
Anti-Obesity Effect and Signaling Mechanism of Potassium Poly-γ-Glutamate Produced by Bacillus subtilis Chungkookjang in High-Fat Diet-Induced Obese Mice


 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of PGA-K
2.2. Animals
2.3. Experimental Groups
2.4. Evaluation of Biomarkers in Serum
2.5. Histology
2.6. Statistical Analyses
3. Results
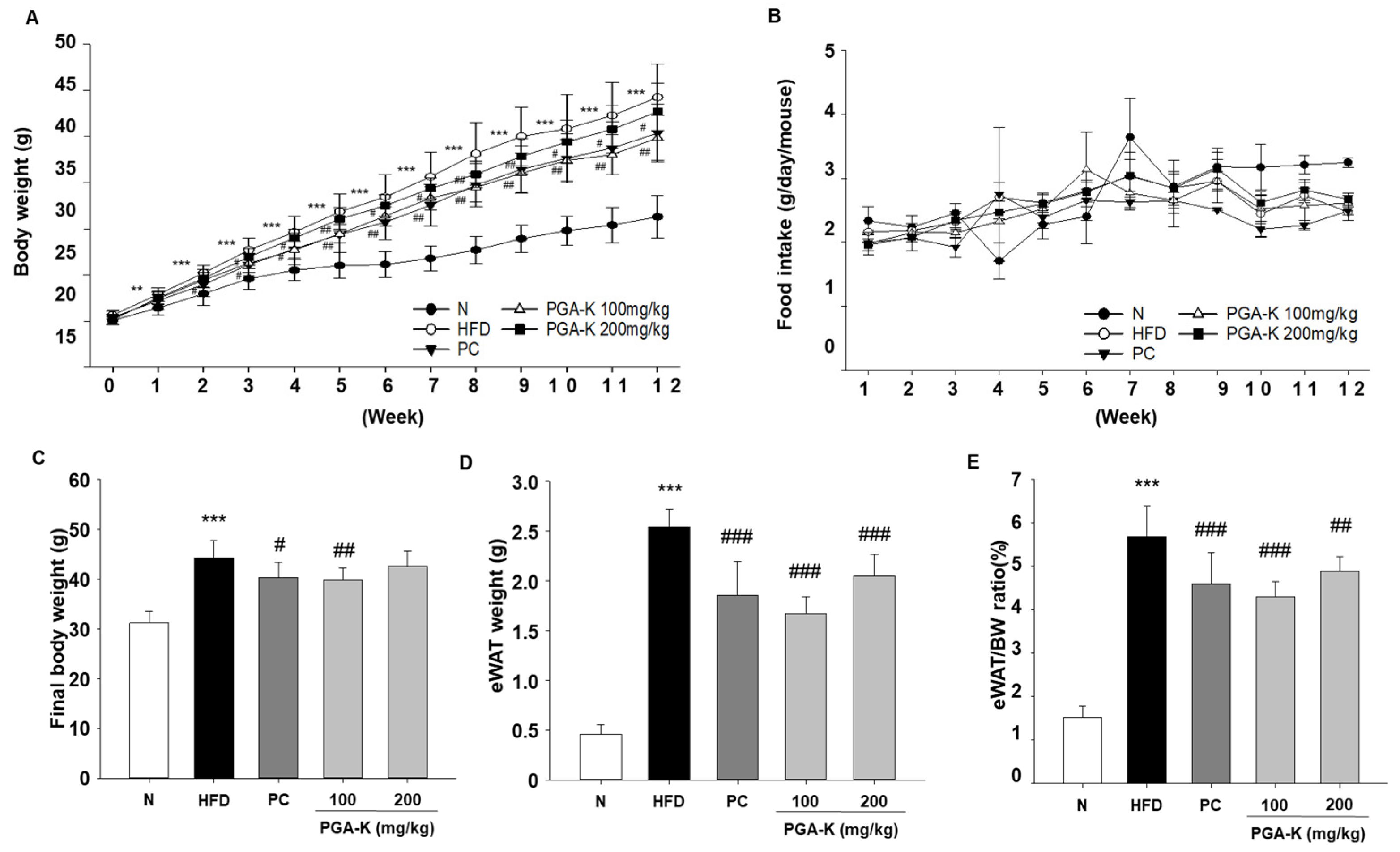
3.1. Effects of PGA-K Administration on Body Weight and Food Intake in HFD-Fed Mice
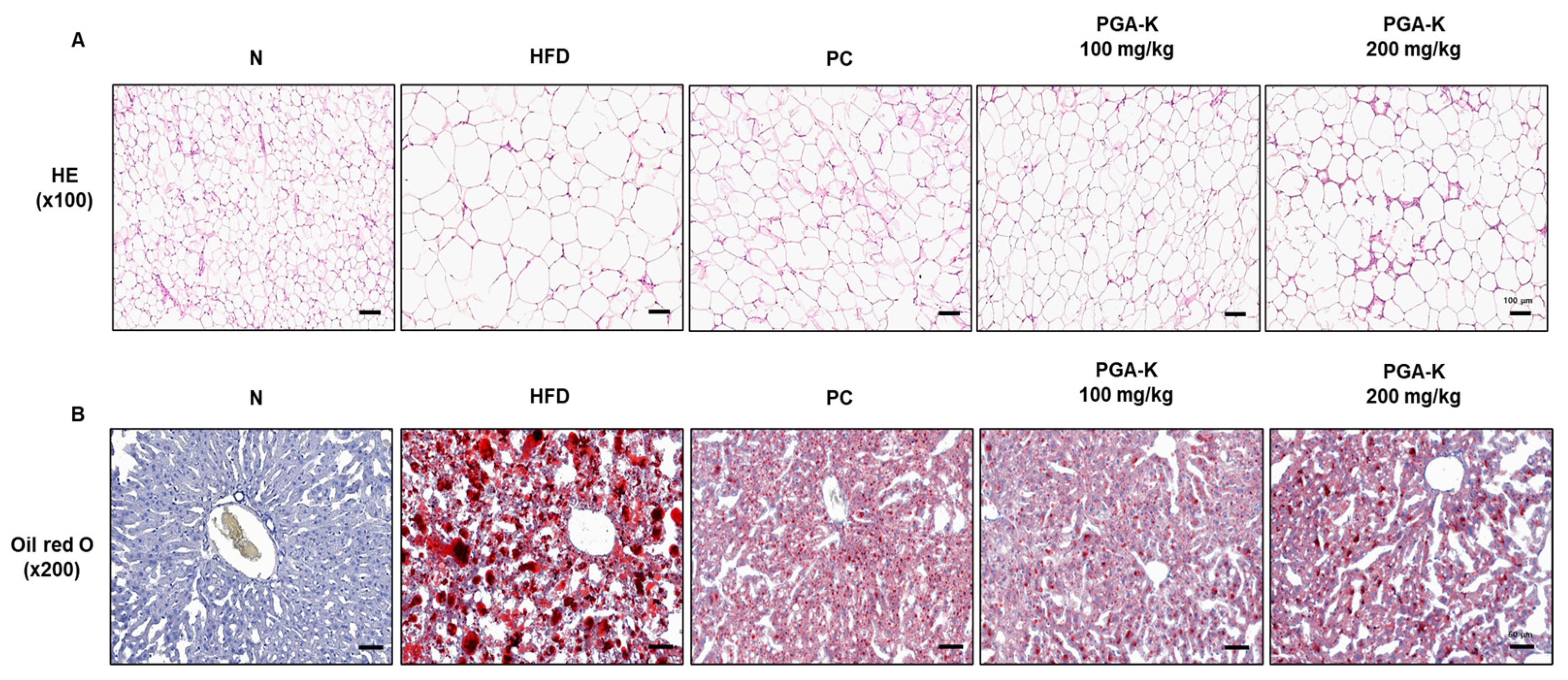
3.2. PGA-K Prevents Adipogenesis and Lipid Accumulation in HFD-Fed Mice
3.3. Effects of PGA-K on Serum Lipid Profiles in HFD-Fed Mice
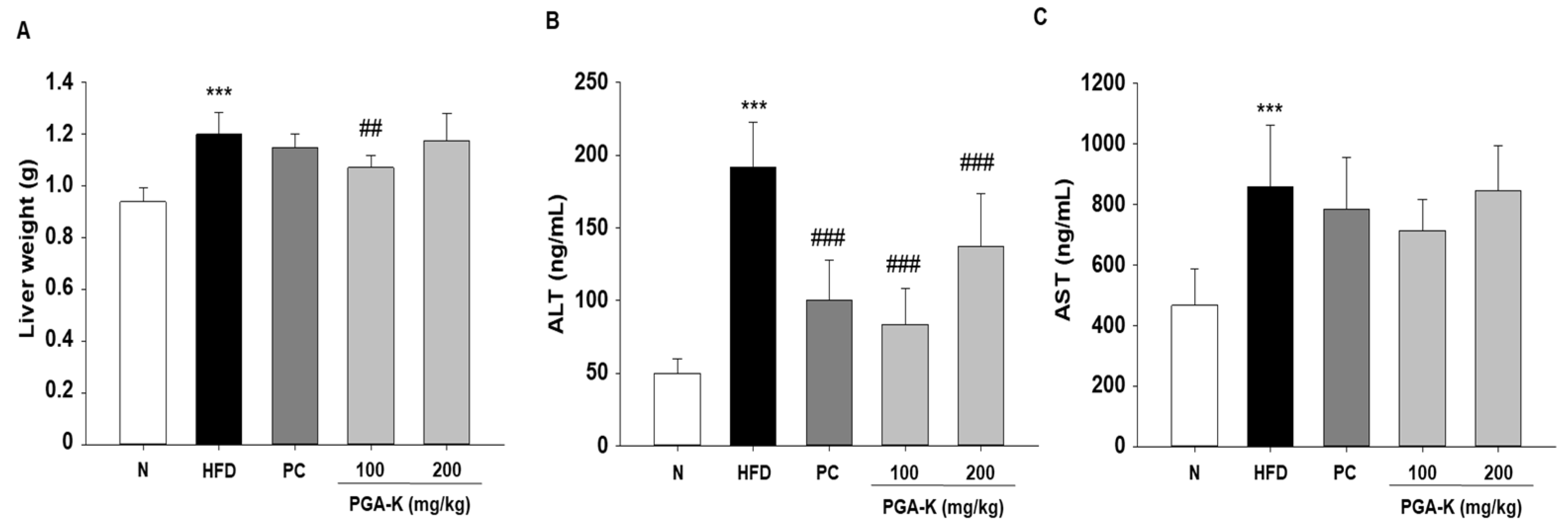
3.4. Effects of PGA-K on Liver Damage in HFD-Fed Mice
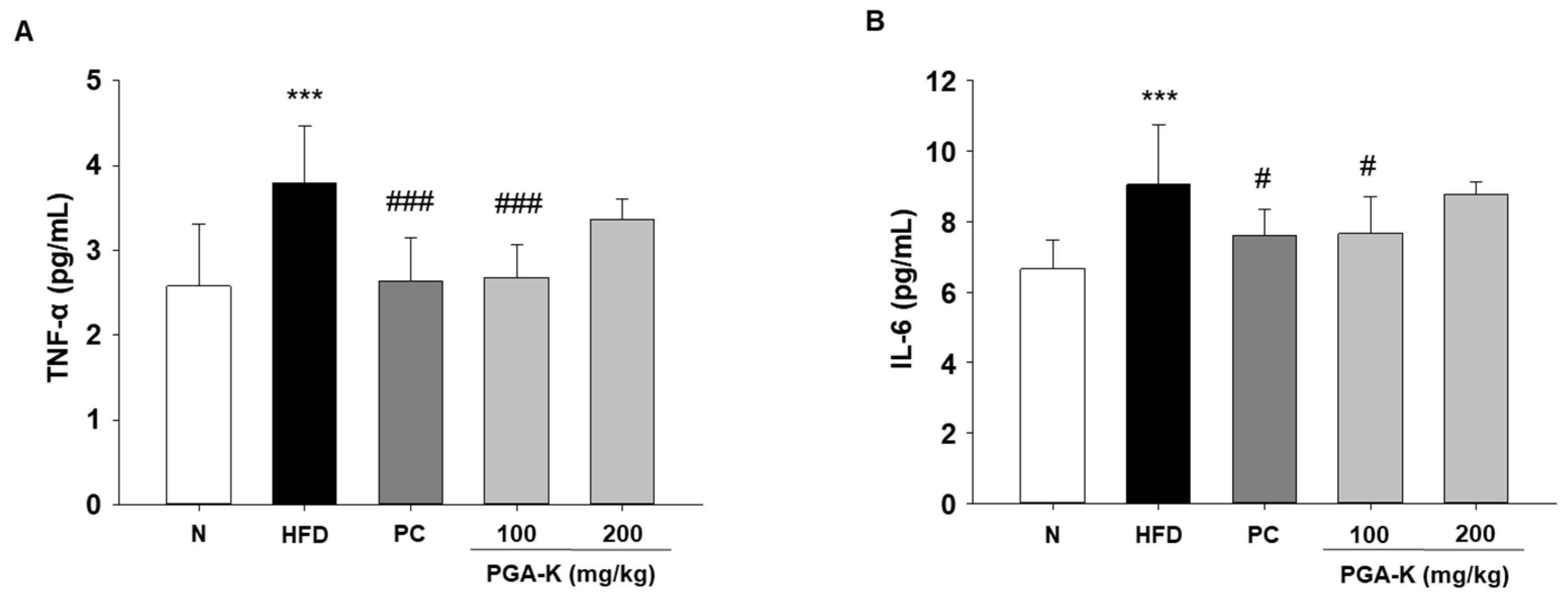
3.5. Effects of PGA-K on HFD-Induced Pro-Inflammatory Cytokines
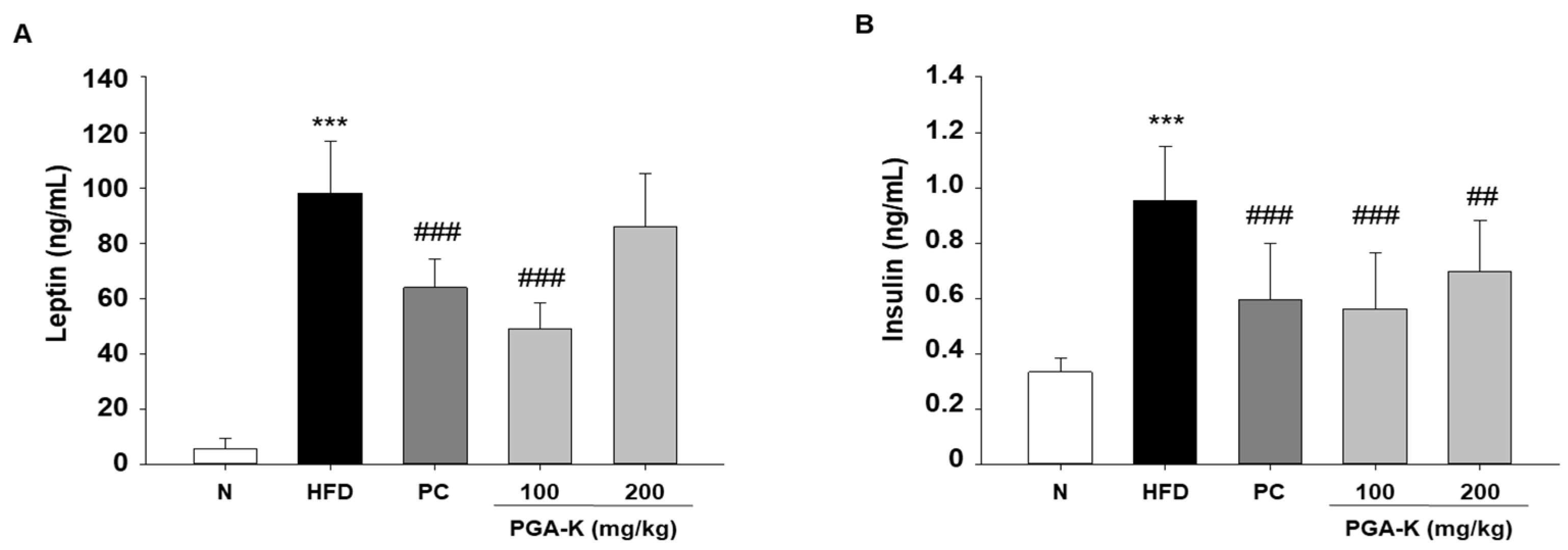
3.6. Effects of PGA-K on HFD-Induced Insulin Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ruze, R.; Liu, T.; Zou, X.; Song, J.; Chen, Y.; Xu, R.; Yin, X.; Xu, Q. Obesity and type 2 diabetes mellitus: Connections in epidemiology, pathogenesis, and treatments. Front. Endocrinol. 2023, 14, 1161521. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, J.E.; Kang, H.M.; Song, H.J.; Kang, N.J.; Hwang, D.Y.; Choi, Y.W. Adiposity Reduction by Cucumis melo var. gaettongchamoe Extract in High-Fat Diet-Induced Obese Mice. Nutrients 2023, 15, 3292. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, A.; Birk, R. Adipose Tissue Hyperplasia and Hypertrophy in Common and Syndromic Obesity-The Case of BBS Obesity. Nutrients 2023, 15, 3445. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, X.; Ibrahim, M.; Peltzer, N. Cell death and inflammation during obesity: “Know my methods, WAT(son)”. Cell Death Differ. 2023, 30, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Cavaliere, G.; Cimmino, F.; Trinchese, G.; Catapano, A.; Petrella, L.; D’Angelo, M.; Lucchin, L.; Mollica, M.P. From Obesity-Induced Low-Grade Inflammation to Lipotoxicity and Mitochondrial Dysfunction: Altered Multi-Crosstalk between Adipose Tissue and Metabolically Active Organs. Antioxidants 2023, 12, 1172. [Google Scholar] [CrossRef] [PubMed]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Gan, R.Y.; Xu, X.Y.; Mao, Q.Q.; Zhang, P.Z.; Li, H.B. Effects and mechanisms of edible and medicinal plants on obesity: An updated review. Crit. Rev. Food Sci. Nutr. 2021, 61, 2061–2077. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, M.S.; Jung, S.; Son, H.Y.; Park, S.; Kang, B.; Kim, S.Y.; Kim, I.H.; Kim, C.T.; Kim, Y. Ginger Extract Ameliorates Obesity and Inflammation via Regulating MicroRNA-21/132 Expression and AMPK Activation in White Adipose Tissue. Nutrients 2018, 10, 1567. [Google Scholar] [CrossRef]
- Pelczyńska, M.; Moszak, M.; Wesołek, A.; Bogdański, P. The Preventive Mechanisms of Bioactive Food Compounds against Obesity-Induced Inflammation. Antioxidants 2023, 12, 1232. [Google Scholar] [CrossRef]
- Al-Mansoori, L.; Al-Jaber, H.; Prince, M.S.; Elrayess, M.A. Role of Inflammatory Cytokines, Growth Factors and Adipokines in Adipogenesis and Insulin Resistance. Inflammation 2022, 45, 31–44. [Google Scholar] [CrossRef]
- Yao, J.; Wu, D.; Qiu, Y. Adipose tissue macrophage in obesity-associated metabolic diseases. Front. Immunol. 2022, 13, 977485. [Google Scholar] [CrossRef]
- Lindhorst, A.; Raulien, N.; Wieghofer, P.; Eilers, J.; Rossi, F.M.V.; Bechmann, I.; Gericke, M. Adipocyte death triggers a pro-inflammatory response and induces metabolic activation of resident macrophages. Cell Death Dis. 2021, 12, 579. [Google Scholar] [CrossRef]
- Li, X.; Ren, Y.; Chang, K.; Wu, W.; Griffiths, H.R.; Lu, S.; Gao, D. Adipose tissue macrophages as potential targets for obesity and metabolic diseases. Front. Immunol. 2023, 14, 1153915. [Google Scholar] [CrossRef] [PubMed]
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented Soy Products and Their Potential Health Benefits: A Review. Microorganisms 2022, 10, 1606. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Hwang, C.W.; Yang, W.S.; Kim, C.H. Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang. Int. J. Mol. Sci. 2021, 22, 5746. [Google Scholar] [CrossRef] [PubMed]
- Gopikrishna, T.; Suresh Kumar, H.K.; Perumal, K.; Elangovan, E. Impact of Bacillus in fermented soybean foods on human health. Ann. Microbiol. 2021, 71, 30. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Yang, Z.; Chen, G.; Liang, Z.; Zeng, W. Enhanced Production of Poly-γ-glutamic Acid by Bacillus subtilis Using Stage-controlled Fermentation and Viscosity Reduction Strategy. Appl. Biochem. Biotechnol. 2024, 196, 1527–1543. [Google Scholar] [CrossRef] [PubMed]
- Afzaal, M.; Saeed, F.; Islam, F.; Ateeq, H.; Asghar, A.; Shah, Y.A.; Ofoedu, C.E.; Chacha, J.S. Nutritional Health Perspective of Natto: A Critical Review. Biochem. Res. Int. 2022, 2022, 5863887. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.H.; Noh, H.; Kim, H.W.; Cho, S.Y.; Kim, H.J.; Lee, S.H.; Lee, S.H.; Gunter, M.J.; Ferrari, P.; Scalbert, A.; et al. Metabolic tracking of isoflavones in soybean products and biosamples from healthy adults after fermented soybean consumption. Food Chem. 2020, 330, 127317. [Google Scholar] [CrossRef]
- Kim, I.S.; Yang, W.S.; Kim, C.H. Beneficial Effects of Soybean-Derived Bioactive Peptides. Int. J. Mol. Sci. 2021, 22, 8570. [Google Scholar] [CrossRef]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.P.; Sulaiman Rahman, H. Antioxidant and Oxidative Stress: A Mutual Interplay in Age-Related Diseases. Front. Pharmacol. 2018, 9, 1162. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Song, J.-L.; Wang, Q.; Qian, Y.; Li, G.-J.; Pang, L. Comparisons of Shuidouchi, Natto, and Cheonggukjang in their physicochemical properties, and antimutagenic and anticancer effects. Food Sci. Biotechnol. 2013, 22, 1077–1084. [Google Scholar] [CrossRef]
- Fuentes, E.; Fuentes, F.; Vilahur, G.; Badimon, L.; Palomo, I. Mechanisms of chronic state of inflammation as mediators that link obese adipose tissue and metabolic syndrome. Mediat. Inflamm. 2013, 2013, 136584. [Google Scholar] [CrossRef]
- Kirichenko, T.V.; Markina, Y.V.; Bogatyreva, A.I.; Tolstik, T.V.; Varaeva, Y.R.; Starodubova, A.V. The Role of Adipokines in Inflammatory Mechanisms of Obesity. Int. J. Mol. Sci. 2022, 23, 14982. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Ellulu, M.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Wondmkun, Y.T. Obesity, insulin resistance, and type 2 diabetes: Associations and therapeutic implications. Diabetes Metab. Syndr. Obes. 2020, 13, 3611–3616. [Google Scholar] [CrossRef]
- Kojta, I.; Chacińska, M.; Błachnio-Zabielska, A. Obesity, Bioactive Lipids, and Adipose Tissue Inflammation in Insulin Resistance. Nutrients 2020, 12, 1305. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2019, 10, 1607. [Google Scholar] [CrossRef]
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose Tissue Remodeling: Its Role in Energy Metabolism and Metabolic Disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef]
- Stadler, J.T.; Marsche, G. Obesity-Related Changes in High-Density Lipoprotein Metabolism and Function. Int. J. Mol. Sci. 2020, 21, 8985. [Google Scholar] [CrossRef] [PubMed]
- Chakhtoura, M.; Haber, R.; Ghezzawi, M.; Rhayem, C.; Tcheroyan, R.; Mantzoros, C.S. Pharmacotherapy of obesity: An update on the available medications and drugs under investigation. EClinicalMedicine 2023, 58, 101882. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.G.; Park, C.Y. Anti-Obesity Drugs: A Review about Their Effects and Safety. Diabetes Metab. J. 2012, 36, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.J.; Lee, S.Y. Anti-Obesity Drugs: Long-Term Efficacy and Safety: An Updated Review. World J. Mens Health 2021, 39, 208–221. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Huang, J.; Zhou, J.; Zhi, C.; Bai, Y.; Che, Q.; Cao, H.; Guo, J.; Su, Z. Anti-Obesity Effect and Mechanism of Chitooligosaccharides Were Revealed Based on Lipidomics in Diet-Induced Obese Mice. Molecules 2023, 28, 5595. [Google Scholar] [CrossRef]
- Farias, T.C.; Abreu, J.P.; Oliveira, J.P.S.; Macedo, A.F.; Rodríguez-Vega, A.; Tonin, A.P.; Cardoso, F.S.N.; Meurer, E.C.; Koblitz, M.G.B. Bioactive properties of peptide fractions from Brazilian soy protein hydrolysates. In silico evaluation and experimental evidence. Food Hydrocoll. Health 2023, 3, 100112. [Google Scholar] [CrossRef]
- Bhathena, S.J.; Velasquez, M.T. Beneficial role of dietary phytoestrogens in obesity and diabetes. Am. J. Clin. Nutr. 2002, 76, 1191–1201. [Google Scholar] [CrossRef]
- Chi, X.X.; Zhang, T.; Zhang, D.J.; Yu, W.; Wang, Q.Y.; Zhen, J.L. Effects of isoflavones on lipid and apolipoprotein levels in patients with type 2 diabetes in Heilongjiang Province in China. J. Clin. Biochem. Nutr. 2016, 59, 134–138. [Google Scholar] [CrossRef]
- Ioniță-Mîndrican, C.B.; Ziani, K.; Mititelu, M.; Oprea, E.; Neacșu, S.M.; Moroșan, E.; Dumitrescu, D.E.; Roșca, A.C.; Drăgănescu, D.; Negrei, C. Therapeutic Benefits and Dietary Restrictions of Fiber Intake: A State of the Art Review. Nutrients 2022, 14, 2641. [Google Scholar] [CrossRef]
- Leeuwendaal, N.K.; Stanton, C.; O’Toole, P.W.; Beresford, T.P. Fermented Foods, Health and the Gut Microbiome. Nutrients 2022, 14, 1527. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Gębalski, J.; Gołębiewski, J.; Malinowski, B. Probiotics for the Treatment of Overweight and Obesity in Humans-A Review of Clinical Trials. Microorganisms 2020, 8, 1148. [Google Scholar] [CrossRef]
- Einarson, T.R.; Acs, A.; Ludwig, C.; Panton, U.H. Prevalence of cardiovascular disease in type 2 diabetes: A systematic literature review of scientific evidence from across the world in 2007–2017. Cardiovasc. Diabetol. 2018, 17, 83. [Google Scholar] [CrossRef]
- Akhlaghi, M.; Zare, M.; Nouripour, F. Effect of Soy and Soy Isoflavones on Obesity-Related Anthropometric Measures: A Systematic Review and Meta-analysis of Randomized Controlled Clinical Trials. Adv. Nutr. 2017, 8, 705–717. [Google Scholar] [CrossRef]
- Byun, M.S.; Yu, O.K.; Cha, Y.S.; Park, T.S. Korean traditional Chungkookjang improves body composition, lipid profiles and atherogenic indices in overweight/obese subjects: A double-blind, randomized, crossover, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 2016, 70, 1116–1122. [Google Scholar] [CrossRef]
- Langa, S.; Peirotén, Á.; Curiel, J.A.; de la Bastida, A.R.; Landete, J.M. Isoflavone Metabolism by Lactic Acid Bacteria and Its Application in the Development of Fermented Soy Food with Beneficial Effects on Human Health. Foods 2023, 12, 1293. [Google Scholar] [CrossRef]
- Saad, B. A Review of the Anti-Obesity Effects of Wild Edible Plants in the Mediterranean Diet and Their Active Compounds: From Traditional Uses to Action Mechanisms and Therapeutic Targets. Int. J. Mol. Sci. 2023, 24, 12641. [Google Scholar] [CrossRef]
- Ogunleye, A.; Bhat, A.; Irorere, V.U.; Hill, D.; Williams, C.; Radecka, I. Poly-γ-glutamic acid: Production, properties and applications. Microbiology 2015, 161, 1–17. [Google Scholar] [CrossRef]
- Balogun-Agbaje, O.A.; Odeniyi, O.A.; Odeniyi, M.A. Drug delivery applications of poly-γ-glutamic acid. Future J. Pharm. Sci. 2021, 7, 125. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Q.; Wang, R.; Sun, K.; Li, S.; Lin, G.; Lei, P.; Xu, H. Effect of dietary poly-γ-glutamic acid on growth, digestive enzyme activity, antioxidant capacity, and TOR pathway gene expression of gibel carp (Carassius auratus gibelio). Aquac. Rep. 2022, 27, 101412. [Google Scholar] [CrossRef]
- Zorena, K.; Jachimowicz-Duda, O.; Ślęzak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Wang, X. The Role of Gut Microbiota in High-Fat-Diet-Induced Diabetes: Lessons from Animal Models and Humans. Nutrients 2023, 15, 922. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Laps (Weeks) | PGA-K 100 mg/kg | HFD | Difference in Means (SE; 95%CI) | p Value |
|---|---|---|---|---|
| Baseline | 20.26 ± 0.53 | 20.66 ± 0.43 | ||
| 1 | 2.17 ± 0.23 | 2.25 ± 0.27 | −0.08 (0.36, −0.84 to 0.68) | 0.82 |
| 2 | 4.20 ± 0.32 | 4.55 ± 0.36 | −0.36 (0.48, −1.93 to 0.68) | 0.47 |
| 3 | 6.02 ± 0.42 | 7.05 ± 0.52 | −1.03 (0.66, −2.46 to 0.40 | 0.14 |
| 4 | 7.46 ± 0.37 | 8.99 ± 0.65 | −1.53 (0.75, −3.14 to 0.08) | 0.06 |
| 5 | 9.2 ± 0.43 | 11.2 ± 0.72 | −1.20 (0.84, −3.8 to −0.19) | 0.03 |
| 6 | 11.14 ± 0.44 | 12.8 ± 0.90 | −1.66 (1.0, −3.81 to 0.49) | 0.12 |
| 7 | 13.08 ± 0.51 | 14.99 ± 0.97 | −1.91 (1.10, −4.26 to 0.45) | 0.10 |
| 8 | 14.24 ± 0.62 | 17.48 ± 1.18 | −3.24 (1.33, −6.1 to −0.38) | 0.03 |
| 9 | 15.78 ± 0.89 | 19.34 ± 1.11 | −3.57 (−6.62, 0.51 to −0.51) | 0.03 |
| 10 | 17.16 ± 0.96 | 20.19 ± 1.27 | −3.03 (1.59, −6.44 to 0.38) | 0.08 |
| 11 | 17.79 ± 0.94 | 21.62 ± 1.21 | −3.84 (1.54, −7.13 to −0.54) | 0.03 |
| 12 | 19.60 ± 1.02 | 23.60 ± 1.20 | −4.00 (1.58, −7.39 to −0.60) | 0.02 |
| Efficacy Parameters | PGA-K 100 mg/kg | HFD | Difference in Means | p Value |
|---|---|---|---|---|
| TG | 56.88 ± 1.43 | 71.61 ± 4.30 | −14.74 | <0.001 |
| TC | 66.52 ± 1.30 | 95.31 ± 5.02 | −28.79 | <0.001 |
| HDL | 14.41 ± 1.11 | 23.12 ± 1.70 | −8.71 | <0.001 |
| LDL | 56.21 ± 3.17 | 83.44 ± 3.69 | −27.23 | <0.001 |
| eWAT weight | 1.67 ± 0.60 | 2.54 ± 0.06 | −0.87 | <0.001 |
| eWAT/BW (%) | 4.29 ± 0.12 | 5.68 ± 0.25 | −1.39 | <0.001 |
| Liver weight | 1.07 ± 0.02 | 1.20 ± 0.03 | −0.13 | 0.002 |
| ALT | 83.49 ± 8.84 | 191.83 ± 11 | −108.34 | <0.001 |
| AST | 714.69 ± 36.40 | 860.42 ± 71.86 | −145.73 | 0.09 |
| TNF-a | 2.68 ± 0.14 | 3.80 ± 0.24 | −1.12 | <0.001 |
| IL-6 | 7.67 ± 0.37 | 9.05 ± 0.60 | −1.38 | 0.017 |
| Leptin | 49.08 ± 3.22 | 97.99 ± 6.70 | −48.91 | <0.001 |
| Insulin | 0.56 ± 0.07 | 0.96 ± 0.07 | −0.39 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-H.; Choi, J.; Park, J.Y.; Kim, H.-R.; Shim, M.; Im, K.; Choe, H.; Choi, J.-C.; Park, Y.-C.; Lim, T.-G.; et al. Anti-Obesity Effect and Signaling Mechanism of Potassium Poly-γ-Glutamate Produced by Bacillus subtilis Chungkookjang in High-Fat Diet-Induced Obese Mice. Nutrients 2024, 16, 809. https://doi.org/10.3390/nu16060809
Lee S-H, Choi J, Park JY, Kim H-R, Shim M, Im K, Choe H, Choi J-C, Park Y-C, Lim T-G, et al. Anti-Obesity Effect and Signaling Mechanism of Potassium Poly-γ-Glutamate Produced by Bacillus subtilis Chungkookjang in High-Fat Diet-Induced Obese Mice. Nutrients. 2024; 16(6):809. https://doi.org/10.3390/nu16060809
Chicago/Turabian StyleLee, Seung-Hyeon, Jiwon Choi, Jae Young Park, Ha-Rim Kim, Myeongkuk Shim, Kyunghyun Im, Hyeonjeong Choe, Jae-Chul Choi, Young-Chul Park, Tae-Gyu Lim, and et al. 2024. "Anti-Obesity Effect and Signaling Mechanism of Potassium Poly-γ-Glutamate Produced by Bacillus subtilis Chungkookjang in High-Fat Diet-Induced Obese Mice" Nutrients 16, no. 6: 809. https://doi.org/10.3390/nu16060809
APA StyleLee, S.-H., Choi, J., Park, J. Y., Kim, H.-R., Shim, M., Im, K., Choe, H., Choi, J.-C., Park, Y.-C., Lim, T.-G., Seo, H., Jang, H., Oh, B.-J., Kim, S.-Y., & Park, M. H. (2024). Anti-Obesity Effect and Signaling Mechanism of Potassium Poly-γ-Glutamate Produced by Bacillus subtilis Chungkookjang in High-Fat Diet-Induced Obese Mice. Nutrients, 16(6), 809. https://doi.org/10.3390/nu16060809