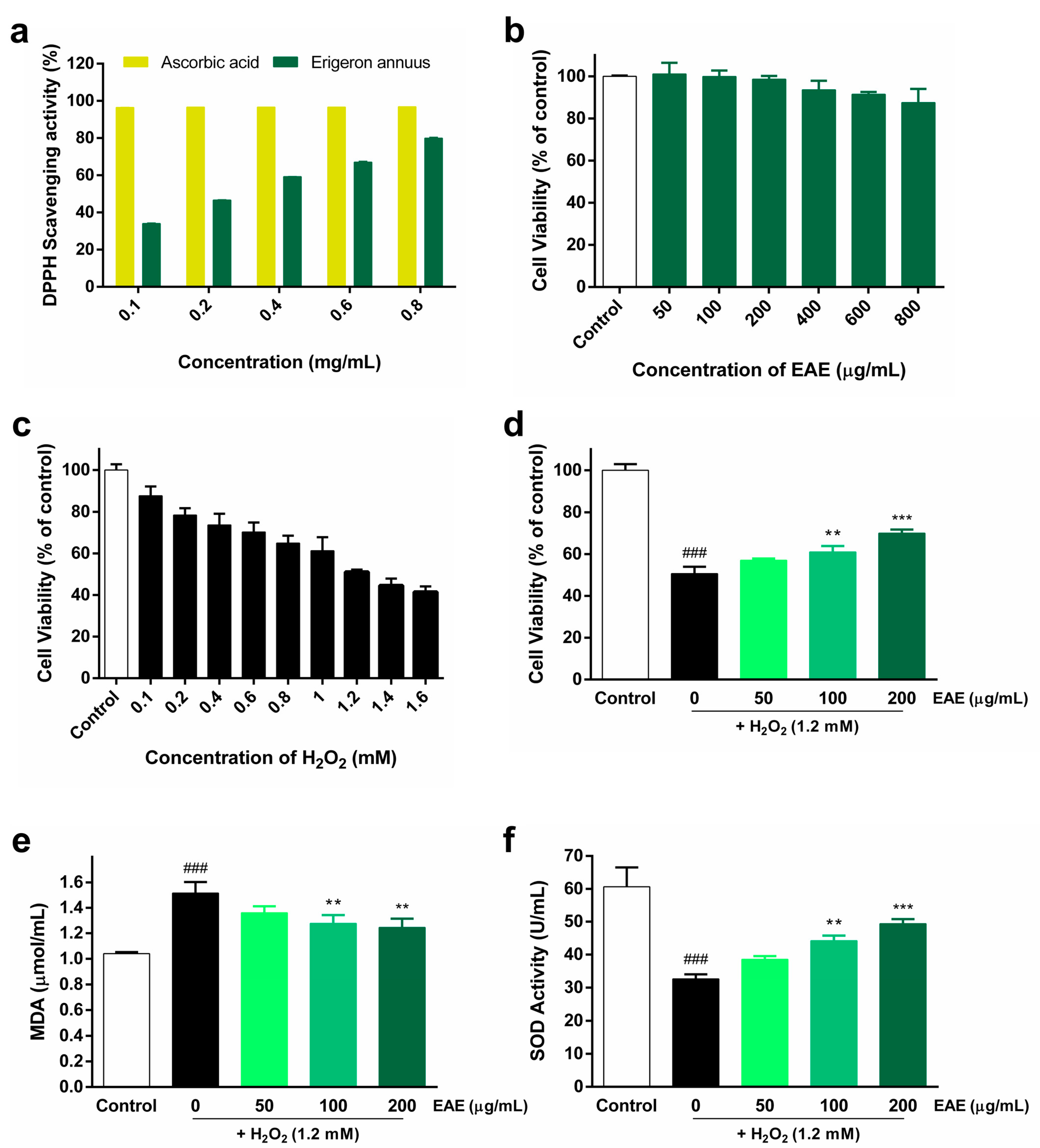
3.1. Antioxidant Activity of EAE against H2O2-Induced Oxidative Stress
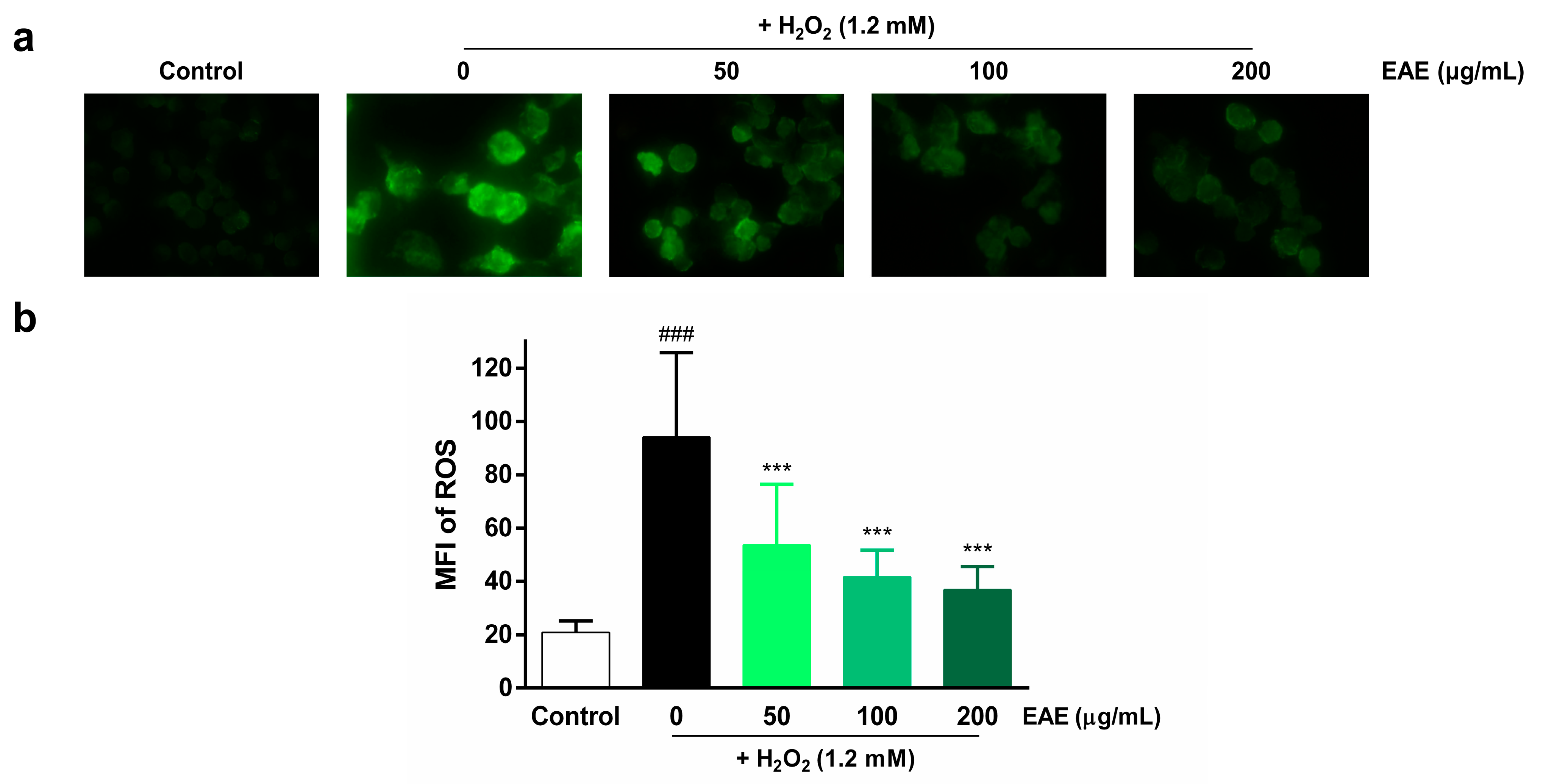
3.2. Protective Effect of EAE on H2O2-Induced ROS Production in HaCaT Cells
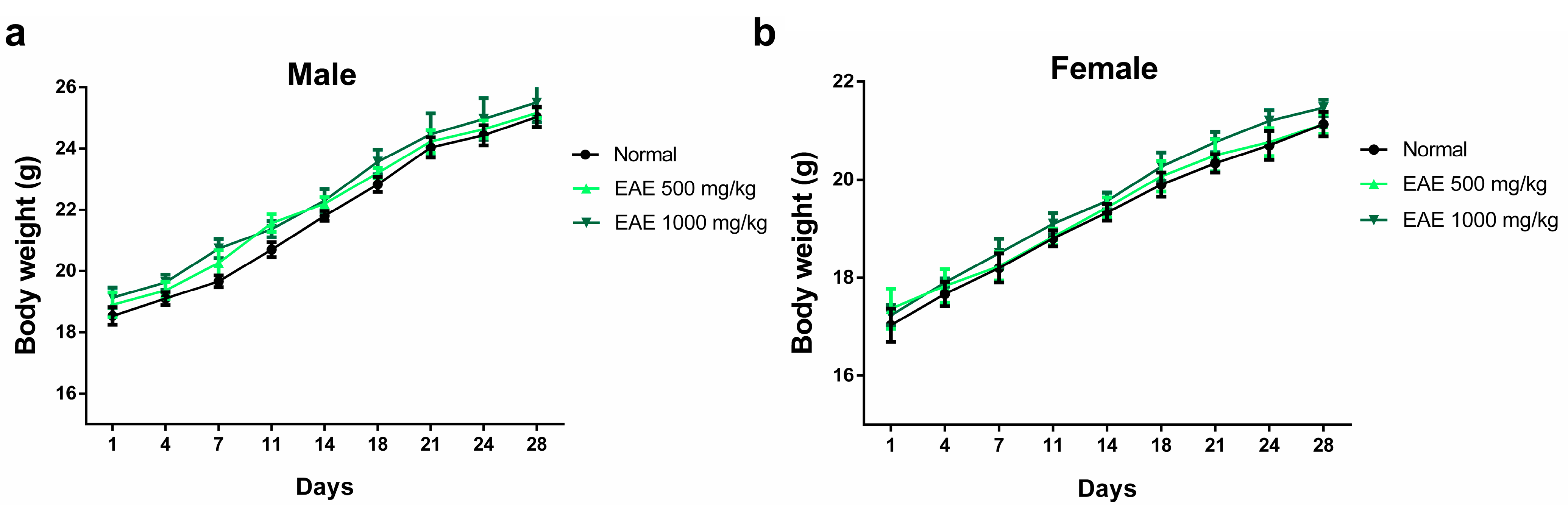
3.3. Subacute Oral Toxicity Studies
3.3.1. Assessment of Body Weight and Major Organ Weights
3.3.2. Hematological Parameters
3.3.3. Biochemical Parameters
3.4. Effect of Oral Administration of EAE on Mice
3.5. Histologic Analysis of AD Disease Model Induced by DNCB and the Impact of EAE on the Nrf2/HO-1 Signaling Pathway
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaufman, B.P.; Guttman-Yassky, E.; Alexis, A.F. Atopic dermatitis in diverse racial and ethnic groups—Variations in epidemiology, genetics, clinical presentation and treatment. Exp. Dermatol. 2018, 27, 340–357. [Google Scholar] [CrossRef]
- Peng, W.; Novak, N. Pathogenesis of atopic dermatitis. Clin. Exp. Allergy 2015, 45, 566–574. [Google Scholar] [CrossRef]
- Kim, B.E.; Leung, D.Y. Epidermal barrier in atopic dermatitis. Allergy Asthma Immunol. Res. 2012, 4, 12–16. [Google Scholar] [CrossRef] [PubMed]
- Lowe, A.J.; Leung, D.Y.; Tang, M.L.; Su, J.C.; Allen, K.J. The skin as a target for prevention of the atopic march. Ann. Allergy Asthma Immunol. 2018, 120, 145–151. [Google Scholar] [CrossRef] [PubMed]
- David Boothe, W.; Tarbox, J.A.; Tarbox, M.B. Atopic dermatitis: Pathophysiology. In Management of Atopic Dermatitis: Methods and Challenges; Springer: Berlin/Heidelberg, Germany, 2017; pp. 21–37. [Google Scholar]
- Kim, Y.E.; Choi, S.W.; Kim, M.K.; Nguyen, T.L.; Kim, J. Therapeutic hydrogel patch to treat atopic dermatitis by regulating oxidative stress. Nano Lett. 2022, 22, 2038–2047. [Google Scholar] [CrossRef] [PubMed]
- Paller, A.; Jaworski, J.C.; Simpson, E.L.; Boguniewicz, M.; Russell, J.J.; Block, J.K.; Tofte, S.; Dunn, J.D.; Feldman, S.R.; Clark, A.R. Major comorbidities of atopic dermatitis: Beyond allergic disorders. Am. J. Clin. Dermatol. 2018, 19, 821–838. [Google Scholar] [CrossRef] [PubMed]
- Chrostowska-Plak, D.; Salomon, J.; Reich, A.; Szepietowski, J.C. Clinical aspects of itch in adult atopic dermatitis patients. Acta Derm.-Venereol. 2009, 89, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.X.; Zug, K.A. Diagnosis and management of dermatitis, including atopic, contact, and hand eczemas. Med. Clin. 2021, 105, 611–626. [Google Scholar] [CrossRef] [PubMed]
- Lewis-Jones, S. Quality of life and childhood atopic dermatitis: The misery of living with childhood eczema. Int. J. Clin. Pract. 2006, 60, 984–992. [Google Scholar] [CrossRef]
- Grob, J.; Revuz, J.; Ortonne, J.; Auquier, P.; Lorette, G. Comparative study of the impact of chronic urticaria, psoriasis and atopic dermatitis on the quality of life. Br. J. Dermatol. 2005, 152, 289–295. [Google Scholar] [CrossRef]
- Lifschitz, C. The impact of atopic dermatitis on quality of life. Ann. Nutr. Metab. 2015, 66, 34–40. [Google Scholar] [CrossRef]
- Hong, J.; Buddenkotte, J.; Berger, T.G.; Steinhoff, M. Management of itch in atopic dermatitis. In Seminars in Cutaneous Medicine and Surgery; NIH Public Access: Bethesda, MD, USA, 2011; p. 71. [Google Scholar]
- Leung, D.Y.; Boguniewicz, M.; Howell, M.D.; Nomura, I.; Hamid, Q.A. New insights into atopic dermatitis. J. Clin. Investig. 2004, 113, 651–657. [Google Scholar] [CrossRef]
- DaVeiga, S.P. Epidemiology of atopic dermatitis: A review. Allergy Asthma Proc. 2012, 33, 227–234. [Google Scholar] [CrossRef]
- Berke, R.; Singh, A.; Guralnick, M. Atopic dermatitis: An overview. Am. Fam. Physician 2012, 86, 35–42. [Google Scholar]
- Fukaya, M.; Sato, K.; Sato, M.; Kimata, H.; Fujisawa, S.; Dozono, H.; Yoshizawa, J.; Minaguchi, S. Topical steroid addiction in atopic dermatitis. In Drug, Healthcare and Patient Safety; Taylor & Francis: New York, NY, USA, 2014; pp. 131–138. [Google Scholar]
- Eichenfield, L.F.; Tom, W.L.; Berger, T.G.; Krol, A.; Paller, A.S.; Schwarzenberger, K.; Bergman, J.N.; Chamlin, S.L.; Cohen, D.E.; Cooper, K.D. Guidelines of care for the management of atopic dermatitis: Section 2. Management and treatment of atopic dermatitis with topical therapies. J. Am. Acad. Dermatol. 2014, 71, 116–132. [Google Scholar] [CrossRef] [PubMed]
- Tempark, T.; Phatarakijnirund, V.; Chatproedprai, S.; Watcharasindhu, S.; Supornsilchai, V.; Wananukul, S. Exogenous Cushing’s syndrome due to topical corticosteroid application: Case report and review literature. Endocrine 2010, 38, 328–334. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Ma, Q.; Ye, L.; Piao, G. The traditional medicine and modern medicine from natural products. Molecules 2016, 21, 559. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Global Report on Traditional and Complementary Medicine 2019; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Shanker, C.; Tyagi, L.K.; Singh, M.; Rao, C.V. Herbal medicine for market potential in India: An overview. Acad. J. Plant Sci. 2008, 1, 26–36. [Google Scholar]
- Khan, M.S.A.; Ahmad, I.; Chattopadhyay, D. New Look to Phytomedicine: Advancements in Herbal Products as Novel Drug Leads; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Bedi, O.; Bijjem, K.R.V.; Kumar, P.; Gauttam, V. Herbal Induced Hepatoprotection and Hepatotoxicity: A Critical Review. Indian J. Physiol. Pharmacol. 2016, 60, 6–21. [Google Scholar] [PubMed]
- Azam, K.; Nur, M.; Rahman, M.M.; Biswas, S.; Ahmed, M.N. Appraisals of Bangladeshi medicinal plants used by folk medicine practitioners in the prevention and management of malignant neoplastic diseases. Int. Sch. Res. Not. 2016, 2016, 7832120. [Google Scholar]
- Greenwell, M.; Rahman, P. Medicinal plants: Their use in anticancer treatment. Int. J. Pharm. Sci. Res. 2015, 6, 4103. [Google Scholar] [PubMed]
- Mechesso, A.F.; Lee, S.-J.; Park, N.-H.; Kim, J.-Y.; Im, Z.-E.; Suh, J.-W.; Park, S.-C. Preventive effects of a novel herbal mixture on atopic dermatitis-like skin lesions in BALB/C mice. BMC Complement. Altern. Med. 2019, 19, 25. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-Y.; Kim, Y.J.; Jegal, J.; Jo, B.-G.; Choi, H.-S.; Yang, M.H. Haplopine Ameliorates 2, 4-Dinitrochlorobenzene-induced atopic dermatitis-like skin lesions in mice and TNF-α/IFN-γ-induced inflammation in human keratinocyte. Antioxidants 2021, 10, 806. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.-Y.; Park, N.-J.; Jegal, H.; Paik, J.-H.; Choi, S.; Kim, S.-N.; Yang, M.H. Nymphoides peltata Root Extracts Improve Atopic Dermatitis by Regulating Skin Inflammatory and Anti-Oxidative Enzymes in 2, 4-Dinitrochlorobenzene (DNCB)-Induced SKH-1 Hairless Mice. Antioxidants 2023, 12, 873. [Google Scholar] [CrossRef]
- Kang, B.; Shim, S. Overall status of naturalized plants in Korea. Korean J. Weed Sci. 2002, 22, 207–226. [Google Scholar]
- Jeong, C.-H.; Jeong, H.R.; Choi, G.N.; Kim, D.-O.; Lee, U.; Heo, H.J. Neuroprotective and anti-oxidant effects of caffeic acid isolated from Erigeron annuus leaf. Chin. Med. 2011, 6, 25. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, J.-Y.; Kim, D.H.; Kim, H.D.; Ji, Y.-J.; Seo, K.H. Erigeron annuus protects PC12 neuronal cells from oxidative stress induced by ROS-mediated apoptosis. Evid.-Based Complement. Altern. Med. 2020, 2020, 3945194. [Google Scholar] [CrossRef]
- Jo, M.J.; Lee, J.R.; Cho, I.J.; Kim, Y.W.; Kim, S.C. Roots of Erigeron annuus Attenuate Acute Inflammation as Mediated with the Inhibition of NF-κB-Associated Nitric Oxide and Prostaglandin E 2 production. Evid.-Based Complement. Altern. Med. 2013, 2013, 297427. [Google Scholar] [CrossRef]
- Liu, D.; Pan, F.; Liu, J.; Wang, Y.; Zhang, T.; Wang, E.; Liu, J. Individual and combined antioxidant effects of ginsenoside F2 and cyanidin-3-O-glucoside in human embryonic kidney 293 cells. RSC Adv. 2016, 6, 81092–81100. [Google Scholar] [CrossRef]
- Sies, H. On the history of oxidative stress: Concept and some aspects of current development. Curr. Opin. Toxicol. 2018, 7, 122–126. [Google Scholar] [CrossRef]
- Toyokuni, S. Reactive oxygen species-induced molecular damage and its application in pathology. Pathol. Int. 1999, 49, 91–102. [Google Scholar] [CrossRef]
- Devadasan, S.; Sarkar, R.; Barman, K.D.; Kaushik, S. Role of serum melatonin and oxidative stress in childhood atopic dermatitis: A prospective study. Indian Dermatol. Online J. 2020, 11, 925. [Google Scholar]
- Bhaskaran, N.; Shukla, S.; Kanwal, R.; Srivastava, J.K.; Gupta, S. Induction of heme oxygenase-1 by chamomile protects murine macrophages against oxidative stress. Life Sci. 2012, 90, 1027–1033. [Google Scholar] [CrossRef]
- Li, W.; Kong, A.N. Molecular mechanisms of Nrf2-mediated antioxidant response. In Molecular Carcinogenesis; University of Texas MD Anderson Cancer Center: Houston, TX, USA, 2009; Volume 48, pp. 91–104. [Google Scholar]
- Wu, W.; Peng, G.; Yang, F.; Zhang, Y.; Mu, Z.; Han, X. Sulforaphane has a therapeutic effect in an atopic dermatitis murine model and activates the Nrf2/HO-1 axis. Mol. Med. Rep. 2019, 20, 1761–1771. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Qi, C.; Guo, H.; Jiao, X.; Yan, J.; Wang, Y.; Li, Q.; Zhao, M.; Guo, X. Ursolic acid ameliorates DNCB-induced atopic dermatitis-like symptoms in mice by regulating TLR4/NF-κB and Nrf2/HO-1 signaling pathways. Int. Immunopharmacol. 2023, 118, 110079. [Google Scholar] [CrossRef]
- Shim, K.-S.; Park, M.; Yang, W.-K.; Lee, H.; Kim, S.-H.; Choo, B.-K.; Chae, S.; Kim, H.-K.; Kim, T.; Kim, K.-M. Veronica persica Ethanol Extract Ameliorates Dinitrochlorobenzene-Induced Atopic Dermatitis-like Skin Inflammation in Mice, Likely by Inducing Nrf2/HO-1 Signaling. Antioxidants 2023, 12, 1267. [Google Scholar] [CrossRef]
- Furfaro, A.; Traverso, N.; Domenicotti, C.; Piras, S.; Moretta, L.; Marinari, U.; Pronzato, M.; Nitti, M. The Nrf2/HO-1 axis in cancer cell growth and chemoresistance. Oxidative Med. Cell. Longev. 2016, 2016, 1958174. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, Q.; Li, L.; Lin, L.; Yu, J.; Zhu, J.; Zhang, H.; Xia, G.; Zang, H. Antioxidant and enzyme-inhibitory activity of extracts from Erigeron annuus flower. Ind. Crops Prod. 2020, 148, 112283. [Google Scholar] [CrossRef]
- Jeong, M.; Ju, Y.; Kwon, H.; Kim, Y.; Hyun, K.-Y.; Choi, G.-E. Protocatechuic Acid and Syringin from Saussurea neoserrata Nakai Attenuate Prostaglandin Production in Human Keratinocytes Exposed to Airborne Particulate Matter. Curr. Issues Mol. Biol. 2023, 45, 5950–5966. [Google Scholar] [CrossRef] [PubMed]
- OECD. Guidelines for the Testing of Chemicals, Test № 407, Repeated Dose 28-Day Oral Toxicity Study in Rodents; OECD: Paris, France, 2008. [Google Scholar]
- Kim, Y.-H.; Kim, T.H.; Kang, M.S.; Ahn, J.-O.; Choi, J.H.; Chung, J.-Y. Comparison of the presentation of atopic dermatitis induced by trinitrochlorobenzene and house dust mite in NC/Nga mice. J. Vet. Sci. 2020, 21, e59. [Google Scholar] [CrossRef]
- Li, X.; Luo, Y.; Wang, L.; Li, Y.; Shi, Y.; Cui, Y.; Xue, M. Acute and subacute toxicity of ethanol extracts from Salvia przewalskii Maxim in rodents. J. Ethnopharmacol. 2010, 131, 110–115. [Google Scholar] [CrossRef]
- Bondet, V.; Brand-Williams, W.; Berset, C. Kinetics and mechanisms of antioxidant activity using the DPPH. free radical method. LWT-Food Sci. Technol. 1997, 30, 609–615. [Google Scholar] [CrossRef]
- Silverberg, N.B.; Silverberg, J.I. Inside out or outside in: Does atopic dermatitis disrupt barrier function or does disruption of barrier function trigger atopic dermatitis. Cutis 2015, 96, 359–361. [Google Scholar]
- Chen, Z.; Wang, C.; Yu, N.; Si, L.; Zhu, L.; Zeng, A.; Liu, Z.; Wang, X. INF2 regulates oxidative stress-induced apoptosis in epidermal HaCaT cells by modulating the HIF1 signaling pathway. Biomed. Pharmacother. 2019, 111, 151–161. [Google Scholar] [CrossRef]
- Bao, D.; Wang, J.; Pang, X.; Liu, H. Protective effect of quercetin against oxidative stress-induced cytotoxicity in rat pheochromocytoma (PC-12) cells. Molecules 2017, 22, 1122. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.N.; Kim, H.E.; Lee, S.G. Caffeoylserotonin protects human keratinocyte HaCaT cells against H2O2-induced oxidative stress and apoptosis through upregulation of HO-1 expression via activation of the PI3K/Akt/Nrf2 pathway. Phytother. Res. 2013, 27, 1810–1818. [Google Scholar] [CrossRef] [PubMed]
- Jayawardena, T.U.; Wang, L.; Sanjeewa, K.A.; Kang, S.I.; Lee, J.-S.; Jeon, Y.-J. Antioxidant potential of sulfated polysaccharides from Padina boryana; Protective effect against oxidative stress in in vitro and in vivo zebrafish model. Mar. Drugs 2020, 18, 212. [Google Scholar] [CrossRef] [PubMed]
- WHO. Research Guidelines for Evaluating the Safety and Efficacy of Herbal Medicines; WHO: Geneva, Switzerland, 1993; Volume 94. [Google Scholar]
- Galiniak, S.; Mołoń, M.; Biesiadecki, M.; Bożek, A.; Rachel, M. The role of oxidative stress in atopic dermatitis and chronic urticaria. Antioxidants 2022, 11, 1590. [Google Scholar] [CrossRef] [PubMed]
- Yan, C.; Ying, J.; Changzhi, Y.; Qihong, Q.; Jingzhu, M.; Dongjie, S.; Tingting, Z. MiR-1294 suppresses ROS-dependent inflammatory response in atopic dermatitis via restraining STAT3/NF-κB pathway. Cell. Immunol. 2022, 371, 104452. [Google Scholar] [CrossRef] [PubMed]
- Lee, S. Immunomodulatory Effects of Eurya emarginata on NC/Nga Mice as Models for Atopic Dermatitis. J. Life Sci. 2014, 24, 447–453. [Google Scholar] [CrossRef]
- Nomura, T.; Kabashima, K. Advances in atopic dermatitis in 2015. J. Allergy Clin. Immunol. 2016, 138, 1548–1555. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xian, Y.-F.; Hu, Z.; Loo, S.K.F.; Ip, S.P.; Chan, W.Y.; Lin, Z.-X.; Wu, J.C.Y. Efficacy and action mechanisms of a Chinese herbal formula on experimental models of atopic dermatitis. J. Ethnopharmacol. 2021, 274, 114021. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Li, S.; Liu, J.; Kong, D.; Han, X.; Lei, P.; Xu, M.; Guan, H.; Hou, D. Ameliorative effects of sea buckthorn oil on DNCB induced atopic dermatitis model mice via regulation the balance of Th1/Th2. BMC Complement. Med. Ther. 2020, 20, 263. [Google Scholar] [CrossRef] [PubMed]
- Ku, J.M.; Hong, S.H.; Kim, S.R.; Choi, H.-S.; Kim, H.I.; Kim, D.U.; Oh, S.M.; Seo, H.S.; Kim, T.Y.; Shin, Y.C. The prevention of 2, 4-dinitrochlorobenzene-induced inflammation in atopic dermatitis-like skin lesions in BALB/c mice by Jawoongo. BMC Complement. Altern. Med. 2018, 18, 215. [Google Scholar] [CrossRef]
- Jin, H.; He, R.; Oyoshi, M.; Geha, R.S. Animal models of atopic dermatitis. J. Investig. Dermatol. 2009, 129, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, S.M.U.; Luo, L.; Namani, A.; Wang, X.J.; Tang, X. Nrf2 signaling pathway: Pivotal roles in inflammation. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Bryan, H.K.; Olayanju, A.; Goldring, C.E.; Park, B.K. The Nrf2 cell defence pathway: Keap1-dependent and-independent mechanisms of regulation. Biochem. Pharmacol. 2013, 85, 705–717. [Google Scholar] [CrossRef]
- Helou, D.G.; Martin, S.F.; Pallardy, M.; Chollet-Martin, S.; Kerdine-Römer, S. Nrf2 involvement in chemical-induced skin innate immunity. Front. Immunol. 2019, 10, 1004. [Google Scholar] [CrossRef]
- Jin, W.; Botchway, B.O.; Liu, X. Curcumin can activate the Nrf2/HO-1 signaling pathway and scavenge free radicals in spinal cord injury treatment. Neurorehabilit. Neural Repair 2021, 35, 576–584. [Google Scholar] [CrossRef]
- Hennig, P.; Fenini, G.; Di Filippo, M.; Beer, H.-D. Electrophiles against (skin) diseases: More than Nrf2. Biomolecules 2020, 10, 271. [Google Scholar] [CrossRef]
- Beyer, T.; Auf dem Keller, U.; Braun, S.; Schäfer, M.; Werner, S. Roles and mechanisms of action of the Nrf2 transcription factor in skin morphogenesis, wound repair and skin cancer. Cell Death Differ. 2007, 14, 1250. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Ishitsuka, Y. The role of KEAP1-NRF2 system in atopic dermatitis and psoriasis. Antioxidants 2022, 11, 1397. [Google Scholar] [CrossRef]
- Kim, T.-Y.; Park, N.-J.; Jo, B.-G.; Paik, J.-H.; Choi, S.; Kim, S.-N.; Yang, M.H. 7-O-Methylluteolin Suppresses the 2, 4-Dinitrochlorobenzene-Induced Nrf2/HO-1 Pathway and Atopic Dermatitis-like Lesions. Antioxidants 2022, 11, 1344. [Google Scholar] [CrossRef] [PubMed]
- Han, E.-J.; Fernando, I.P.S.; Kim, H.-S.; Lee, D.-S.; Kim, A.; Je, J.-G.; Seo, M.-J.; Jee, Y.-H.; Jeon, Y.-J.; Kim, S.-Y. (–)-Loliolide Isolated from Sargassum horneri Suppressed Oxidative Stress and Inflammation by Activating Nrf2/HO-1 Signaling in IFN-γ/TNF-α-Stimulated HaCaT Keratinocytes. Antioxidants 2021, 10, 856. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Normal | EAE (mg/kg) | |
|---|---|---|---|
| 500 | 1000 | ||
| Males | |||
| Heart | 0.115 ± 0.021 | 0.113 ± 0.015 | 0.120 ± 0.012 |
| Liver | 0.978 ± 0.035 | 1.015 ± 0.061 | 1.033 ± 0.058 |
| Spleen | 0.093 ± 0.008 | 0.095 ± 0.015 | 0.090 ± 0.007 |
| Kidney | 0.333 ± 0.011 | 0.335 ± 0.038 | 0.350 ± 0.017 |
| Lung | 0.170 ± 0.036 | 0.175 ± 0.025 | 0.170 ± 0.025 |
| Females | |||
| Heart | 0.108 ± 0.020 | 0.100 ± 0.012 | 0.105 ± 0.009 |
| Liver | 0.793 ± 0.029 | 0.800 ± 0.047 | 0.803 ± 0.036 |
| Spleen | 0.090 ± 0.010 | 0.088 ± 0.013 | 0.078 ± 0.004 |
| Kidney | 0.245 ± 0.021 | 0.250 ± 0.025 | 0.238 ± 0.022 |
| Lung | 0.145 ± 0.005 | 0.140 ± 0.012 | 0.143 ± 0.015 |
| Parameters | Normal | EAE (mg/kg) | |
|---|---|---|---|
| 500 | 1000 | ||
| Males | |||
| WBC (103/µL) | 5.113 ± 0.342 | 5.003 ± 0.328 | 4.947 ± 0.483 |
| LY (%) | 96.343 ± 0.313 | 96.693 ± 0.054 | 96.397 ± 1.666 |
| MO (%) | 0.833 ± 0.090 | 0.870 ± 0.179 | 0.840 ± 0.067 |
| NE (%) | 2.167 ± 0.107 | 2.027 ± 0.131 | 2.227 ± 0.727 |
| EO (%) | 0.167 ± 0.029 | 0.143 ± 0.021 | 0.153 ± 0.034 |
| RBC (106/µL) | 6.907 ± 0.612 | 6.873 ± 0.596 | 7.055 ± 0.154 |
| PLT (103/µL) | 836.433 ± 86.753 | 813.800 ± 80.851 | 795.675 ± 45.653 |
| HGB (g/dL) | 14.777 ± 0.614 | 14.968 ± 0.956 | 15.155 ± 0.831 |
| HCT (%) | 43.833 ± 2.695 | 43.600 ± 3.439 | 45.200 ± 2.239 |
| MCV (fL) | 63.600 ± 2.707 | 63.475 ± 0.750 | 62.325 ± 2.552 |
| MCH (pg) | 21.500 ± 1.042 | 21.825 ± 0.580 | 20.900 ± 1.022 |
| MCHC (g/dL) | 33.767 ± 0.953 | 34.350 ± 0.541 | 33.525 ± 0.432 |
| Females | |||
| WBC (103/µL) | 3.650 ± 0.282 | 3.867 ± 0.212 | 3.817 ± 0.095 |
| LY (%) | 95.850 ± 0.947 | 96.587 ± 0.764 | 96.133 ± 0.910 |
| MO (%) | 1.300 ± 0.211 | 1.377 ± 0.437 | 1.273 ± 0.127 |
| NE (%) | 1.697 ± 0.418 | 1.607 ± 0.450 | 1.757 ± 0.452 |
| EO (%) | 0.227 ± 0.025 | 0.210 ± 0.051 | 0.213 ± 0.040 |
| RBC (106/µL) | 6.573 ± 0.510 | 7.080 ± 0.262 | 6.987 ± 0.262 |
| PLT (103/µL) | 857.133 ± 35.171 | 823.633 ± 65.037 | 818.100 ± 34.851 |
| HGB (g/dL) | 14.120 ± 0.115 | 15.257 ± 0.794 | 14.930 ± 0.692 |
| HCT (%) | 39.900 ± 0.735 | 42.767 ± 1.605 | 42.667 ± 0.785 |
| MCV (fL) | 60.500 ± 4.032 | 61.833 ± 2.517 | 61.067 ± 1.482 |
| MCH (pg) | 21.600 ± 1.715 | 21.600 ± 1.687 | 21.433 ± 1.470 |
| MCHC (g/dL) | 35.700 ± 0.510 | 34.833 ± 1.352 | 35.000 ± 1.558 |
| Parameters | Normal | EAE (mg/kg) | |
|---|---|---|---|
| 500 | 1000 | ||
| Males | |||
| ALP (U/L) | 43.967 ± 14.281 | 39.467 ± 3.262 | 43.767 ± 7.144 |
| ALT (U/L) | 42.667 ± 2.357 | 44.333 ± 2.494 | 49.000 ± 3.742 |
| AST (U/L) | 183.000 ± 3.559 | 186.333 ± 5.558 | 191.667 ± 6.182 |
| TB (mg/dL) | 1.160 ± 0.008 | 1.177 ± 0.009 | 1.177 ± 0.009 |
| TP (g/dL) | 5.457 ± 0.054 | 5.737 ± 0.153 | 5.717 ± 0.270 |
| ALB (g/dL) | 3.060 ± 0.037 | 3.253 ± 0.116 | 3.247 ± 0.049 |
| TG (mg/dL) | 38.533 ± 0.499 | 37.167 ± 2.193 | 40.567 ± 6.975 |
| TC (mg/dL) | 112.000 ± 3.742 | 129.333 ± 10.209 | 126.267 ± 19.072 |
| LDH (U/L) | 416.000 ± 4.546 | 433.333 ± 23.977 | 447.333 ± 25.773 |
| Females | |||
| ALP (U/L) | 11.133 ± 2.106 | 11.117 ± 1.899 | 12.133 ± 1.791 |
| ALT (U/L) | 33.000 ± 0.816 | 31.000 ± 1.633 | 30.333 ± 0.471 |
| AST (U/L) | 118.333 ± 4.497 | 112.333 ± 10.209 | 110.867 ± 9.015 |
| TB (mg/dL) | 1.133 ± 0.005 | 1.137 ± 0.005 | 1.123 ± 0.005 |
| TP (g/dL) | 4.997 ± 0.302 | 4.633 ± 0.139 | 4.533 ± 0.097 |
| ALB (g/dL) | 2.830 ± 0.134 | 2.787 ± 0.100 | 2.660 ± 0.090 |
| TG (mg/dL) | 54.867 ± 6.715 | 57.567 ± 8.892 | 58.033 ± 4.151 |
| TC (mg/dL) | 67.467 ± 3.332 | 62.333 ± 5.526 | 60.833 ± 2.941 |
| LDH (U/L) | 380.667 ± 20.677 | 372.333 ± 10.873 | 372.000 ± 6.377 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, M.; Kwon, H.; Kim, Y.; Jin, H.; Choi, G.-E.; Hyun, K.-Y. Erigeron annuus Extract Improves DNCB-Induced Atopic Dermatitis in a Mouse Model via the Nrf2/HO-1 Pathway. Nutrients 2024, 16, 451. https://doi.org/10.3390/nu16030451
Jeong M, Kwon H, Kim Y, Jin H, Choi G-E, Hyun K-Y. Erigeron annuus Extract Improves DNCB-Induced Atopic Dermatitis in a Mouse Model via the Nrf2/HO-1 Pathway. Nutrients. 2024; 16(3):451. https://doi.org/10.3390/nu16030451
Chicago/Turabian StyleJeong, Myeongguk, Hyeokjin Kwon, Yeeun Kim, Hyunwoo Jin, Go-Eun Choi, and Kyung-Yae Hyun. 2024. "Erigeron annuus Extract Improves DNCB-Induced Atopic Dermatitis in a Mouse Model via the Nrf2/HO-1 Pathway" Nutrients 16, no. 3: 451. https://doi.org/10.3390/nu16030451
APA StyleJeong, M., Kwon, H., Kim, Y., Jin, H., Choi, G.-E., & Hyun, K.-Y. (2024). Erigeron annuus Extract Improves DNCB-Induced Atopic Dermatitis in a Mouse Model via the Nrf2/HO-1 Pathway. Nutrients, 16(3), 451. https://doi.org/10.3390/nu16030451