
Ginsenoside Rg5 Activates the LKB1/AMPK/mTOR Signaling Pathway and Modifies the Gut Microbiota to Alleviate Nonalcoholic Fatty Liver Disease Induced by a High-Fat Diet

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Fecal Microbiota Transplantation
2.4. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT) Iochemical Parameter Analysis
2.5. Biochemical Parameter Analysis
2.6. Histological Analysis
2.7. Western Blotting
2.8. 16S rRNA Sequencing
2.9. Detection of SCFAs Content in Mouse Feces
2.10. Statistical Analysis
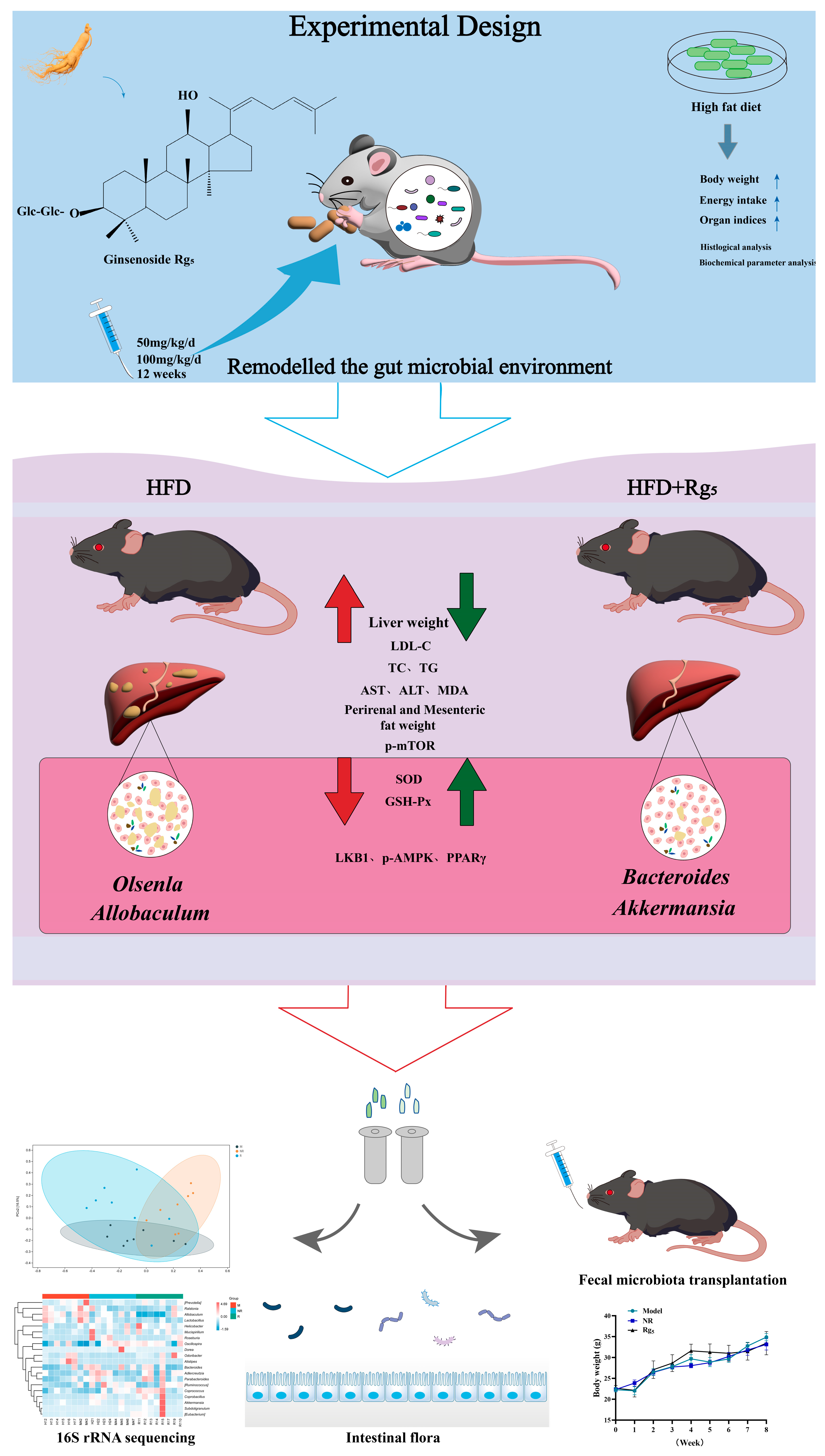
3. Results
3.1. Effects of Rg5 Intervention on Body Weight, Food Intake, and Organ Indices in NAFLD Mice
3.2. Effects of Rg5 on the Phenotype and Protein Expression in NAFLD Mice
3.3. Effects of Rg5 Intervention on the Intestinal Homeostasis of NAFLD Mice
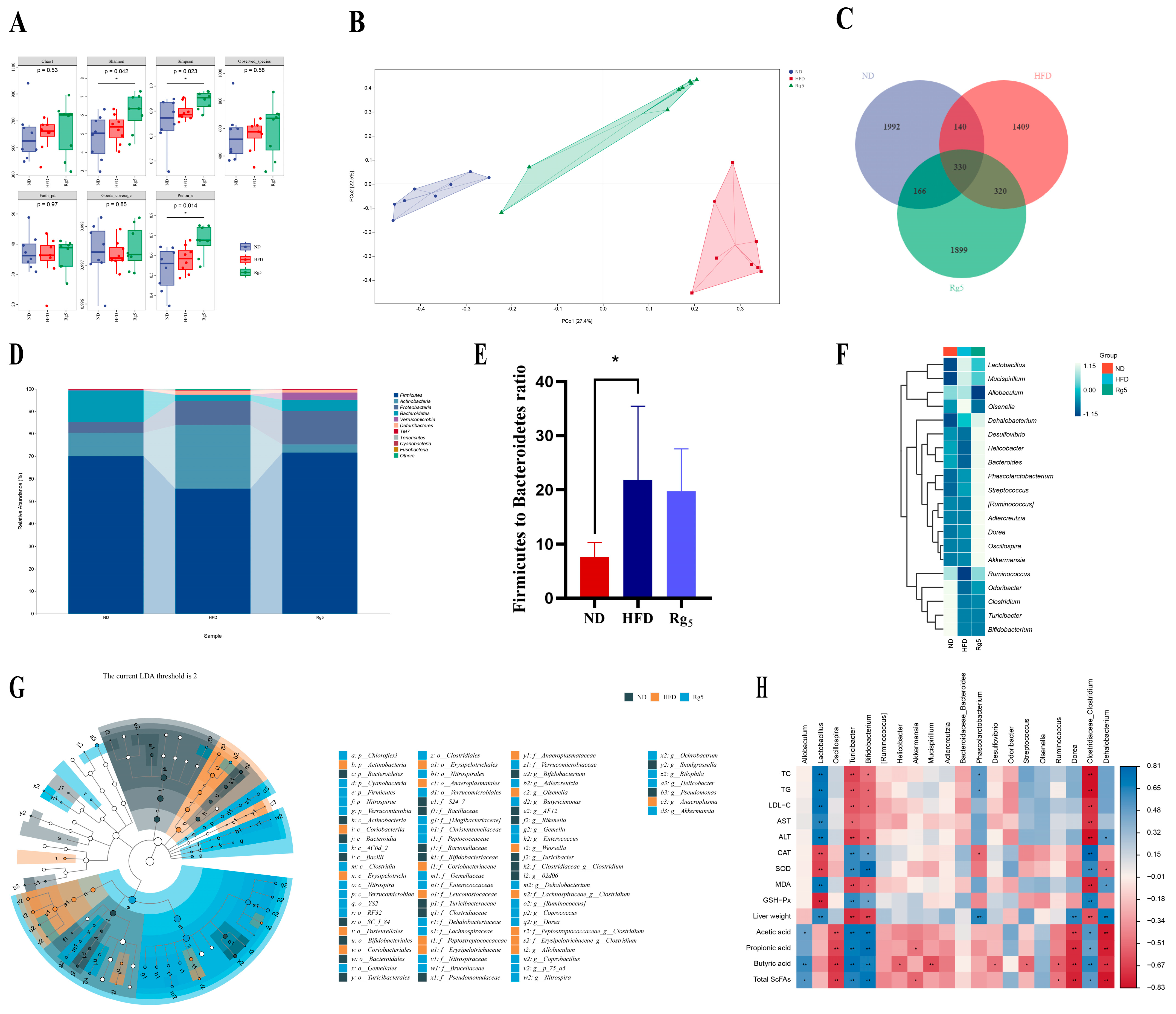
3.4. Effects of Rg5 on Gut Microbiota Communities of NAFLD Mice
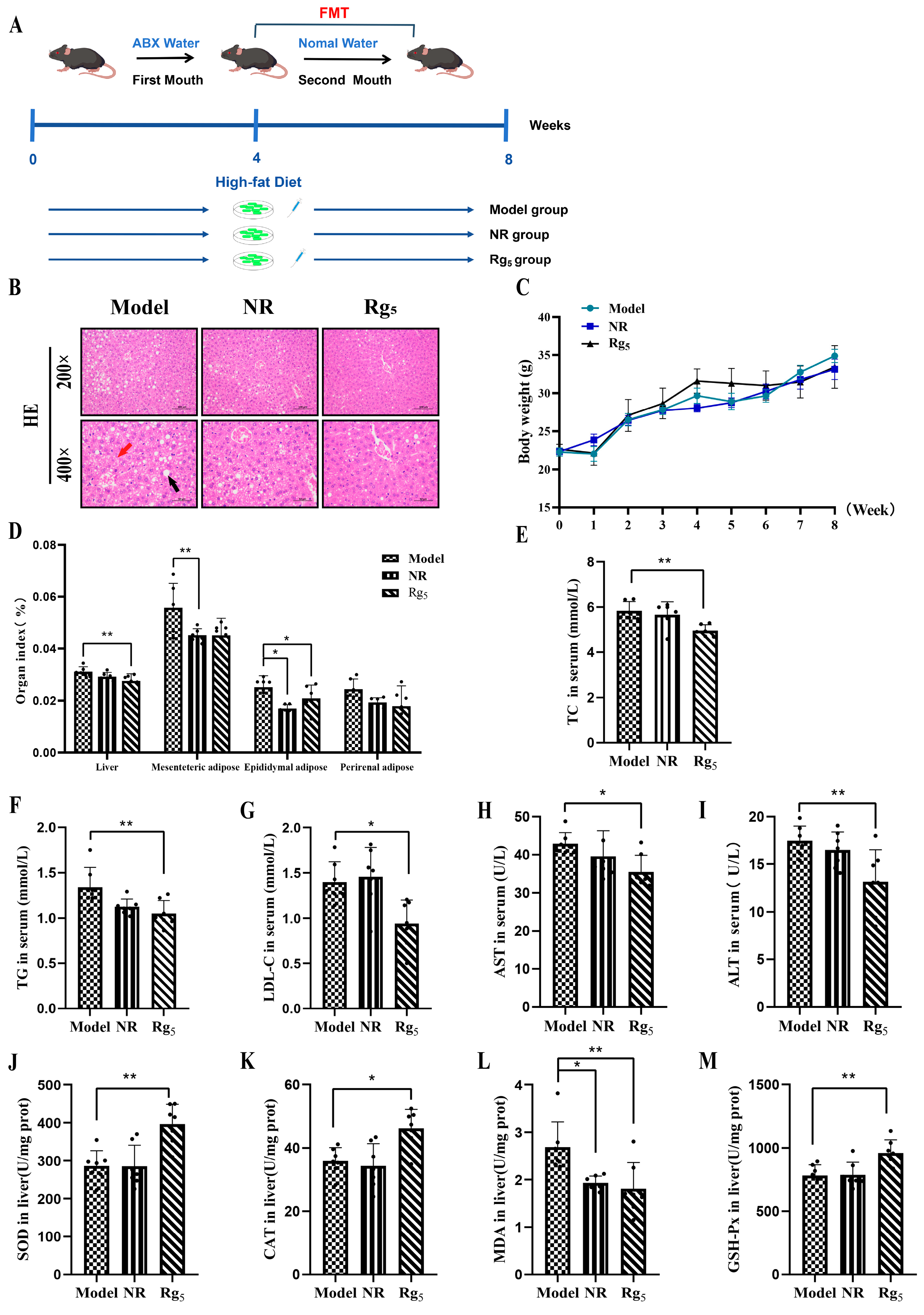
3.5. Effect of FMT on the Phenotype in NAFLD Mice
3.6. Effect of FMT on Intestinal Bacterial Communities in NAFLD Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Powell, E.E.; Wong, V.W.S.; Rinella, M. Non-alcoholic fatty liver disease. Lancet 2021, 397, 2212–2224. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD development and therapeutic strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Youssefian, L.; Vahidnezhad, H.; Saeidian, A.H.; Pajouhanfar, S.; Sotoudeh, S.; Mansouri, P.; Amirkashani, D.; Zeinali, S.; Levine, M.A.; Peris, K.; et al. Inherited non-alcoholic fatty liver disease and dyslipidemia due to monoallelic ABHD5 mutations. J. Hepatol. 2019, 71, 366–370. [Google Scholar] [CrossRef]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.G.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. 2020, 17, 279–297. [Google Scholar] [CrossRef]
- Liu, J.; Wu, A.; Cai, J.; She, Z.-G.; Li, H. The contribution of the gut-liver axis to the immune signaling pathway of NAFLD. Front. Immunol. 2022, 13, 968799. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef]
- Roselli, M.; Natella, F.; Zinno, P.; Guantario, B.; Canali, R.; Schifano, E.; De Angelis, M.; Nikoloudaki, O.; Gobbetti, M.; Perozzi, G.; et al. Colonization ability and impact on hu-man gut microbiota of foodborne microbes from traditional or probiotic-added fermented foods: A systematic review. Front. Nutr. 2021, 8, 689084. [Google Scholar] [CrossRef] [PubMed]
- Vijay, A.; Valdes, A.M. Role of the gut microbiome in chronic diseases: A narrative review. Eur. J. Clin. Nutr. 2022, 76, 489–501. [Google Scholar] [CrossRef]
- Su, Y.; Li, J.; Wu, L.; Kuang, H. Polysaccharides from TCM herbs exhibit potent anti-obesity effect by mediating the community structure of gut microbiota. Pharmazie 2021, 76, 473–479. [Google Scholar]
- Wu, Z.; Tian, E.; Chen, Y.; Dong, Z.; Peng, Q. Gut microbiota and its roles in the pathogenesis and therapy of en-do-crine system diseases. Microbiol. Res. 2023, 268, 127291. [Google Scholar] [CrossRef]
- Blaak, E.E.; Canfora, E.E.; Theis, S.; Frost, G.; Groen, A.K.; Mithieux, G.; Nauta, A.; Scott, K.; Stahl, B.; Van Harsselaar, J.; et al. Short chain fatty acids in human gut and metabolic health. Benef. Microbes 2020, 11, 411–455. [Google Scholar] [CrossRef]
- Chen, C.; Lv, Q.; Li, Y.; Jin, Y.H. The Anti-Tumor Effect and Underlying Apoptotic Mechanism of Ginsenoside Rk1 and Rg5 in Human Liver Cancer Cells. Molecules 2021, 26, 3926. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, X.; Yang, X.; Yu, H.; Bu, M.; Fu, J.; Zhang, Z.; Xu, H.; Hu, J.; Lu, J.; et al. Revitalizing myocarditis treatment through gut microbiota modulation: Unveiling a promising therapeutic avenue. Front. Cell. Infect. Microbiol. 2023, 13, 1191936. [Google Scholar] [CrossRef] [PubMed]
- Gao, P.; Zheng, M.; Lu, H.; Lu, S. The progressive utilization of ponkan peel residue for regulating human gut microbiota through sequential extraction and modification of its dietary fibers. Foods 2023, 12, 4148. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Lu, J.; Cheng, S.; Zou, X.; Tao, Q.; Wang, M.; Wang, N.; Zheng, L.; Liao, W.; Li, Y.; et al. Periodontitis salivary microbiota exacerbates colitis-induced anxiety-like behavior via gut microbiota. Npj Biofilms Microbiomes 2023, 9, 93. [Google Scholar] [CrossRef]
- Bai, Z.; Huang, X.; Wu, G.; Ye, H.; Huang, W.; Nie, Q.; Chen, H.; Yin, J.; Chen, Y.; Nie, S. Polysaccharides from red kidney bean alleviating hyperglycemia and hyperlipidemia in type 2 diabetic rats via gut microbiota and lipid metabolic modulation. Food Chem. 2023, 404 Pt A, 134598. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, in-teractive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2′s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Ren, D.D.; Chen, K.C.; Li, S.S.; Zhang, Y.T.; Li, Z.M.; Liu, S.; Sun, Y.S. Panax quinquefolius polysaccharides ameliorate ulcerative colitis in mice induced by dextran sulfate sodium. Front. Immunol. 2023, 14, 1161625. [Google Scholar] [CrossRef]
- Zheng, Z.; Lyu, W.; Ren, Y.; Li, X.; Zhao, S.; Yang, H.; Xiao, Y. Allobaculum involves in the modulation of intestinal ANGPTLT4 expression in mice treated by high-fat diet. Front. Nutr. 2021, 8, 690138. [Google Scholar] [CrossRef]
- Zhang, D.; Jian, Y.P.; Zhang, Y.N.; Li, Y.; Gu, L.T.; Sun, H.H.; Liu, M.D.; Zhou, H.L.; Wang, Y.S.; Xu, Z.X. Short-chain fatty acids in diseases. Cell Commun. Signal. CCS 2023, 21, 212. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Jiang, Y.; Wang, M.; Melaku, M.; Liu, L.; Zhao, Y.; Everaert, N.; Yi, B.; Zhang, H. Intestinal dysbiosis in nonalcoholic fatty liver disease (NAFLD): Focusing on the gut-liver axis. Crit. Rev. Food Sci. Nutr. 2023, 63, 1689–1706. [Google Scholar] [CrossRef] [PubMed]
- Everard, A.; Lazarevic, V.; Gaïa, N.; Johansson, M.; Ståhlman, M.; Backhed, F.; Delzenne, N.M.; Schrenzel, J.; François, P.; Cani, P.D. Microbiome of prebiotic-treated mice reveals novel targets involved in host response during obesity. ISME J. 2014, 8, 2116–2130. [Google Scholar] [CrossRef] [PubMed]
- Bao, T.; He, F.; Zhang, X.; Zhu, L.; Wang, Z.; Lu, H.; Wang, T.; Li, Y.; Yang, S.; Wang, H. Inulin exerts beneficial effects on non-alcoholic fatty liver disease via modulating gut microbiome and suppressing the lipopolysaccha-ride-toll-like receptor 4-Mψ-nuclear factor-κB-nod-Like receptor protein 3 pathway via gut-liver axis in mice. Front. Pharmacol. 2020, 11, 558525. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Li, Z.; Chen, W.; Rong, T.; Wang, G.; Ma, X. Microbiome-metabolomics analysis investigating the impacts of dietary starch types on the composition and metabolism of colonic microbiota in finishing pigs. Front. Microbiol. 2019, 10, 1143. [Google Scholar] [CrossRef]
- Csala, A.; Hof, M.H.; Zwinderman, A.H. Multiset sparse redundancy analysis for high-dimensional omics data. Biom. J. 2019, 61, 406–423. [Google Scholar] [CrossRef]
- Xie, Z.; Du, J.; Gan, M.; Zhou, C.; Li, M.; Liu, C.; Wang, M.; Chen, L.; Zhao, Y.; Wang, Y. Short-term dietary choline supplementation alters the gut microbiota and liver metabolism of finishing pigs. Front. Microbiol. 2023, 14, 1266042. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease-Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Jin, W.; Li, C.; Yang, S.; Song, S.; Hou, W.; Song, Y.; Du, Q. Hypolipidemic effect and molecular mechanism of ginsenosides: A review based on oxidative stress. Front. Pharmacol. 2023, 14, 1166898. [Google Scholar] [CrossRef]
- Liang, W.; Zhou, K.; Jian, P.; Chang, Z.; Zhang, Q.; Liu, Y.; Xiao, S.; Zhang, L. Ginsenosides improve nonalcoholic fatty liver disease via integrated regulation of gut microbiota, inflammation and energy homeostasis. Front. Pharmacol. 2021, 12, 622841. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhu, C.; Yang, H.; Deng, J.; Fan, D. Protective effect of ginsenoside Rg5 against kidney injury via inhibition of NLRP3 inflammasome activation and the MAPK signaling pathway in high-fat diet/streptozotocin-induced diabetic mice. Pharmacol. Res. 2020, 155, 104746. [Google Scholar] [CrossRef]
- Liu, M.Y.; Liu, F.; Gao, Y.L.; Yin, J.N.; Yan, W.Q.; Liu, J.G.; Li, H.J. Pharmacological activities of ginsenoside Rg5 (Review). Exp. Ther. Med. 2021, 22, 840. [Google Scholar] [CrossRef]
- Xiao, N.; Lou, M.D.; Lu, Y.T.; Yang, L.L.; Liu, Q.; Liu, B.; Qi, L.W.; Li, P. Ginsenoside Rg5 attenuates hepatic glucagon response via suppression of succinate-associated HIF-1α induction in HFD-fed mice. Diabetologia 2017, 60, 1084–1093. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, H.; Zhu, C.; Deng, J.; Fan, D. Hypoglycemic Effect of Ginsenoside Rg5 Mediated Partly by Modulating Gut Microbiota Dysbiosis in Diabetic db/db Mice. J. Agric. Food Chem. 2020, 68, 5107–5117. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhu, C.; Fu, R.; Ma, X.; Duan, Z.; Fan, D. Ginsenoside Rg5 inhibits lipid accumulation and hepatocyte apoptosis via the Notch1 signaling pathway in NASH mice. Phytomedicine 2024, 124, 155287. [Google Scholar] [CrossRef] [PubMed]
- Garcia, D.; Shaw, R.J. AMPK: Mechanisms of cellular energy sensing and restoration of metabolic balance. Mol. Cell 2017, 66, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Xu, Z.; Wang, Y.; Kuang, S.; Shan, T. Adipocyte-specific DKO of Lkb1 and mTOR protects mice against HFD-induced obesity, but results in insulin resistance. J. Lipid Res. 2018, 59, 974–981. [Google Scholar] [CrossRef] [PubMed]
- Panwar, V.; Singh, A.; Bhatt, M.; Tonk, R.K.; Azizov, S.; Raza, A.S.; Sengupta, S.; Kumar, D.; Garg, M. Multifaceted role of mTOR (mammalian target of rapamycin) signaling pathway in human health and disease. Signal Transduct. Target. Ther. 2023, 8, 375. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Zhou, Y.; Tsao, R.; Dong, H.; Zhang, H. Amelioratory effect of resistant starch on non-alcoholic fatty liver disease via the gut-liver axis. Front. Nutr. 2022, 9, 861854. [Google Scholar] [CrossRef]
- Zhang, Q.; Xing, W.; Wang, Q.; Tang, Z.; Wang, Y.; Gao, W. Gut microbiota-mitochondrial inter-talk in nonalcoholic fatty liver disease. Front. Nutr. 2022, 9, 934113. [Google Scholar] [CrossRef]
- Hrncir, T.; Hrncirova, L.; Kverka, M.; Hromadka, R.; Machova, V.; Trckova, E.; Kostovcikova, K.; Kralickova, P.; Krejsek, J.; Tlaskalova-Hogenova, H. Gut Microbiota and NAFLD: Pathogenetic Mechanisms, Microbiota Signatures, and Therapeutic Interventions. Microorganisms 2021, 9, 957. [Google Scholar] [CrossRef]
- Plaza-Díaz, J.; Solis-Urra, P.; Aragón-Vela, J.; Rodríguez-Rodríguez, F.; Olivares-Arancibia, J.; Álvarez-Mercado, A.I. Insights into the Impact of Microbiota in the Treatment of NAFLD/NASH and Its Potential as a Biomarker for Prognosis and Diagnosis. Biomedicines 2021, 9, 145. [Google Scholar] [CrossRef]
- Rodríguez-Daza, M.C.; Pulido-Mateos, E.C.; Lupien-Meilleur, J.; Guyonnet, D.; Desjardins, Y.; Roy, D. Polyphenolmediated gut microbiota modulation: Toward prebiotics and further. Front. Nutr. 2021, 8, 689456. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Li, Y.; Han, H.; Ma, J.; Liu, G.; Wu, X.; Huang, X.; Fang, R.; Baba, K.; Bin, P. Administration of exogenous melatonin improves the diurnal rhythms of the gut microbiota in mice fed a high-fat diet. mSystems 2020, 5, e00002-20. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira-A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef]
- Carvalho, B.M.; Guadagnini, D.; Tsukumo, D.M.L.; Schenka, A.A.; Latuf-Filho, P.; Vassallo, J.; Dias, J.C.; Kubota, L.T.; Carvalheira, J.B.C.; Saad, M.J.A. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 2012, 55, 2823–2834. [Google Scholar] [CrossRef] [PubMed]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Ak-ker-mansia muciniphila in overweight and obese human volunteers: A proof-of-concept exploratory study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- van Muijlwijk, G.H.; van Mierlo, G.; Jansen, P.W.T.C.; Vermeulen, M.; Bleumink-Pluym, N.M.C.; Palm, N.W.; van Putten, J.P.M.; de Zoete, M.R. Identification of Allobaculum mucolyticum as a novel human intestinal mucin degrader. Gut Microbes 2021, 13, 1966278. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Sheng, L.; Zhong, J.; Tao, X.; Zhu, W.; Ma, J.; Yan, J.; Zhao, A.; Zheng, X.; Wu, G. Desulfovibrio vulgaris, a potent acetic acid-producing bacterium, attenuates nonalcoholic fatty liver disease in mice. Gut Microbes 2021, 13, 1930874. [Google Scholar] [CrossRef]
- Zikou, E.; Dovrolis, N.; Dimosthenopoulos, C.; Gazouli, M.; Makrilakis, K. The effect of probiotic supplements on metabolic parameters of people with type 2 diabetes in greece-a randomized, double-blind, placebo-controlled study. Nutrients 2023, 15, 4663. [Google Scholar] [CrossRef]
- Cotillard, A.; Kennedy, S.P.; Kong, L.C.; Prifti, E.; Pons, N.; Le Chatelier, E.; Almeida, M.; Quinquis, B.; Levenez, F.; Galleron, N.; et al. Dietary intervention impact on gut microbial gene richness. Nature 2013, 500, 585–588. [Google Scholar] [CrossRef]
- Xue, L.; Deng, Z.; Luo, W.; He, X.; Chen, Y. Effect of fecal microbiota transplantation on non-alcoholic fatty liver disease: A randomized clinical trial. Front. Cell. Infect. Microbiol. 2022, 12, 759306. [Google Scholar] [CrossRef]
- Tariq, R.; Syed, T.; Yadav, D.; Prokop, L.J.; Singh, S.; Loftus, E.V., Jr.; Pardi, D.S.; Khanna, S. Outcomes of fecal microbiota transplantation for C. difficile infection in inflammatory bowel disease: A systematic review and meta-analysis. J. Clin. Gastroenterol. 2023, 57, 285–293. [Google Scholar] [CrossRef]
- Coia, H.; Ma, N.; Hou, Y.; Dyba, M.D.; Fu, Y.; Cruz, M.I.; Benitez, C.; Graham, G.T.; McCutcheon, J.N.; Zheng, Y.L.; et al. Prevention of Lipid Peroxidation-derived Cyclic DNA Adduct and Mutation in High Fat Diet-induced Hepatocarcinogenesis by Theaphenon E. Cancer Prev. Res. 2018, 11, 665–676. [Google Scholar] [CrossRef]
- Guo, J.; Pereira, T.J.; Dalvi, P.; Yeung, L.S.N.; Swain, N.; Breen, D.M.; Lam, L.; Dolinsky, V.W.; Giacca, A. High-dose metformin (420 mg/kg daily p.o.) increases insulin sensitivity but does not affect neointimal thickness in the rat carotid balloon injury model of restenosis. Metabolism 2016, 68, 108–118. [Google Scholar] [CrossRef]
- Norris, G.H.; Porter, C.M.; Jiang, C.; Millar, C.L.; Blesso, C.N. Dietary sphingomyelin attenuates hepatic steatosis and adipose tissue inflammation in high-fat-diet-induced obese mice. J. Nutr. Biochem. 2017, 40, 36–43. [Google Scholar] [CrossRef]
- Tordoff, M.G.; Aleman, T.R.; Murphy, M.C. No effects of monosodium glutamate consumption on the body weight or composition of adult rats and mice. Physiol. Behav. 2012, 107, 338–345. [Google Scholar] [CrossRef]
- Lee, Y.J.; Ko, E.H.; Kim, J.E.; Kim, E.; Lee, H.; Choi, H.; Yu, J.H.; Kim, H.J.; Seong, J.K.; Kim, K.S.; et al. Nuclear receptor PPARγ-regulated monoacylglycerol O-acyltransferase 1 (MGAT1) expression is responsible for the lipid accumulation in diet-induced hepatic steatosis. Proc. Natl. Acad. Sci. USA 2012, 109, 13656–13661. [Google Scholar] [CrossRef]
- Perry, R.J.; Resch, J.M.; Douglass, A.M.; Madara, J.C.; Rabin-Court, A.; Kucukdereli, H.; Wu, C.; Song, J.D.; Lowell, B.B.; Shulman, G.I. Leptin’s hunger-suppressing effects are mediated by the hypothalamic–pituitary–adrenocortical axis in rodents. ESPE Yearb. Paediatr. Endocrinol. 2020, 17. [Google Scholar] [CrossRef]
- Miranda, C.L.; Johnson, L.A.; de Montgolfier, O.; Elias, V.D.; Ullrich, L.S.; Hay, J.J.; Paraiso, I.L.; Choi, J.; Reed, R.L.; Revel, J.S.; et al. Non-estrogenic Xanthohumol Derivatives Mitigate Insulin Resistance and Cognitive Impairment in High-Fat Diet-induced Obese Mice. Sci. Rep. 2018, 8, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.J.; Bernier, M.; Aon, M.A.; Cortassa, S.; Kim, E.Y.; Fang, E.F.; Palacios, H.H.; Ali, A.; Navas-Enamorado, I.; Di Francesco, A.; et al. Nicotinamide Improves Aspects of Healthspan, but Not Lifespan, in Mice. Cell Metab. 2018, 27, 667–676.e4. [Google Scholar] [CrossRef] [PubMed]
- Kopec, A.K.; Abrahams, S.R.; Thornton, S.; Palumbo, J.S.; Mullins, E.S.; Divanovic, S.; Weiler, H.; Owens, A.P., 3rd; Mackman, N.; Goss, A.; et al. Thrombin promotes diet-induced obesity through fibrin-driven inflammation. J. Clin. Investig. 2017, 127, 3152–3166. [Google Scholar] [CrossRef]
- Yang, J.W.; Kim, H.S.; Im, J.H.; Kim, J.W.; Jun, D.W.; Lim, S.C.; Lee, K.; Choi, J.M.; Kim, S.K.; Kang, K.W. GPR119: A promising target for nonalcoholic fatty liver disease. FASEB J. 2016, 30, 324–335. [Google Scholar] [CrossRef]
- Miyao, M.; Kotani, H.; Ishida, T.; Kawai, C.; Manabe, S.; Abiru, H.; Tamaki, K. Pivotal role of liver sinusoidal endothelial cells in NAFLD/NASH progression. Mod. Pathol. 2015, 95, 1130–1144. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, Y.; Chen, J.; Qu, D.; Sun, Q.; Yu, Y.; Zhang, H.; Liu, Z.; Sha, J.; Sun, Y. Ginsenoside Rg5 Activates the LKB1/AMPK/mTOR Signaling Pathway and Modifies the Gut Microbiota to Alleviate Nonalcoholic Fatty Liver Disease Induced by a High-Fat Diet. Nutrients 2024, 16, 842. https://doi.org/10.3390/nu16060842
Shi Y, Chen J, Qu D, Sun Q, Yu Y, Zhang H, Liu Z, Sha J, Sun Y. Ginsenoside Rg5 Activates the LKB1/AMPK/mTOR Signaling Pathway and Modifies the Gut Microbiota to Alleviate Nonalcoholic Fatty Liver Disease Induced by a High-Fat Diet. Nutrients. 2024; 16(6):842. https://doi.org/10.3390/nu16060842
Chicago/Turabian StyleShi, Yingying, Jianbo Chen, Di Qu, Qiang Sun, Yang Yu, Hao Zhang, Zhengbo Liu, Jiyue Sha, and Yinshi Sun. 2024. "Ginsenoside Rg5 Activates the LKB1/AMPK/mTOR Signaling Pathway and Modifies the Gut Microbiota to Alleviate Nonalcoholic Fatty Liver Disease Induced by a High-Fat Diet" Nutrients 16, no. 6: 842. https://doi.org/10.3390/nu16060842