
A Gnotobiotic Mouse Model with Divergent Equol-Producing Phenotypes: Potential for Determining Microbial-Driven Health Impacts of Soy Isoflavone Daidzein
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
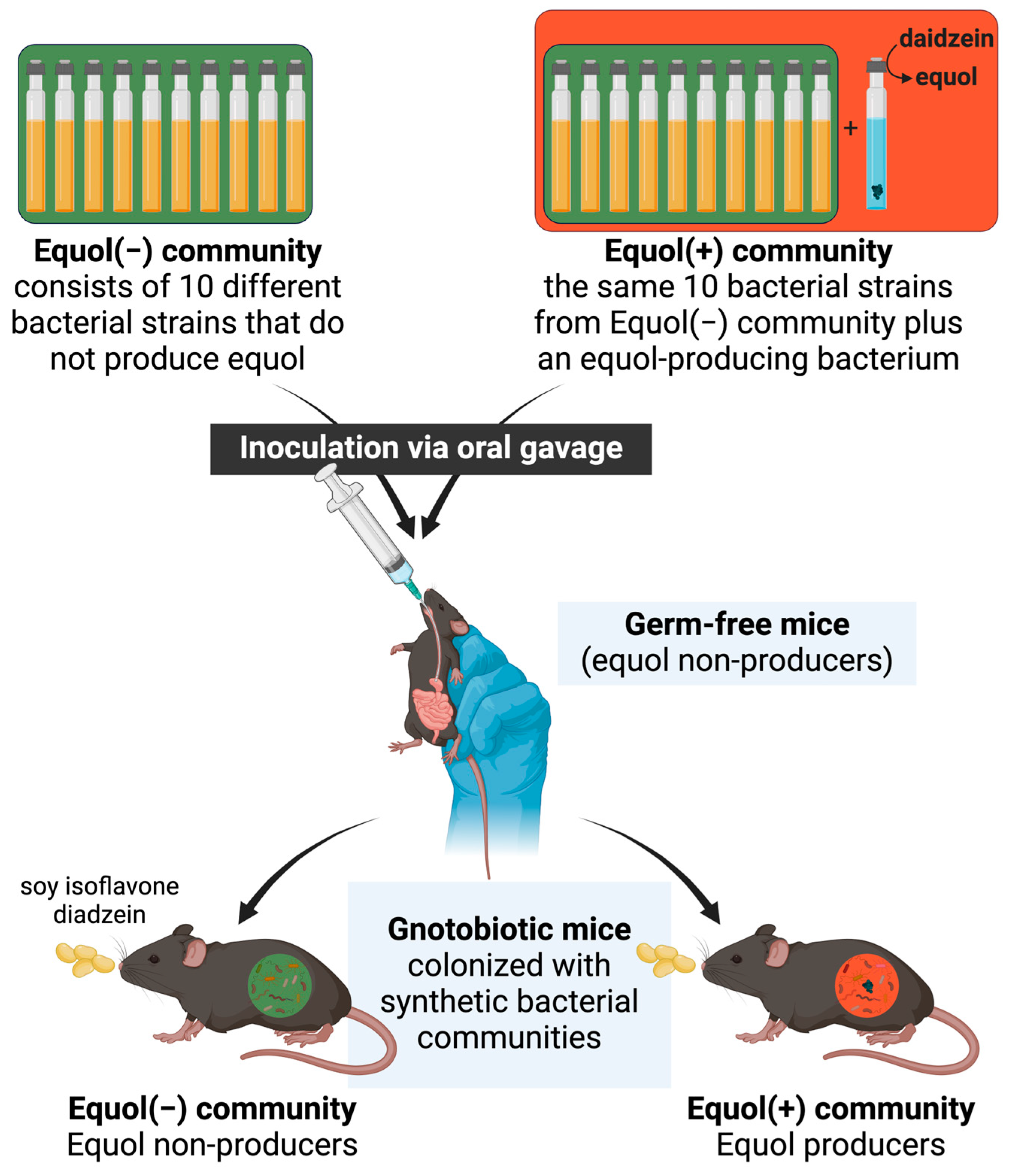
2.1. Design of Synthetic Bacterial Communities
2.2. Gnotobiotic Mouse Study
2.3. Quantification of Daidzein and Equol
2.4. Microbiota Analysis
2.5. Strain-Specific qPCRs
2.6. Cecal Short Chain Fatty Acids
2.7. Statistical Analysis
3. Results
3.1. Body Weight, Mesenteric Fat, and Gonadal Fat Mass Did Not Differ in Mice Based on the Microbiota Received
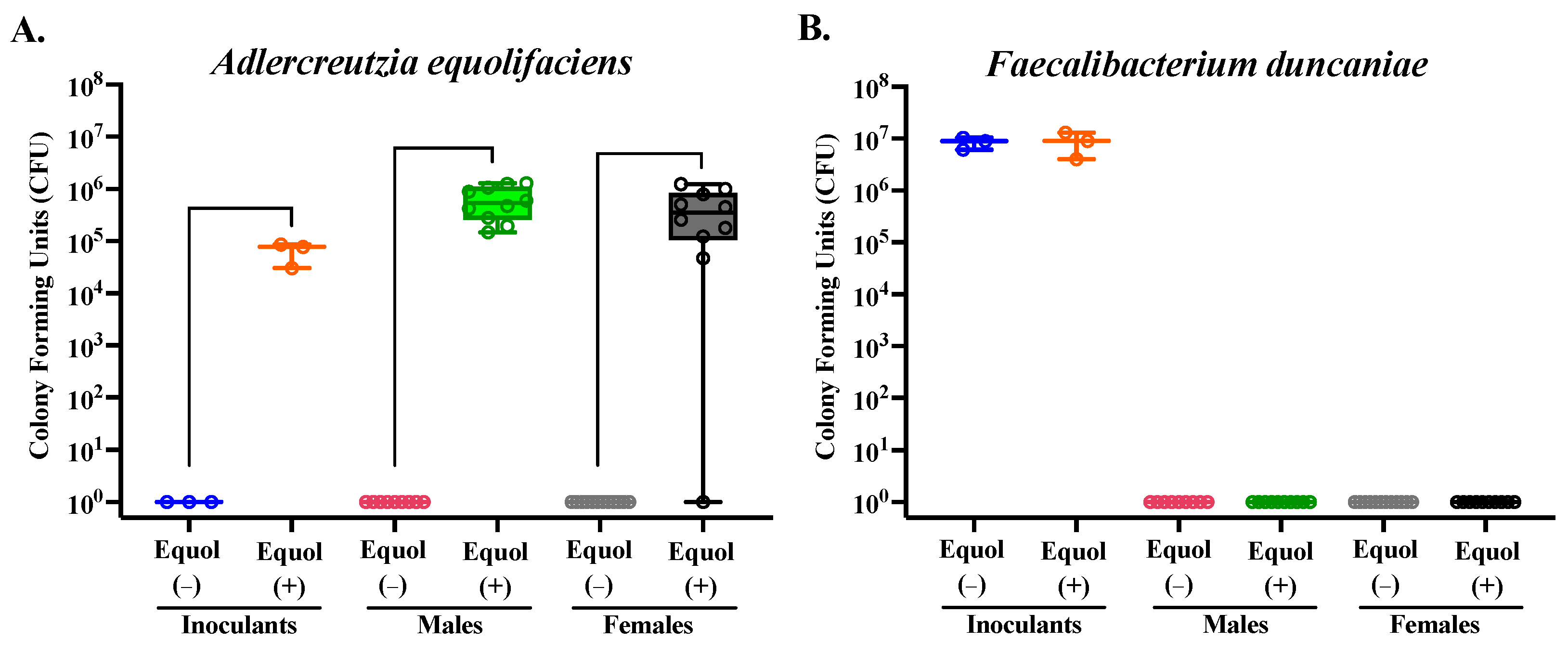
3.2. The Synthetic Bacterial Communities Produce Equol-Producing Capacity as Designed in Gnotobiotic Mice Fed a High Daidzein Diet
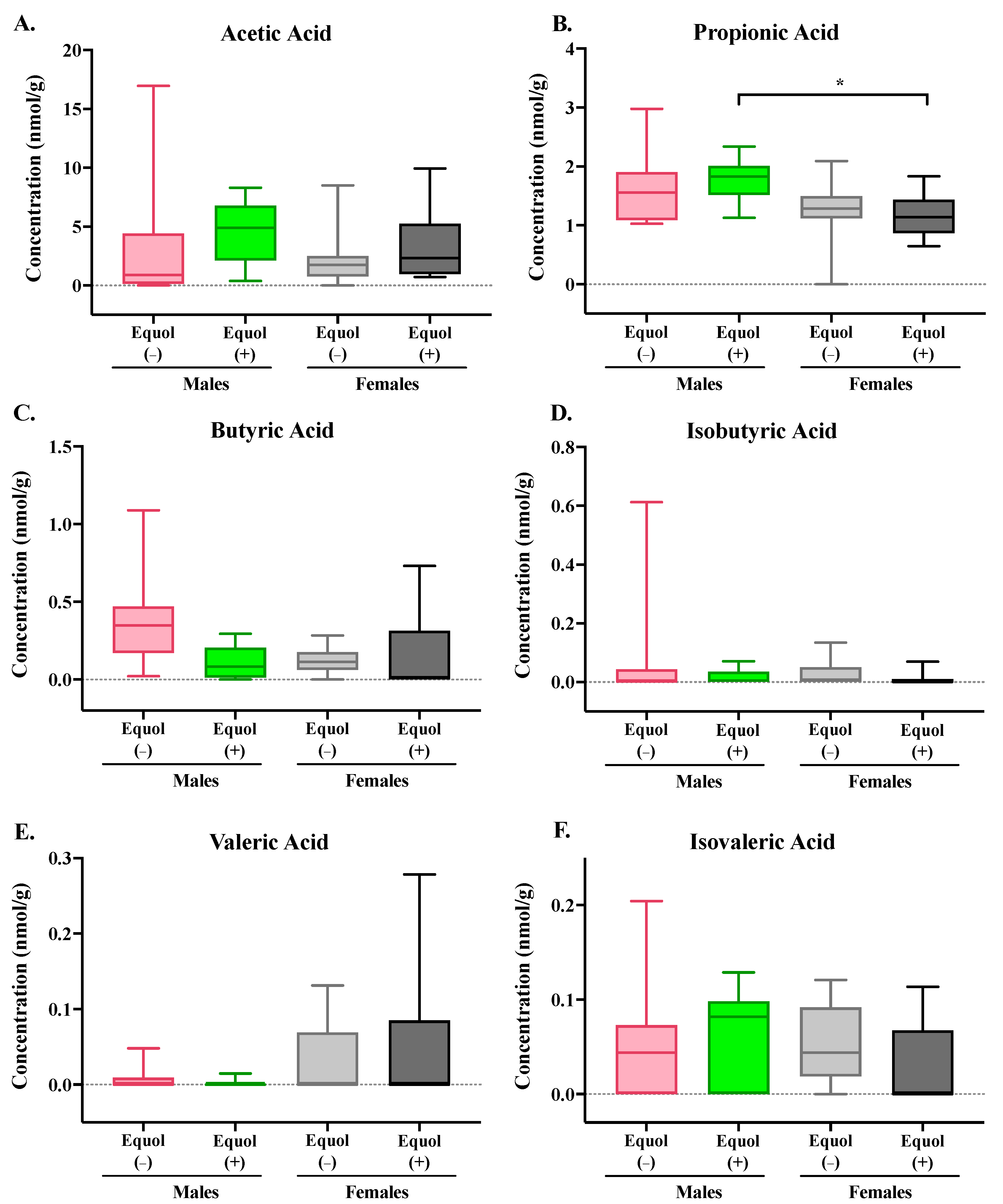
3.3. Equol-Producing Status Did Not Affect the Concentration of Short-Chain Fatty Acids in the Cecum of Gnotobiotic Mice
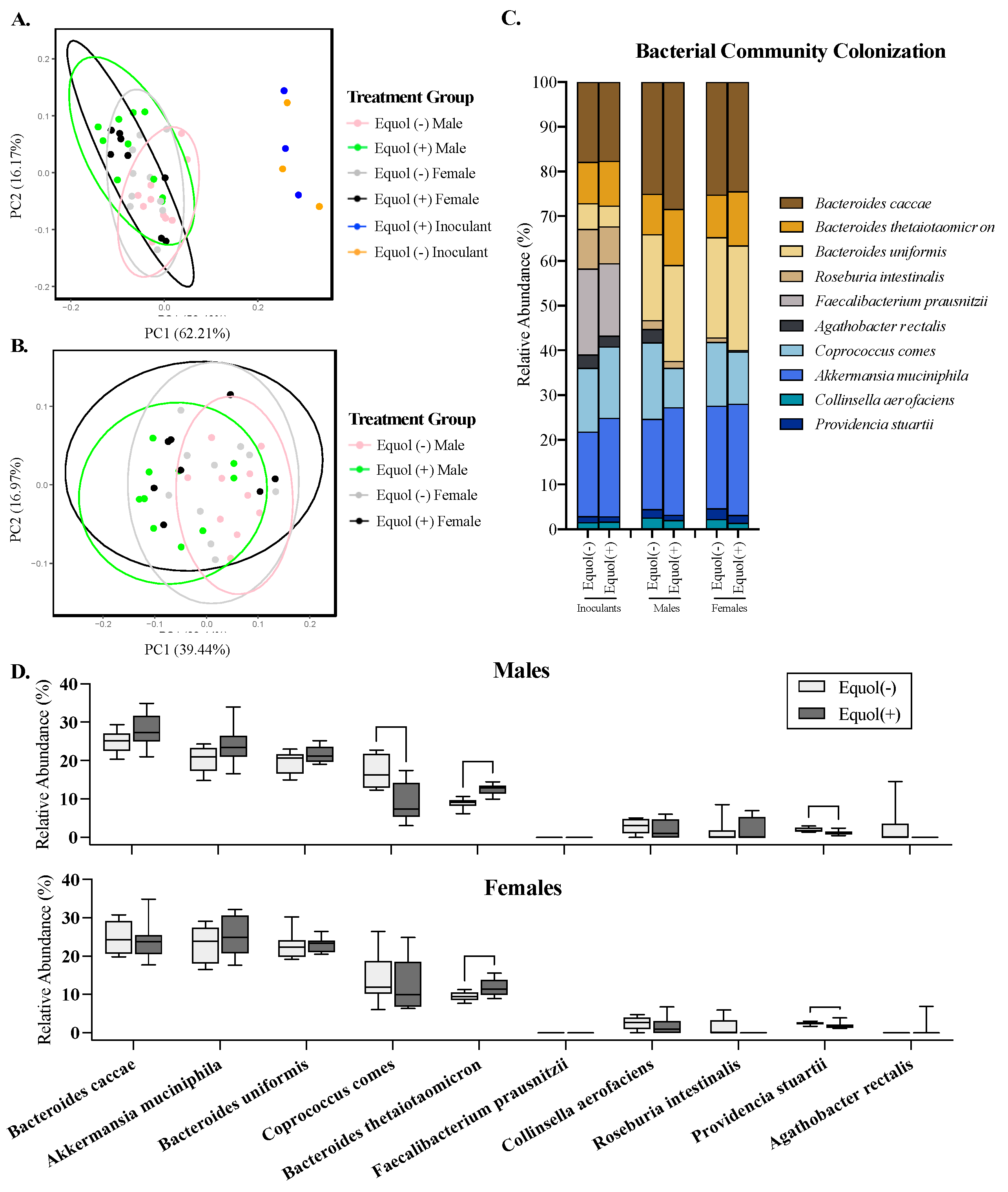
3.4. Community-Level Assessment of the Gut Microbiota Revealed Differences between the Bacterial Inoculants and Mouse Gut, but Similarity Exists among Treatment Groups in Mice
3.5. The Relative Abundance of Some Bacterial Taxa in Mice Colonized with Equol(−) and Equol(+) Differ Significantly Statistically despite the Similarity Observed in the Overall Community Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salas-Huetos, A.; Bulló, M.; Salas-Salvadó, J. Dietary Patterns, Foods and Nutrients in Male Fertility Parameters and Fecundability: A Systematic Review of Observational Studies. Hum. Reprod. Update 2017, 23, 371–389. [Google Scholar] [CrossRef] [PubMed]
- Cederroth, C.R.; Zimmermann, C.; Beny, J.-L.; Schaad, O.; Combepine, C.; Descombes, P.; Doerge, D.R.; Pralong, F.P.; Vassalli, J.-D.; Nef, S. Potential Detrimental Effects of a Phytoestrogen-Rich Diet on Male Fertility in Mice. Mol. Cell. Endocrinol. 2010, 321, 152–160. [Google Scholar] [CrossRef]
- Nagata, C.; Takatsuka, N.; Kawakami, N.; Shimizu, H. Soy Product Intake and Hot Flashes in Japanese Women: Results from a Community-Based Prospective Study. Am. J. Epidemiol. 2001, 153, 790–793. [Google Scholar] [CrossRef]
- Zhang, Y.-B.; Chen, W.-H.; Guo, J.-J.; Fu, Z.-H.; Yi, C.; Zhang, M.; Na, X.-L. Soy Isoflavone Supplementation Could Reduce Body Weight and Improve Glucose Metabolism in Non-Asian Postmenopausal Women—A Meta-Analysis. Nutr. Burbank Los Angel. Cty. Calif 2013, 29, 8–14. [Google Scholar] [CrossRef]
- Setchell, K.D. Phytoestrogens: The Biochemistry, Physiology, and Implications for Human Health of Soy Isoflavones. Am. J. Clin. Nutr. 1998, 68, 1333S–1346S. [Google Scholar] [CrossRef]
- Yoshikata, R.; Myint, K.Z.; Ohta, H. Relationship between Equol Producer Status and Metabolic Parameters in 743 Japanese Women: Equol Producer Status Is Associated with Antiatherosclerotic Conditions in Women around Menopause and Early Postmenopause. Menopause 2017, 24, 216–224. [Google Scholar] [CrossRef]
- Acharjee, S.; Zhou, J.-R.; Elajami, T.K.; Welty, F.K. Effect of Soy Nuts and Equol Status on Blood Pressure, Lipids and Inflammation in Postmenopausal Women Stratified by Metabolic Syndrome Status. Metabolism 2015, 64, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, A.; Anzai, Y.; Tanji, N.; Imaizumi, H.; Fujita, M.; Hayashi, M.; Abe, K.; Ohira, H. Association of Equol with Obesity in Postmenopausal Women. Menopause 2021, 28, 807. [Google Scholar] [CrossRef]
- Lampe, J.W. Is Equol the Key to the Efficacy of Soy Foods? Am. J. Clin. Nutr. 2009, 89, 1664S–1667S. [Google Scholar] [CrossRef]
- Fujitani, T.; Fujii, Y.; Lyu, Z.; Harada Sassa, M.; Harada, K.H. Urinary Equol Levels Are Positively Associated with Urinary Estradiol Excretion in Women. Sci. Rep. 2021, 11, 19532. [Google Scholar] [CrossRef]
- Liu, B.; Qin, L.; Liu, A.; Uchiyama, S.; Ueno, T.; Li, X.; Wang, P. Prevalence of the Equol-Producer Phenotype and Its Relationship with Dietary Isoflavone and Serum Lipids in Healthy Chinese Adults. J. Epidemiol. 2010, 20, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Axelson, M.; Setchell, K.D.R. The Excretion of Lignans in Rats—Evidence for an Intestinal Bacterial Source for This New Group of Compounds. FEBS Lett. 1981, 123, 337–342. [Google Scholar] [CrossRef]
- Usui, T.; Tochiya, M.; Sasaki, Y.; Muranaka, K.; Yamakage, H.; Himeno, A.; Shimatsu, A.; Inaguma, A.; Ueno, T.; Uchiyama, S.; et al. Effects of Natural S -Equol Supplements on Overweight or Obesity and Metabolic Syndrome in the Japanese, Based on Sex and Equol Status. Clin. Endocrinol. 2013, 78, 365–372. [Google Scholar] [CrossRef]
- Yee, S.; Burdock, G.A.; Kurata, Y.; Enomoto, Y.; Narumi, K.; Hamada, S.; Itoh, T.; Shimomura, Y.; Ueno, T. Acute and Subchronic Toxicity and Genotoxicity of SE5-OH, an Equol-Rich Product Produced by Lactococcus Garvieae. Food Chem. Toxicol. 2008, 46, 2713–2720. [Google Scholar] [CrossRef]
- Tousen, Y.; Ezaki, J.; Fujii, Y.; Ueno, T.; Nishimuta, M.; Ishimi, Y. Natural S-Equol Decreases Bone Resorption in Postmenopausal, Non-Equol-Producing Japanese Women: A Pilot Randomized, Placebo-Controlled Trial. Menopause 2011, 18, 563–574. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Zhao, X.; Shoaf, S.E.; Ragland, K. The Pharmacokinetics of S-(-)Equol Administered as SE5-OH Tablets to Healthy Postmenopausal Women. J. Nutr. 2009, 139, 2037–2043. [Google Scholar] [CrossRef] [PubMed]
- Legette, L.L.; Prasain, J.; King, J.; Arabshahi, A.; Barnes, S.; Weaver, C.M. Pharmacokinetics of Equol, a Soy Isoflavone Metabolite, Changes with the Form of Equol (Dietary versus Intestinal Production) in Ovariectomized Rats. J. Agric. Food Chem. 2014, 62, 1294–1300. [Google Scholar] [CrossRef]
- Hazim, S.; Curtis, P.J.; Schär, M.Y.; Ostertag, L.M.; Kay, C.D.; Minihane, A.-M.; Cassidy, A. Acute Benefits of the Microbial-Derived Isoflavone Metabolite Equol on Arterial Stiffness in Men Prospectively Recruited According to Equol Producer Phenotype: A Double-Blind Randomized Controlled Trial. Am. J. Clin. Nutr. 2016, 103, 694–702. [Google Scholar] [CrossRef]
- Bax, E.N.; Cochran, K.E.; Mao, J.; Wiedmeyer, C.E.; Rosenfeld, C.S. Opposing Effects of S-Equol Supplementation on Metabolic and Behavioral Parameters in Mice Fed a High-Fat Diet. Nutr. Res. 2019, 64, 39–48. [Google Scholar] [CrossRef]
- Bowey, E.; Adlercreutz, H.; Rowland, I. Metabolism of Isoflavones and Lignans by the Gut Microflora: A Study in Germ-Free and Human Flora Associated Rats. Food Chem. Toxicol. 2003, 41, 631–636. [Google Scholar] [CrossRef] [PubMed]
- Franzosa, E.A.; Huang, K.; Meadow, J.F.; Gevers, D.; Lemon, K.P.; Bohannan, B.J.M.; Huttenhower, C. Identifying Personal Microbiomes Using Metagenomic Codes. Proc. Natl. Acad. Sci. USA 2015, 112, E2930–E2938. [Google Scholar] [CrossRef] [PubMed]
- Orcutt, R.P.; Gianni, F.J.; Judge, R.J. Development of an “Altered Schaedler Flora” for NCI Gnotobiotic Rodents. Microecol. Ther. 1987, 17, 59. [Google Scholar]
- Becker, N.; Kunath, J.; Loh, G.; Blaut, M. Human Intestinal Microbiota: Characterization of a Simplified and Stable Gnotobiotic Rat Model. Gut Microbes 2011, 2, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Brugiroux, S.; Beutler, M.; Pfann, C.; Garzetti, D.; Ruscheweyh, H.-J.; Ring, D.; Diehl, M.; Herp, S.; Lötscher, Y.; Hussain, S.; et al. Genome-Guided Design of a Defined Mouse Microbiota That Confers Colonization Resistance against Salmonella Enterica Serovar Typhimurium. Nat. Microbiol. 2017, 2, 16215. [Google Scholar] [CrossRef] [PubMed]
- Dewi, F.N.; Wood, C.E.; Lampe, J.W.; Hullar, M.A.J.; Franke, A.A.; Golden, D.L.; Adams, M.R.; Cline, J.M. Endogenous and Exogenous Equol Are Antiestrogenic in Reproductive Tissues of Apolipoprotein E-Null Mice. J. Nutr. 2012, 142, 1829–1835. [Google Scholar] [CrossRef]
- Matthies, A.; Loh, G.; Blaut, M.; Braune, A. Daidzein and Genistein Are Converted to Equol and 5-Hydroxy-Equol by Human Intestinal Slackia Isoflavoniconvertens in Gnotobiotic Rats1–23. J. Nutr. 2012, 142, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A Human Gut Microbial Gene Catalogue Established by Metagenomic Sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A Dietary Fiber-Deprived Gut Microbiota Degrades the Colonic Mucus Barrier and Enhances Pathogen Susceptibility. Cell 2016, 167, 1339–1353.e21. [Google Scholar] [CrossRef]
- Kasahara, K.; Krautkramer, K.A.; Org, E.; Romano, K.A.; Kerby, R.L.; Vivas, E.I.; Mehrabian, M.; Denu, J.M.; Bäckhed, F.; Lusis, A.J.; et al. Interactions between Roseburia Intestinalis and Diet Modulate Atherogenesis in a Murine Model. Nat. Microbiol. 2018, 3, 1461–1471. [Google Scholar] [CrossRef]
- Flórez, A.B.; Vázquez, L.; Rodríguez, J.; Redruello, B.; Mayo, B. Transcriptional Regulation of the Equol Biosynthesis Gene Cluster in Adlercreutzia Equolifaciens DSM19450T. Nutrients 2019, 11, 993. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S Ribosomal DNA Amplification for Phylogenetic Study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Krsek, M.; Baker, P.; Smalla, K.; Wellington, E.M. Analysis of Actinomycete Communities by Specific Amplification of Genes Encoding 16S rRNA and Gel-Electrophoretic Separation in Denaturing Gradients. Appl. Environ. Microbiol. 1997, 63, 3233–3241. [Google Scholar] [CrossRef]
- dos Santos, H.R.M.; Argolo, C.S.; Argôlo-Filho, R.C.; Loguercio, L.L. A 16S rDNA PCR-Based Theoretical to Actual Delta Approach on Culturable Mock Communities Revealed Severe Losses of Diversity Information. BMC Microbiol. 2019, 19, 74. [Google Scholar] [CrossRef] [PubMed]
- Decroos, K.; Vanhemmens, S.; Cattoir, S.; Boon, N.; Verstraete, W. Isolation and Characterisation of an Equol-Producing Mixed Microbial Culture from a Human Faecal Sample and Its Activity under Gastrointestinal Conditions. Arch. Microbiol. 2005, 183, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Minamida, K.; Tanaka, M.; Abe, A.; Sone, T.; Tomita, F.; Hara, H.; Asano, K. Production of Equol from Daidzein by Gram-Positive Rod-Shaped Bacterium Isolated from Rat Intestine. J. Biosci. Bioeng. 2006, 102, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Zhao, L.; Zhang, J.; Fang, X.; Zhong, Q.; Liao, Z.; Wang, J.; Guo, Y.; Liang, H.; Wang, L. Colonization Potential to Reconstitute a Microbe Community in Pseudo Germ-Free Mice After Fecal Microbe Transplant From Equol Producer. Front. Microbiol. 2020, 11, 1221. [Google Scholar] [CrossRef] [PubMed]
- Kano, M.; Takayanagi, T.; Harada, K.; Sawada, S.; Ishikawa, F. Bioavailability of Isoflavones after Ingestion of Soy Beverages in Healthy Adults1. J. Nutr. 2006, 136, 2291–2296. [Google Scholar] [CrossRef]
- Kemis, J.H.; Linke, V.; Barrett, K.L.; Boehm, F.J.; Traeger, L.L.; Keller, M.P.; Rabaglia, M.E.; Schueler, K.L.; Stapleton, D.S.; Gatti, D.M.; et al. Genetic Determinants of Gut Microbiota Composition and Bile Acid Profiles in Mice. PLoS Genet. 2019, 15, e1008073. [Google Scholar] [CrossRef] [PubMed]
- Kozich, J.J.; Westcott, S.L.; Baxter, N.T.; Highlander, S.K.; Schloss, P.D. Development of a Dual-Index Sequencing Strategy and Curation Pipeline for Analyzing Amplicon Sequence Data on the MiSeq Illumina Sequencing Platform. Appl. Environ. Microbiol. 2013, 79, 5112–5120. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Vázquez, L.; Guadamuro, L.; Giganto, F.; Mayo, B.; Flórez, A.B. Development and Use of a Real-Time Quantitative PCR Method for Detecting and Quantifying Equol-Producing Bacteria in Human Faecal Samples and Slurry Cultures. Front. Microbiol. 2017, 8, 1155. [Google Scholar] [CrossRef]
- Li, S.; Wang, T.; Fu, W.; Kennett, M.; Cox, A.D.; Lee, D.; Vanamala, J.K.P.; Reddivari, L. Role of Gut Microbiota in the Anti-Colitic Effects of Anthocyanin-Containing Potatoes. Mol. Nutr. Food Res. 2021, 65, 2100152. [Google Scholar] [CrossRef]
- Bisanz, J.E. qiime2R: Importing QIIME2 Artifacts and Associated Data into R Sessions. Version 099. 2018, Volume 13. Available online: https://github.com/jbisanz/qiime2R (accessed on 5 March 2024).
- Huttenhower, C.; Gevers, D.; Knight, R.; Abubucker, S.; Badger, J.H.; Chinwalla, A.T.; Creasy, H.H.; Earl, A.M.; FitzGerald, M.G.; Fulton, R.S.; et al. Structure, Function and Diversity of the Healthy Human Microbiome. Nature 2012, 486, 207–214. [Google Scholar] [CrossRef]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and Disease Markers Correlate with Gut Microbiome Composition across Thousands of People. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef]
- Vaiserman, A.; Romanenko, M.; Piven, L.; Moseiko, V.; Lushchak, O.; Kryzhanovska, N.; Guryanov, V.; Koliada, A. Differences in the Gut Firmicutes to Bacteroidetes Ratio across Age Groups in Healthy Ukrainian Population. BMC Microbiol. 2020, 20, 221. [Google Scholar] [CrossRef]
- Zafar, H.; Saier, M.H. Gut Bacteroides Species in Health and Disease. Gut Microbes 2021, 13, 1848158. [Google Scholar] [CrossRef]
- Lu, L.J.; Anderson, K.E. Sex and Long-Term Soy Diets Affect the Metabolism and Excretion of Soy Isoflavones in Humans. Am. J. Clin. Nutr. 1998, 68, 1500S–1504S. [Google Scholar] [CrossRef]
- Ohta, A.; Sakai, K.; Takasaki, M.; Uehara, M.; Adlercreutz, H.; Morohashi, T.; Ishimi, Y. A Combination of Dietary Fructooligosaccharides and Isoflavone Conjugates Increases Femoral Bone Mineral Density and Equol Production in Ovariectomized Mice. J. Nutr. 2002, 132, 2048–2054. [Google Scholar] [CrossRef]
- Tousen, Y.; Abe, F.; Ishida, T.; Uehara, M.; Ishimi, Y. Resistant Starch Promotes Equol Production and Inhibits Tibial Bone Loss in Ovariectomized Mice Treated with Daidzein. Metabolism 2011, 60, 1425–1432. [Google Scholar] [CrossRef]
- Tousen, Y.; Matsumoto, Y.; Matsumoto, C.; Nishide, Y.; Nagahata, Y.; Kobayashi, I.; Ishimi, Y. The Combined Effects of Soya Isoflavones and Resistant Starch on Equol Production and Trabecular Bone Loss in Ovariectomised Mice. Br. J. Nutr. 2016, 116, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Setchell, K.D.R.; Brown, N.M.; Lydeking-Olsen, E. The Clinical Importance of the Metabolite Equol-a Clue to the Effectiveness of Soy and Its Isoflavones. J. Nutr. 2002, 132, 3577–3584. [Google Scholar] [CrossRef] [PubMed]
- Leonard, L.M.; Choi, M.S.; Cross, T.-W.L. Maximizing the Estrogenic Potential of Soy Isoflavones through the Gut Microbiome: Implication for Cardiometabolic Health in Postmenopausal Women. Nutrients 2022, 14, 553. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-L.; Hur, H.-G.; Lee, J.H.; Kim, K.T.; Kim, S.-I. Enantioselective Synthesis of S -Equol from Dihydrodaidzein by a Newly Isolated Anaerobic Human Intestinal Bacterium. Appl. Environ. Microbiol. 2005, 71, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, S.; Ueno, T.; Suzuki, T. Identification of a Newly Isolated Equol-Producing Lactic Acid Bacterium from the Human Feces. J. Intest. Microbiol. 2007, 21, 217–220. [Google Scholar]
- Shimada, Y.; Yasuda, S.; Takahashi, M.; Hayashi, T.; Miyazawa, N.; Sato, I.; Abiru, Y.; Uchiyama, S.; Hishigaki, H. Cloning and Expression of a Novel NADP(H)-Dependent Daidzein Reductase, an Enzyme Involved in the Metabolism of Daidzein, from Equol-Producing Lactococcus Strain 20–92. Appl. Environ. Microbiol. 2010, 76, 5892–5901. [Google Scholar] [CrossRef] [PubMed]
- Elghali, S. Bioconversion of Daidzein to Equol by Bifidobacterium Breve 15700 and Bifidobacterium Longum BB536. J. Funct. Foods 2012, 10, 736–745. [Google Scholar] [CrossRef]
- Heng, Y.; Kim, M.J.; Yang, H.J.; Kang, S.; Park, S. Lactobacillus Intestinalis Efficiently Produces Equol from Daidzein and Chungkookjang, Short-Term Fermented Soybeans. Arch. Microbiol. 2019, 201, 1009–1017. [Google Scholar] [CrossRef] [PubMed]
- Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients 2019, 11, 433. [Google Scholar] [CrossRef]
- Sakamoto, M.; Sakurai, N.; Tanno, H.; Iino, T.; Ohkuma, M.; Endo, A. Genome-Based, Phenotypic and Chemotaxonomic Classification of Faecalibacterium Strains: Proposal of Three Novel Species Faecalibacterium duncaniae sp. nov., Faecalibacterium hattorii sp. nov. and Faecalibacterium gallinarum sp. nov. Int. J. Syst. Evol. Microbiol. 2022, 72. [Google Scholar] [CrossRef]
- Duncan, S.H. Growth Requirements and Fermentation Products of Fusobacterium Prausnitzii, and a Proposal to Reclassify It as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2002, 52, 2141–2146. [Google Scholar] [CrossRef] [PubMed]
- Rezzonico, E.; Mestdagh, R.; Delley, M.; Combremont, S.; Dumas, M.-E.; Holmes, E.; Nicholson, J.; Bibiloni, R. Bacterial Adaptation to the Gut Environment Favors Successful Colonization: Microbial and Metabonomic Characterization of a Simplified Microbiota Mouse Model. Gut Microbes 2011, 2, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Eberl, C.; Ring, D.; Münch, P.C.; Beutler, M.; Basic, M.; Slack, E.C.; Schwarzer, M.; Srutkova, D.; Lange, A.; Frick, J.S.; et al. Reproducible Colonization of Germ-Free Mice with the Oligo-Mouse-Microbiota in Different Animal Facilities. Front. Microbiol. 2020, 10, 2999. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum * | Species | Strain + | Bacterial Communities | |
|---|---|---|---|---|
| Equol (−) | Equol (+) | |||
| Bacteroidota | Bacteroides caccae | ATCC 43185 | + | + |
| Bacteroides thetaiotaomicron | DSM 2079 | + | + | |
| Bacteroides uniformis | ATCC 8492 | + | + | |
| Bacillota | Roseburia intestinalis | DSM 14610 | + | + |
| Faecalibacterium duncaniae | DSM 17677 | + | + | |
| Agathobacter rectalis | DSM 17629 | + | + | |
| Coprococcus comes | ATCC 27758 | + | + | |
| Verrucomicrobiota | Akkermansia muciniphila | DSM 22959 | + | + |
| Pseudomonadota | Providencia stuartii | ATCC 25826 | + | + |
| Actinomycetota | Collinsella aerofaciens | ATCC 25986 | + | + |
| Adlercreutzia equolifaciens | DSM 19450 | − | + | |
| Macronutrients | g% | kcal% |
|---|---|---|
| Protein | 19% | 20% |
| Carbohydrate | 67% | 70% |
| Fat | 4% | 10% |
| Ingredient | g/kg | kcal/kg |
| Casein- Mineral Acid | 190.18 | 761 |
| L-Cystine | 2.85 | 11 |
| Corn Starch | 476.59 | 1907 |
| Maltodextrin 10 | 118.86 | 475 |
| Sucrose | 60.67 | 242 |
| Inulin | 9.51 | 14.3 |
| short-chain fructo-oligosaccharides (scFOS) (93%) | 9.51 | 0.0 |
| Beta-Glucan (75.9%) | 9.51 | 5 |
| Pectin (80%) | 9.51 | 0 |
| Glucomannan NOW (98%) | 9.51 | 0 |
| Corn Oil | 23.77 | 214 |
| Lard | 19.02 | 171 |
| Mineral Mix S10026 | 9.51 | 0 |
| DiCalcium Phosphate | 12.36 | 0 |
| Calcium Carbonate | 5.23 | 0 |
| Potassium Citrate 1 H2O | 15.69 | 0 |
| Vitamin Mix V10001 | 14.26 | 57 |
| Choline Bitartrate | 1.90 | 0 |
| FD&C Red Dye #40 | 0.05 | 0 |
| Daidzein | 1.50 | 0 |
| Males | Females | ||||
|---|---|---|---|---|---|
| Equol (−) | Equol (+) | Equol (−) | Equol (+) | ||
| Final body weight (g) | 27.19 ± 0.635 | 28.02 ± 0.845 | 22.04 ± 0.445 | 22.40 ± 0.271 | |
| Mesenteric fat mass (g) | 0.28 ± 0.019 | 0.26 ± 0.030 | 0.18 ± 0.007 | 0.21 ± 0.016 | |
| Gonadal fat mass (g) | 0.43 ± 0.028 | 0.45 ± 0.044 | 0.26 ± 0.019 | 0.29 ± 0.028 | |
| Species | Strain | Daidzein (μM) | Equol (μM) |
|---|---|---|---|
| Bacteroides caccae | ATCC 43185 | 46.18 | 0 |
| Bacteroides thetaiotaomicron | DSM 2079 | 7.55 | 0 |
| Bacteroides uniformis | ATCC 8492 | 30.42 | 0 |
| Roseburia intestinalis | DSM 14610 | 5.07 | 0 |
| Faecalibacterium duncaniae | DSM 17677 | 44.43 | 0 |
| Agathobacter rectalis | DSM 17629 | 13.35 | 0 |
| Coprococcus comes | ATCC 27758 | 36.14 | 0 |
| Akkermansia muciniphila | DSM 22959 | 28.04 | 0 |
| Providencia stuartii | ATCC 25826 | 40.21 | 0 |
| Collinsella aerofaciens | ATCC 25986 | 34.81 | 0 |
| Adlercreutzia equolifaciens | DSM 19450 | 3.12 | 29.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leonard, L.M.; Simpson, A.M.R.; Li, S.; Reddivari, L.; Cross, T.-W.L. A Gnotobiotic Mouse Model with Divergent Equol-Producing Phenotypes: Potential for Determining Microbial-Driven Health Impacts of Soy Isoflavone Daidzein. Nutrients 2024, 16, 1079. https://doi.org/10.3390/nu16071079
Leonard LM, Simpson AMR, Li S, Reddivari L, Cross T-WL. A Gnotobiotic Mouse Model with Divergent Equol-Producing Phenotypes: Potential for Determining Microbial-Driven Health Impacts of Soy Isoflavone Daidzein. Nutrients. 2024; 16(7):1079. https://doi.org/10.3390/nu16071079
Chicago/Turabian StyleLeonard, Lindsay M., Abigayle M. R. Simpson, Shiyu Li, Lavanya Reddivari, and Tzu-Wen L. Cross. 2024. "A Gnotobiotic Mouse Model with Divergent Equol-Producing Phenotypes: Potential for Determining Microbial-Driven Health Impacts of Soy Isoflavone Daidzein" Nutrients 16, no. 7: 1079. https://doi.org/10.3390/nu16071079
APA StyleLeonard, L. M., Simpson, A. M. R., Li, S., Reddivari, L., & Cross, T.-W. L. (2024). A Gnotobiotic Mouse Model with Divergent Equol-Producing Phenotypes: Potential for Determining Microbial-Driven Health Impacts of Soy Isoflavone Daidzein. Nutrients, 16(7), 1079. https://doi.org/10.3390/nu16071079