
Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Culturing of Human Endocardial Endothelial Cells
2.2. Confocal Microscopy
2.3. Determination of the Cell Volume
2.4. Loading with the Calcium and ROS Fluorescent Probes
2.5. Statistical Analyses
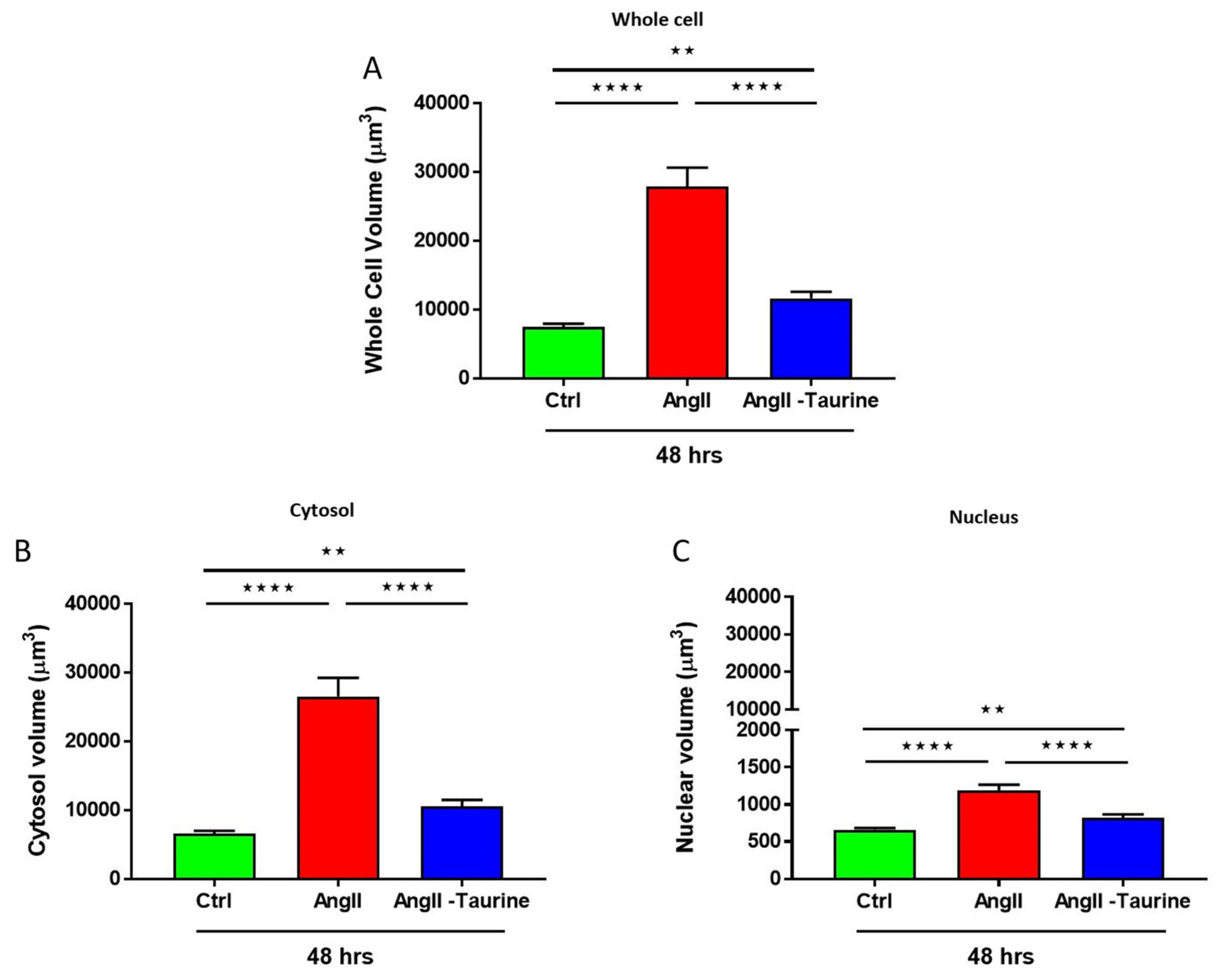
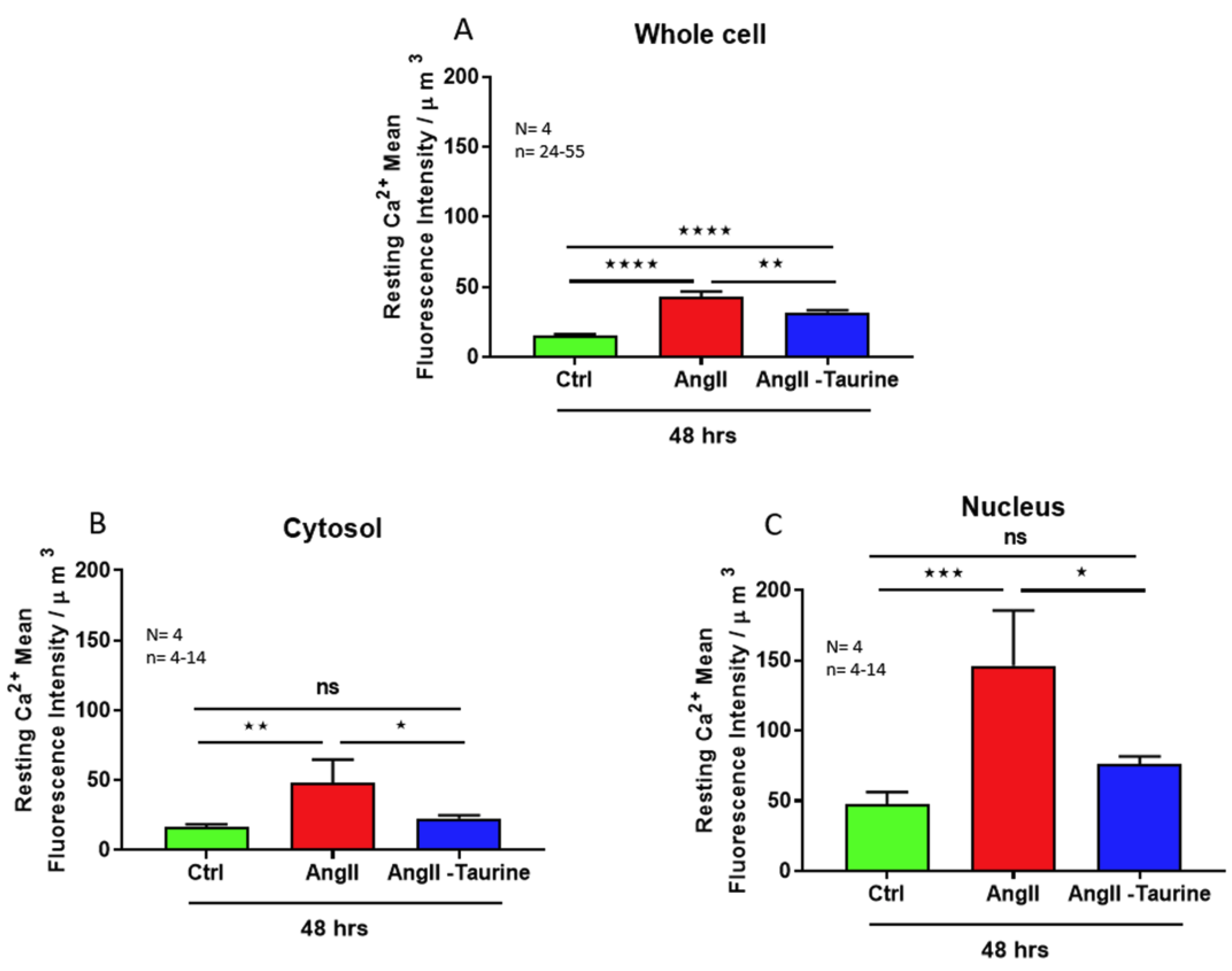
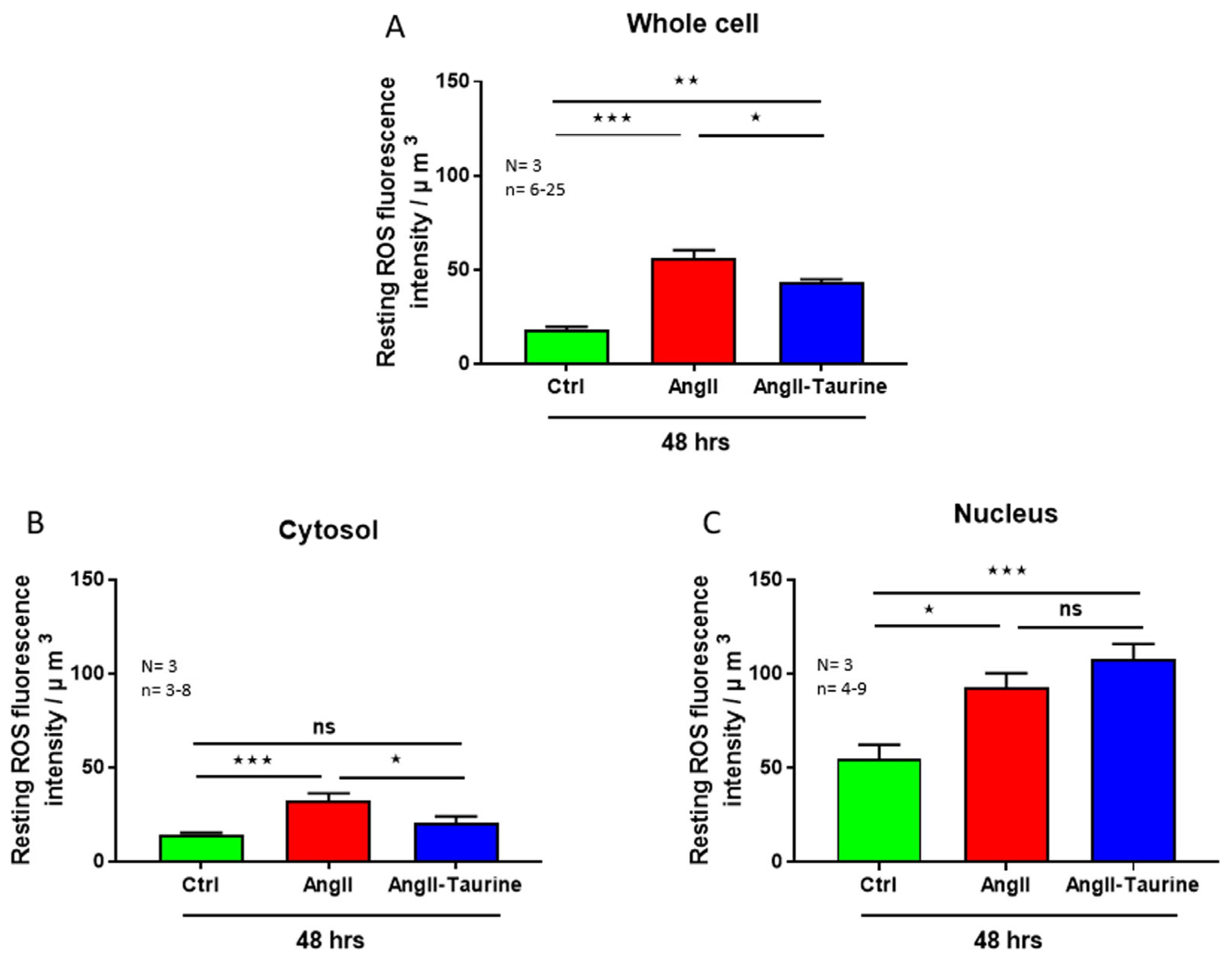
3. Results
Effect of Ang II on Human Ventricular Endocardial Endothelial Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kuruvilla, L.; Kartha, C.C. Molecular mechanisms in endothelial regulation of cardiac function. Mol. Cell. Biochem. 2003, 253, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Brutsaert, D.L.; De Keulenaer, G.W.; Fransen, P.; Mohan, P.; Kaluza, G.L.; Andries, L.J.; Rouleau, J.L.; Sys, S.U. The cardiac endothelium: Functional morphology, development, and physiology. Prog. Cardiovasc. Dis. 1996, 39, 239–262. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Jacques, D. Morphological and Functional Remodeling of Vascular Endothelium in Cardiovascular Diseases. Int. J. Mol. Sci. 2023, 24, 1998. [Google Scholar] [CrossRef] [PubMed]
- Lüscher, T.F.; Barton, M. Biology of the endothelium. Clin. Cardiol. 1997, 20 (Suppl. S2), II-3–II-10. [Google Scholar] [CrossRef]
- Mombouli, J.V.; Vanhoutte, P.M. Endothelial dysfunction: From physiology to therapy. J. Mol. Cell. Cardiol. 1999, 31, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Nilius, B.; Droogmans, G. Ion channels and their functional role in vascular endothelium. Physiol. Rev. 2001, 81, 1415–1459. [Google Scholar] [CrossRef]
- Shah, A.M.; Grocott-Mason, R.M.; Pepper, C.B.; Mebazaa, A.; Henderson, A.H.; Lewis, M.J.; Paulus, W.J. The cardiac endothelium: Cardioactive mediators. Prog. Cardiovasc. Dis. 1996, 39, 263–284. [Google Scholar] [CrossRef]
- Brutsaert, D.L. Cardiac endothelial-myocardial signaling: Its role in cardiac growth, contractile performance, and rhythmicity. Physiol. Rev. 2003, 83, 59–115. [Google Scholar] [CrossRef]
- Xiao, F.; Puddefoot, J.R.; Vinson, G.P. The expression of renin and the formation of angiotensin II in bovine aortic endothelial cells. J. Endocrinol. 2000, 164, 207–214. [Google Scholar] [CrossRef]
- Baudin, B.; Berard, M.; Carrier, J.L.; Legrand, Y.; Drouet, L. Vascular origin determines angiotensin I-converting enzyme expression in endothelial cells. Endothelium 1997, 5, 73–84. [Google Scholar] [CrossRef]
- Carley, W.W.; Tanoue, L.; Merker, M.; Gillis, C.N. Isolation of rabbit pulmonary microvascular endothelial cells and characterization of their angiotensin converting enzyme activity. Pulm. Pharmacol. 1990, 3, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Ryan, U.S.; Ryan, J.W.; Whitaker, C.; Chiu, A. Localization of angiotensin converting enzyme (kininase II). II. Immunocytochemistry and immunofluorescence. Tissue Cell 1976, 8, 125–145. [Google Scholar] [CrossRef]
- Yamazaki, T.; Yazaki, Y. Molecular basis of cardiac hypertrophy. Z. Kardiol. 2000, 89, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bhullar, S.K.; Dhalla, N.S. Adaptive and maladaptive roles of different angiotensin receptors in the development of cardiac hypertrophy and heart failure. Can. J. Physiol. Pharmacol. 2024, 102, 86–104. [Google Scholar] [CrossRef] [PubMed]
- Jacques, D.B.G. Cardiovascular physiopathology of angiotensin II and its plasma and nuclear envelop mem-branes’ receptors. In The Renin Angiotensin System in Cardiovascular Disease. Advances in Biochemistry in Health and Disease; Dhalla, N.S., Bhullar, S.K., Shah, A.K., Eds.; Springer: Cham, Switzerland, 2023; pp. 63–80. [Google Scholar]
- Vandemark, C.; Nguyen, J.; Zhao, Z.Q. Cardiovascular Protection with a Long-Acting GLP-1 Receptor Agonist Liraglutide: An Experimental Update. Molecules 2023, 28, 1369. [Google Scholar] [CrossRef]
- Hussain, M.; Ikram, W.; Ikram, U. Role of c-Src and reactive oxygen species in cardiovascular diseases. Mol. Genet. Genom. 2023, 298, 315–328. [Google Scholar] [CrossRef]
- Bkaily, G.; Simon, Y.; Normand, A.; Jazzar, A.; Najibeddine, H.; Khalil, A.; Jacques, D. Short-Communication: Short-Term Treatment with Taurine Prevents the Development of Cardiac Hypertrophy and Early Death in Hereditary Cardiomyopathy of the Hamster and Is Sex-Dependent. Nutrients 2022, 14, 3287. [Google Scholar] [CrossRef]
- Bkaily, G.; Jazzar, A.; Normand, A.; Simon, Y.; Al-Khoury, J.; Jacques, D. Taurine and cardiac disease: State of the art and perspectives. Can. J. Physiol. Pharmacol. 2020, 98, 67–73. [Google Scholar] [CrossRef]
- Ahmed, K.; Choi, H.N.; Park, J.S.; Kim, Y.G.; Bae, M.K.; Yim, J.E. Taurine supplementation alters gene expression profiles in white adipose tissue of obese C57BL/6J mice: Inflammation and lipid synthesis perspectives. Heliyon 2024, 10, e23288. [Google Scholar] [CrossRef]
- Padovan, J.C.; Dourado, T.M.H.; Pimenta, G.F.; Bruder-Nascimento, T.; Tirapelli, C.R. Reactive Oxygen Species Are Central Mediators of Vascular Dysfunction and Hypertension Induced by Ethanol Consumption. Antioxidants 2023, 12, 1813. [Google Scholar] [CrossRef]
- Braun, H.; Hauke, M.; Petermann, M.; Eckenstaler, R.; Ripperger, A.; Schwedhelm, E.; Ludwig-Kraus, B.; Bernhard Kraus, F.; Jalal Ahmed Shawon, M.; Dubourg, V.; et al. Deletion of vascular thromboxane A(2) receptors and its impact on angiotensin II-induced hypertension and atherosclerotic lesion formation in the aorta of Ldlr-deficient mice. Biochem. Pharmacol. 2024, 219, 115916. [Google Scholar] [CrossRef]
- Wang, J.; Li, D.; Zhang, Y.; Xing, D.; Lei, Z.; Jiao, X. Angiotensin II type 1a receptor knockout ameliorates high-fat diet-induced cardiac dysfunction by regulating glucose and lipid metabolism. Acta Biochim. Biophys. Sin. 2023, 55, 1380–1392. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Gwathmey, J.K.; Xie, L.H. Oxidative stress-mediated effects of angiotensin II in the cardiovascular system. World J. Hypertens. 2012, 2, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Egger, M.; Domenighetti, A.A. Adaptive and maladaptive remodeling of cardiomyocyte excitation-contraction coupling by angiotensin II. Trends Cardiovasc. Med. 2010, 20, 78–85. [Google Scholar] [CrossRef]
- Bkaily, G.; Al-Khoury, J.; Simon, Y.; Jacques, D. Intracellular Free Calcium Measurement Using Confocal Imaging. Methods Mol. Biol. 2017, 1527, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Takahahsi, K.; Azuma, M.; Baba, A.; Schaffer, S.; Azuma, J. Taurine improves angiotensin II-induced hypertrophy of cultured neonatal rat heart cells. Adv. Exp. Med. Biol. 1998, 442, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Tain, Y.L.; Hou, C.Y.; Chang-Chien, G.P.; Lin, S.; Hsu, C.N. Protective Role of Taurine on Rat Offspring Hypertension in the Setting of Maternal Chronic Kidney Disease. Antioxidants 2023, 12, 2059. [Google Scholar] [CrossRef] [PubMed]
- Cheng, T.H.; Liu, J.C.; Lin, H.; Shih, N.L.; Chen, Y.L.; Huang, M.T.; Chan, P.; Cheng, C.F.; Chen, J.J. Inhibitory effect of resveratrol on angiotensin II-induced cardiomyocyte hypertrophy. Naunyn Schmiedebergs Arch. Pharmacol. 2004, 369, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Lemmens, K.; Segers, V.F.; Demolder, M.; De Keulenaer, G.W. Role of neuregulin-1/ErbB2 signaling in endothelium-cardiomyocyte cross-talk. J. Biol. Chem. 2006, 281, 19469–19477. [Google Scholar] [CrossRef]
- Abelanet, A.; Camoin, M.; Rubin, S.; Bougaran, P.; Delobel, V.; Pernot, M.; Forfar, I.; Guilbeau-Frugier, C.; Galès, C.; Bats, M.L.; et al. Increased Capillary Permeability in Heart Induces Diastolic Dysfunction Independently of Inflammation, Fibrosis, or Cardiomyocyte Dysfunction. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 745–763. [Google Scholar] [CrossRef]
- Brutsaert, D.L.; Fransen, P.; Andries, L.J.; De Keulenaer, G.W.; Sys, S.U. Cardiac endothelium and myocardial function. Cardiovasc. Res. 1998, 38, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Fort, S.; Lewis, M.J.; Shah, A.M. The role of endocardial endothelium in the modulation of myocardial contraction in the isolated whole heart. Cardioscience 1993, 4, 217–223. [Google Scholar] [PubMed]
- Smiljic, S. The clinical significance of endocardial endothelial dysfunction. Medicina 2017, 53, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bkaily, G.; Abou Abdallah, N.; Simon, Y.; Jazzar, A.; Jacques, D. Vascular smooth muscle remodeling in health and disease. Can. J. Physiol. Pharmacol. 2021, 99, 171–178. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Wang, X.F.; Chen, H.; Yang, N. Antenatal taurine supplementation improves cerebral neurogenesis in fetal rats with intrauterine growth restriction through the PKA-CREB signal pathway. Nutr. Neurosci. 2013, 16, 282–287. [Google Scholar] [CrossRef]
- Fang, Q.; Liu, J.; Chen, L.; Chen, Q.; Ke, J.; Zhang, J.; Liu, Y.; Fu, W. Taurine improves the differentiation of neural stem cells in fetal rats with intrauterine growth restriction via activation of the PKA-CREB-BDNF signaling pathway. Metab. Brain Dis. 2021, 36, 969–981. [Google Scholar] [CrossRef]
- Qaradakhi, T.; Gadanec, L.K.; McSweeney, K.R.; Abraham, J.R.; Apostolopoulos, V.; Zulli, A. The Anti-Inflammatory Effect of Taurine on Cardiovascular Disease. Nutrients 2020, 12, 2847. [Google Scholar] [CrossRef]
- Schaffer, S.; Solodushko, V.; Pastukh, V.; Ricci, C.; Azuma, J. Possible cause of taurine-deficient cardiomyopathy: Potentiation of angiotensin II action. J. Cardiovasc. Pharmacol. 2003, 41, 751–759. [Google Scholar] [CrossRef]
- Azuma, M.; Takahashi, K.; Fukuda, T.; Ohyabu, Y.; Yamamoto, I.; Kim, S.; Iwao, H.; Schaffer, S.W.; Azuma, J. Taurine attenuates hypertrophy induced by angiotensin II in cultured neonatal rat cardiac myocytes. Eur. J. Pharmacol. 2000, 403, 181–188. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacques, D.; Bkaily, G. Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS. Nutrients 2024, 16, 745. https://doi.org/10.3390/nu16050745
Jacques D, Bkaily G. Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS. Nutrients. 2024; 16(5):745. https://doi.org/10.3390/nu16050745
Chicago/Turabian StyleJacques, Danielle, and Ghassan Bkaily. 2024. "Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS" Nutrients 16, no. 5: 745. https://doi.org/10.3390/nu16050745
APA StyleJacques, D., & Bkaily, G. (2024). Taurine Prevents Angiotensin II-Induced Human Endocardial Endothelium Morphological Remodeling and the Increase in Cytosolic and Nuclear Calcium and ROS. Nutrients, 16(5), 745. https://doi.org/10.3390/nu16050745