1. Introduction
Bone has important physiological functions such as protecting the vital organs of the body, storing minerals, and providing a hematopoietic environment [
1,
2]. Any pathology of bone may damage one or more bodily functions [
3]. The bone formation and absorption cycle maintains bone health during bone development and throughout the life of the animal [
4,
5]. The balance is regulated by hormones, vitamins, growth factors, and cytokines [
6], especially the trace element metabolism of the body [
7]. In children and adolescents, for the accumulation of bone mass, bone formation is greater than bone resorption, but the opposite is true in adults (especially in old age), when bone loss is faster. Therefore, the amount of bone mass accumulated in the early stage of life is a determining factor of the level of bone mass in the later years and the occurrence of fractures due to bone fragility.
Optimizing calcium and protein intake during the growth process increases bone formation in the early stage of bone development so that the body can obtain the best peak bone mass and strength. As the main inorganic component of bone, calcium plays an irreplaceable role in the human body [
8,
9]; 99% of the body’s calcium is stored in the bones in the form of hydroxyapatite, which contributes to their strength. The body’s serum calcium concentration is usually stable, and fluctuation triggers the regulation of calcium metabolism with the deposition or release of bone calcium and the absorption of calcium [
10,
11]. In addition, in the process of maintaining the stability of serum calcium, parathyroid hormone (PTH), 1,25(OH)2D3 (the active form of vitamin D3), and calcitonin (CT) maintain calcium homeostasis by acting on the bone, kidney, and intestine [
12,
13]. PTH mobilizes bone calcium into the blood when serum calcium is decreased and activates 1,25(OH)2D3 to promote intestinal calcium absorption. When serum calcium is increased, CT is released to reduce serum calcium by increasing the excretion of ionic calcium by the kidney, which promotes bone calcium deposition and inhibits its absorption in the intestine.
Bovine colostrum basic protein (CBP) is a milky white powder containing a large amount of protein that is obtained from bovine colostrum by sterilization, degreasing, centrifugation, and removal of casein, α-lactoprotein, and β-lactoglobulin. The composition of CBP is shown in
Table 1. It is believed that proteins with molecular weights of 1–30 kDa are the key components of CBP in promoting bone activity [
14]. However, due to the complex and diverse protein components of CBP, the biological activity of specific protein types has not been analyzed. Interest in the use of CBP to improve bone health is growing because CBP appears to be a potent bone-stimulating factor. CBP potently promoted the bone mineral density (BMD) of rats. In vivo experiments showed that it increased the content of serum osteoblastic markers, indicating that CBP can regulate bone metabolism and promote bone growth [
14]. However, the maintenance of bone growth and health is also affected by the body’s mineral metabolism [
15]. Therefore, this study seeks to explore whether dietary CBP can affect bone development by regulating calcium metabolism.
2. Materials and Methods
2.1. Animals and Experimental Design
In a controlled environment (12-h light/dark cycle; temperature: 22 ± 1 °C), 100 male C57BL/6JN mice (Beijing Vital River Laboratory Animal Technology) aged 4 week were randomly assigned (
n = 20 mice per group) to 5 groups: normal control (NC), 0.015% CBP, 0.15% CBP, 1.5% CBP, and 5% CBP. The composition of both diets is shown in
Table 2. Apart from protein composition, all the other constituents were identical between these 5 diets. All animals were obtained from Weitong Lihua Laboratory Animal Technology Co., Ltd. (Beijing, China).
After 4 week, mice (n = 3) were placed in a special mouse metabolic cage that could separate feces and urine through a funnel at the bottom; the feed intake, urine volume, and fecal volume of mice were recorded within 24-h. A single mouse’s feces should be dried in a drying oven to a constant weight before being weighed and recorded. Collected urine and dried feces were stored at −20 °C for the detection of calcium content. Mice (n = 12) were necropsied, and femur and partial small intestinal segments were excised. The left femur (n = 6) was fixed with 4% paraformaldehyde for microscopic CT analysis. The right part of the femur from the same mouse (n = 6) was wrapped with gauze impregnated with normal saline and stored at −20 °C for bone biomechanical examination. The left tibia (n = 6) from the same mouse was prepared for bone mineral detection. The remaining left femur (n = 6) was for tissue staining, and the remaining right femur (n = 6) was dried in the oven to estimate the weight, length, and diameter of the femur. After centrifuging at 3000× g for 15 min at 4 °C, serum (n = 6) was obtained and stored at −80 °C for calcium (105-000453-00, Mindray, Shenzhen, China), phosphorus (105-015568-00, Mindray, Shenzhen, China), parathyroid hormone (JN19040, Jining Shiye, Shanghai, China), and calcitonin concentration (JN19883, Jining Shiye, Shanghai, China). A portion of the jejunum and ileum were flash-frozen in liquid nitrogen for later protein extraction.
2.2. Body Composition Analysis
Each mouse was weighed and then detected with a sober animal body composition analyzer (QMR, Niumag Corporation, Suzhou, China) for the content and proportion of fat and muscle in the body.
2.3. Motor Ability Test
A motor ability test was performed by the motorized treadmill (ZS-PT-III, Zhongshi Technology, Beijng, China). After 3 days of acclimation, the mice were placed on the treadmill at the speed of 3 m/min. The running speed increased every minute by 1.8 m/min, and the duration of motion was recorded until the mice showed fatigue, defined by an inability to return to the treadmill or staying on the electrical shock grids for 10 s.
2.4. Bone Biomechanical Testing
The right femur samples stored at −20 °C were defrosted at room temperature and then tested by a Univert biomechanical test analyzer (UV-200-01, Cellscale Biomaterials Testing, Waterloo, ON, Canada). The individual femur was placed horizontally, with the broad side of the femur facing upward on two support points with a span of 10 mm. The workstation was operated to make the probe of the tester slowly drop; the loading speed of the probe was 2 mm/min and continued to run 2 mm after the specimen broke. The original data and compression curve were obtained through calculation. The bone mechanical characteristic parameters were analyzed, including the maximum load, stiffness, energy to ultimate load, stress–strain, and breaking energy.
2.5. Tissue Staining
Mouse left femurs were fixed with 4% formaldehyde for 24 h, then embedded in paraffin and cut into 4 μm thick slices. Bone sections were stained by the HE kit (Servicebio, Wuhan, China) and the Masson kit (Servicebio, China). The area of the blue region in the cavity was analyzed using ImageJ v1.53.
2.6. Micro-CT Analysis
The left femurs were soaked in 4% paraformaldehyde over 24 h and then analyzed immediately by a micro-CT system (SkyScan1276, Bruker microCT company, Kontich, Belgium) using 60 kV voltage, 140 μA current, and 9 μm resolution ratio in the distal growth plate of femurs. After the scan was completed, the 3D images were reconstructed. Dataviewer software 1.5.6.2 was used to adjust the direction and other parameters of the scanned sample images to ensure all samples were processed under the same conditions to generate VOI images. Meanwhile, the bone density formula was constructed in CTAn software 1.17.7.2 using standard product parameters. Then, the single VOI image of each sample was imported to select 100–200 layers below the femoral growth plate to obtain bone morphometric parameters, including bone mineral density (BMD), bone volume fraction (BV/TV), trabecular thickness (Tb.Th), trabecular space (Tb.Sp) and trabecular number (Tb.N). BV/TV is the ratio of the total volume of voxels representing bone structures in the ROI to the total volume of all voxels in the region. Tb.Th is the average thickness of the trabecular bone. Tb.N is the number of intersections between bone tissue and non-bone tissue in a given length of bone. Tb.Sp is the average width of the pulp cavity between the trabeculae, indicating the porosity of the trabecular bone.
2.7. Determination of Calcium Content in Bone, Urine, and Stool
The left tibia of the dried mouse was placed in a container. Then, about 5 mL nitric acid and 1 mL hydrogen peroxide were added and the container was heated at 180 degrees for digestion. After the acid was volatilized to the whole volume of 1–2 mL, the volume was fixed to 50 mL with 1% dilute nitric acid. IPC-OES (Agilent company, Santa Clara, CA, USA) was used to detect the content of calcium and phosphorus ions in samples. The calcium content of each mouse’s feed, preserved urine, and feces were digested by the same machine. The IPC machine was calibrated with 50 mL, 100 μg/mL calcium standard GSB 04-2824-2011.
2.8. Western Blot
Jejunum and ileum tissues were lysed using RIPA buffer (Beyotime, Shanghai, China) for 30 min in ice and then centrifuged at 12,000× g for 15 min at 4 °C to obtain the total protein. Protein concentrations were determined by a BCA kit (Beyotime, China). Antibodies for TRPV6 (DF12784, 1:1000), S100G (DF9785, 1:1000), and Claudin-2 (AF0128, 1:1000) were purchased from Affinity (Changzhou, China). Recombinant anti-PMCA1 antibody (ab190355, 1:1000) and β-actin (ab8226, 1:5000) were obtained from Abcam (Shanghai, China), and Claudin-12 antibody (NBP1-87450,1:1000) was from Novus (Shanghai, China). Horseradish peroxidase-labeled goat anti-rabbit antibody (Beyotime, China) was used as a secondary antibody. The expression levels of TRPV6, CaBP-9k, PMCA1, CLDN2, and CLDN12 proteins were detected in the jejunum. The expression levels of CLDN2 and CLDN12 protein were detected in the ileum.
2.9. Statistical Analysis
All statistical analyses were performed using SPSS 26.0., and the results were shown as mean ± SEM. One-factor ANOVA followed by Duncan’s post hoc test was used to evaluate differences between groups, with different letters representing statistical significance (p < 0.05).
4. Discussion
Ninety percent of the bone mass of the human body is accumulated before the age of 20, and bone grows rapidly in children and adolescents. In the present study, the initially 4-week-old mice were fed for 30 days so that the feeding time essentially covered the equivalent of human childhood and adolescence. Studies have shown that 5% CBP given as a dietary supplement can significantly increase bone mineral density and serum levels of osteocalcin, growth hormone, and insulin-like growth factor-1 in young rats [
14], and in 2021, our research group showed that dietary 0.015% CBP combined with milk, calcium, and vitamin D improved bone indexes in low-calcium rats. Therefore, here, four experimental feeding doses (0.015%, 0.15%, 1.5%, and 5% CBP) were applied.
In this study, the proportion of CBP was changed while the amount of total protein in the diet of the mice was kept the same. With increasing CBP dose, the weight growth rate of mice increased significantly. There was no significant difference in organ index and body composition of mice between the NC group and the CBP group, indicating that the basic physiological condition of the mice was not greatly affected by the CBP dose. We observed no significant difference in fat and lean meat mass between groups of mice. Movement of the body is coordinated by bones, muscles, and joints, and the bones, to which muscles and tendons are attached, provide support for the movement. Healthy bones are, thus, the basis of normal movement of the body, and the movement ability of the body can reflect the strength of the bones to a certain extent. Therefore, the increase in treadmill exercise ability of mice fed CBP in this study reflects the health of their bones.
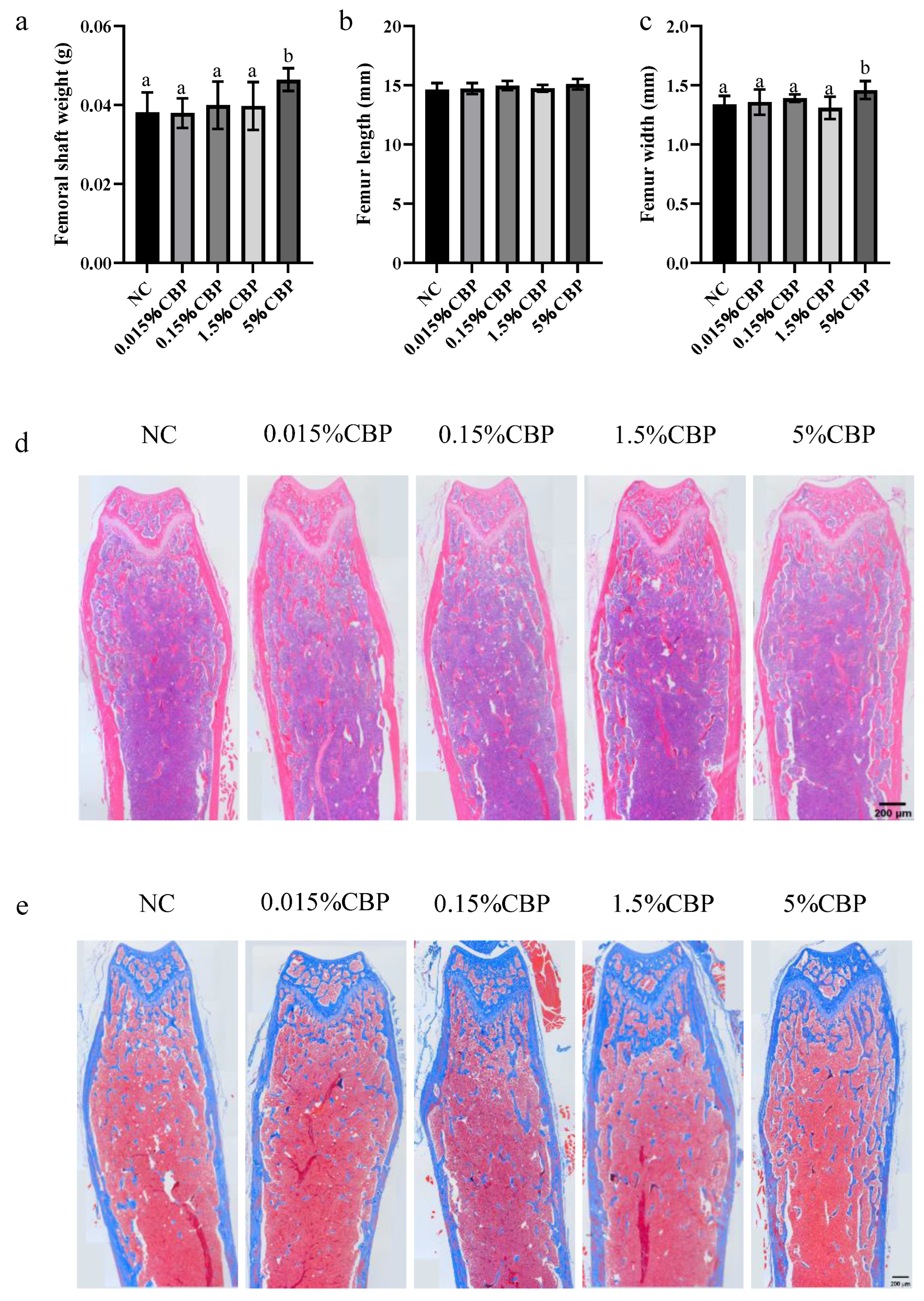
The results show that a certain amount of CBP increased the weight and width of the femur in mice. The index of bone mechanical properties shows the resistance of bone to stress and tension, while the BMD directly represents the bone mass. The current study showed that a high (5%) CBP diet markedly contributes to the compression and bending resistance, rather than the elasticity, of bone. Meanwhile, 5% CBP intake significantly enhanced the BMD of the mice. Numerous previous studies have shown that the structure of bone trabeculae plays an important role in the mechanical properties of bone independently of bone density [
16]. Changes in the microstructure of trabeculae, such as the number, thickness, and cross structure, affect the strength and mechanical properties of bone. The distal femur is often used to detect bone health [
17]. Predictably, it was found here that the bone trabecular indexes above were significantly improved by 5% intake. Through the detection of quantitative parameters of bone health, this study comprehensively evaluated bone growth and showed that CBP is beneficial for bone growth.
The bone matrix contains 70% inorganic mineral salts, which contribute to the rigidity of bone. Calcium is the key element in bone mineral salts, and the balance of calcium metabolism in the body is of great significance for the maintenance of bone health [
9]. Stable serum calcium levels ensure normal physiological activities of the body. The body produces PTH to activate osteoclasts and release bone calcium to replenish serum calcium when needed; if the serum calcium level is consistently too low, long-term effects lead to a large loss of bone calcium and bone damage. When the serum calcium level is too high, CT is produced, and osteoclast activity is inhibited. Bone absorption is reduced, and bone formation continues, which promotes the conversion of serum calcium into stored calcium in bone. Therefore, the homeostasis of calcium metabolism is a critical factor affecting bone formation [
18]. Dietary calcium is the only source of calcium in the body; it is absorbed into the blood through the intestine. When the intake of calcium is the same but the absorption of intestinal calcium increases, the deposition of bone calcium will be triggered [
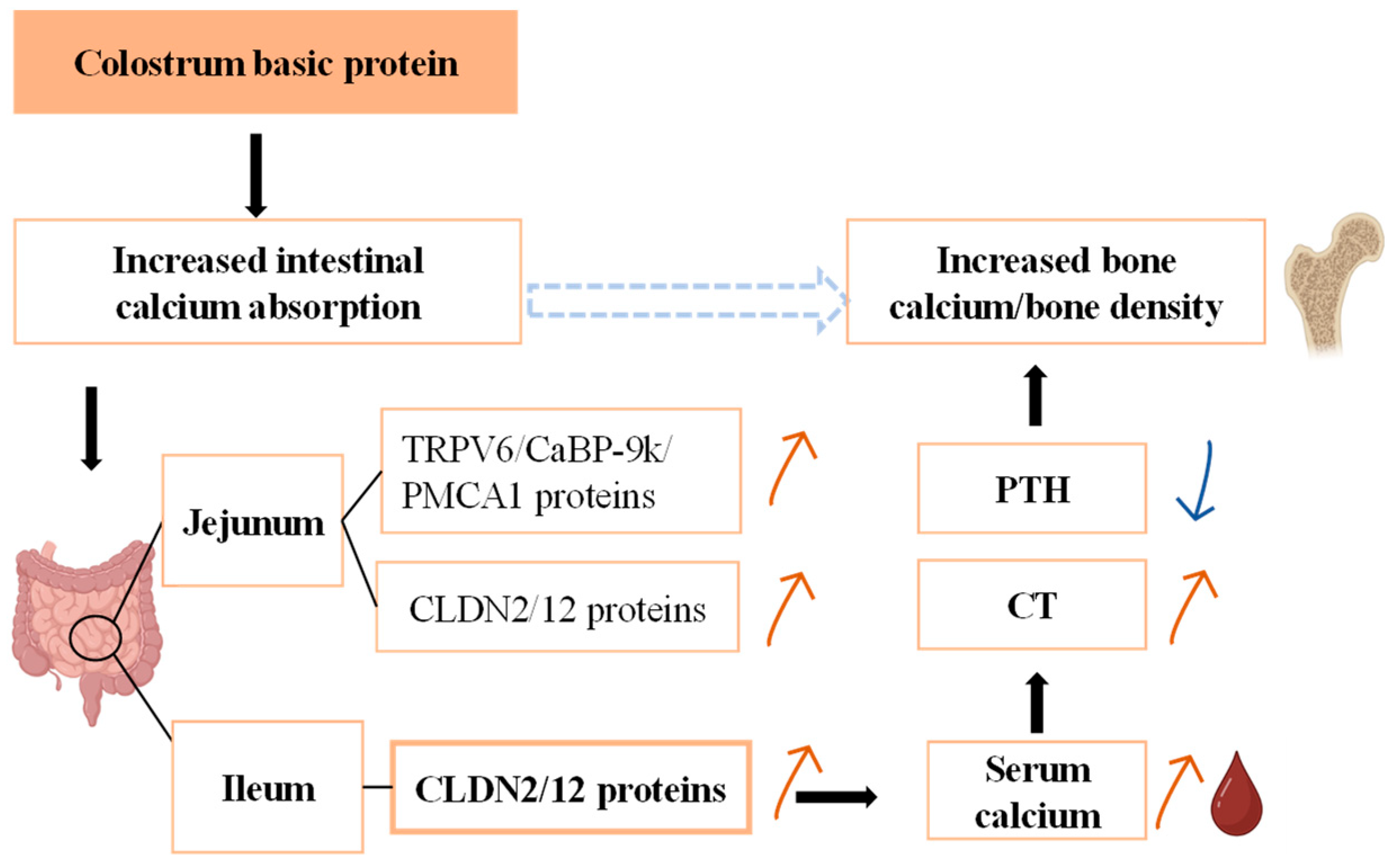
11]. In 2021, our research group showed that dietary 0.015% CBP, in combination with milk and calcium, decreased the excretion of fecal calcium in rats. It can be seen that CBP has the potential to promote bone growth, which may be through increased intestinal calcium absorption, thereby modulating the body’s calcium metabolism.
Here, we explored whether CBP promotes bone growth by affecting calcium metabolism. It was found that the bone strength and bone calcium and phosphorus contents of mice fed a CBP-containing diet increased significantly compared with the NC group. Phosphorus is also an important component of bone; 60% of phosphorus and calcium in the body are present in bone, in a fixed ratio [
18], and thus, the content of phosphorus in bone can also reflect the extent of bone calcium deposition. Meanwhile, the content of calcium and phosphorus in the blood must be maintained in proportion to maintain the balance of calcium and phosphorus metabolism [
19]. After calcium ingested by the body is absorbed into the blood circulation, most of the unused calcium will be excreted in stool, and a small fraction will be excreted in urine after renal reabsorption. The amount of calcium reabsorbed by the kidney is much lower than that absorbed by the intestine, and thus, the contribution of intestinal calcium absorption to serum calcium is higher than that of renal reabsorption [
20]. CBP may regulate calcium metabolism mainly by affecting intestinal calcium absorption. In this study, the decrease in calcium content in the feces of mice fed CBP was consistent with this hypothesis. The significant increase in serum calcium in the 5% CBP group may be due to the fact that CBP mainly affects the absorption of intestinal calcium, and the significant increase in serum CT observed in this study shows that the significant change in serum calcium affects the balance of calcium metabolism in the body.
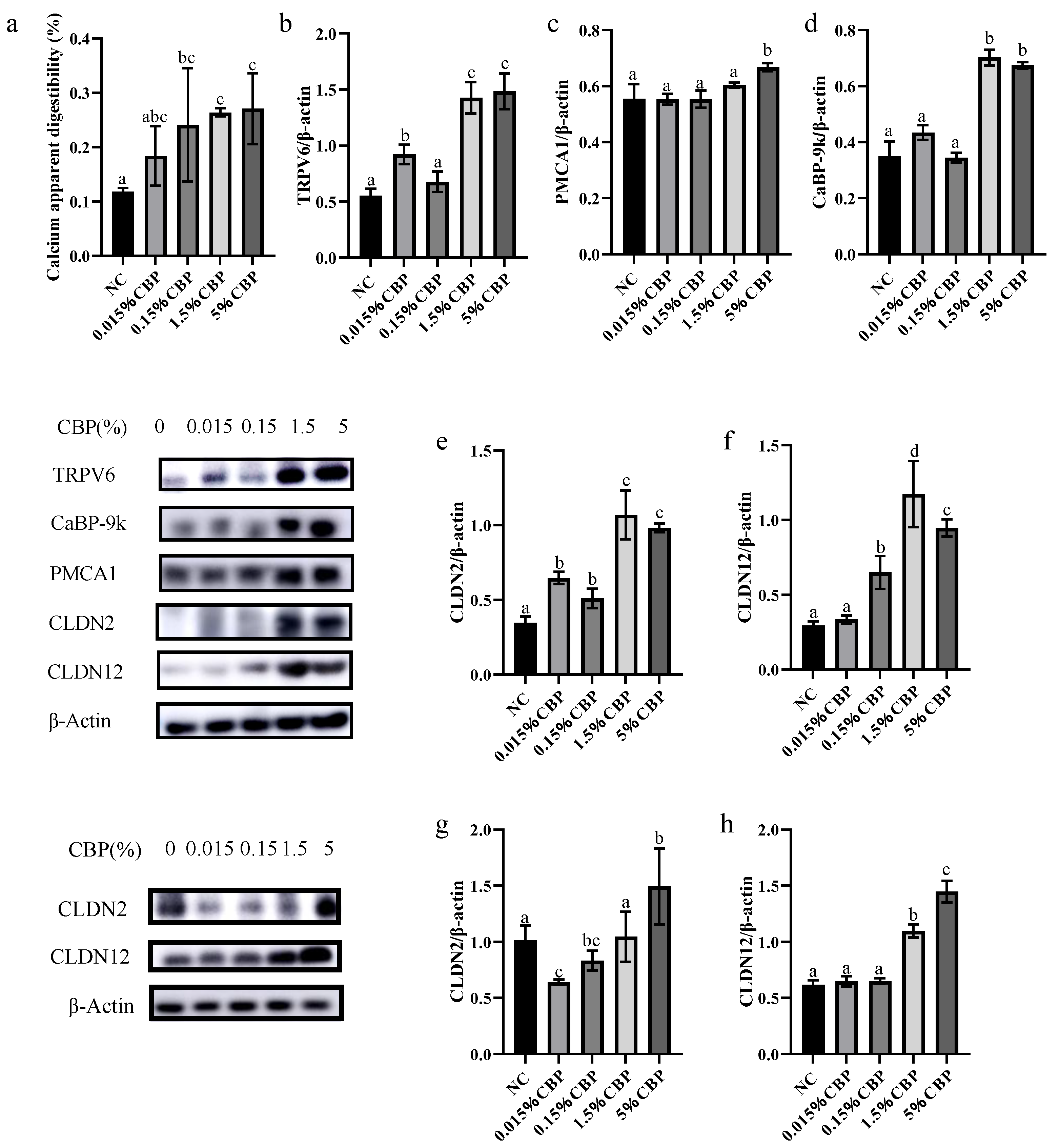
The ileum accounts for >40% of bodily calcium absorption, the largest contribution to the total, and the jejunum is ranked second, accounting for about 17%. Intestinal epithelial cells absorb calcium in two main ways: energy-dependent transcellular transport and concentration-dependent paracellular transport. Absorption by the former may be saturated, while the latter is not [
21]. If there is insufficient calcium intake into the body, calcium intake is dominated by transcellular transport in the duodenum. In the case of sufficient calcium intake, paracellular transport in the ileum dominates (while transcellular transport of calcium in the ileum is silent). Both modes of calcium transport occur in the jejunum. In this study, we found that the expression levels of TRPV6, CaBP-9k, PMCA1, CLDN2, and CLDN12 were all increased in the jejunum by 5% CBP. The expression levels of CLDN2 and CLDN12 in the ileum were also increased in the 5% CBP group. These findings were consistent with the characteristic calcium absorption modes in each intestinal segment, indicating that CBP had a promoting effect on intestinal epithelial calcium absorption. Because the calcium absorption rate of the ileum is significantly higher than that of the jejunum, and the jejunum contains more intestinal endocrine calcium, the calcium absorption of the ileum contributes more to overall serum calcium change. We thus speculate that CBP mainly mediates bone growth by promoting the expression of key proteins in the ileum.
However, we believe that it is useful to explore the harm of high doses of CBP to the human body, and whether it contains bound calcium itself is also worth studying. This study explored the function of CBP in promoting calcium absorption in vivo, suggesting that CBP may be used in combination with calcium as a supplement to achieve better bone-promoting effects in future product applications. In our experiments, CBP may be more appropriate to be used to increase peak bone mass in children and adolescents. Its role in older adults with advanced bone loss or osteoporosis is unclear.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}