
Lactobacillus helveticus-Derived Whey-Calcium Chelate Promotes Calcium Absorption and Bone Health of Rats Fed a Low-Calcium Diet
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Whey-Calcium Chelate
2.3. Fourier-Transform Infrared Spectroscopy (FT-IR)
2.4. In Vitro Digestion
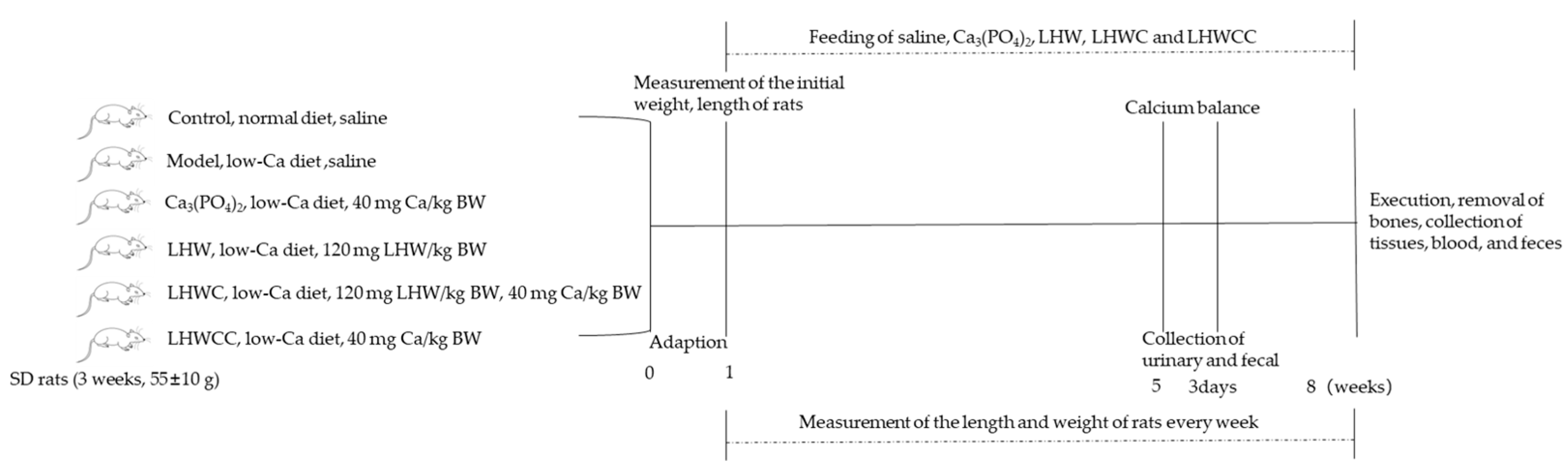
2.5. Animal Experiments
2.6. Analysis of Serum Biochemical Indexes
2.7. Analysis of Femur Length, Diameter, and Weight
2.8. Analysis of Calcium Content in Femur
2.9. BMD and Bone Mass Measurements
2.10. Bone Biomechanical Strength Measurements
2.11. Analysis of Calcium-Apparent Absorption and Retention Rate
2.12. RNA Extraction and Real-Time RT-PCR
2.13. Statistical Analysis
3. Results
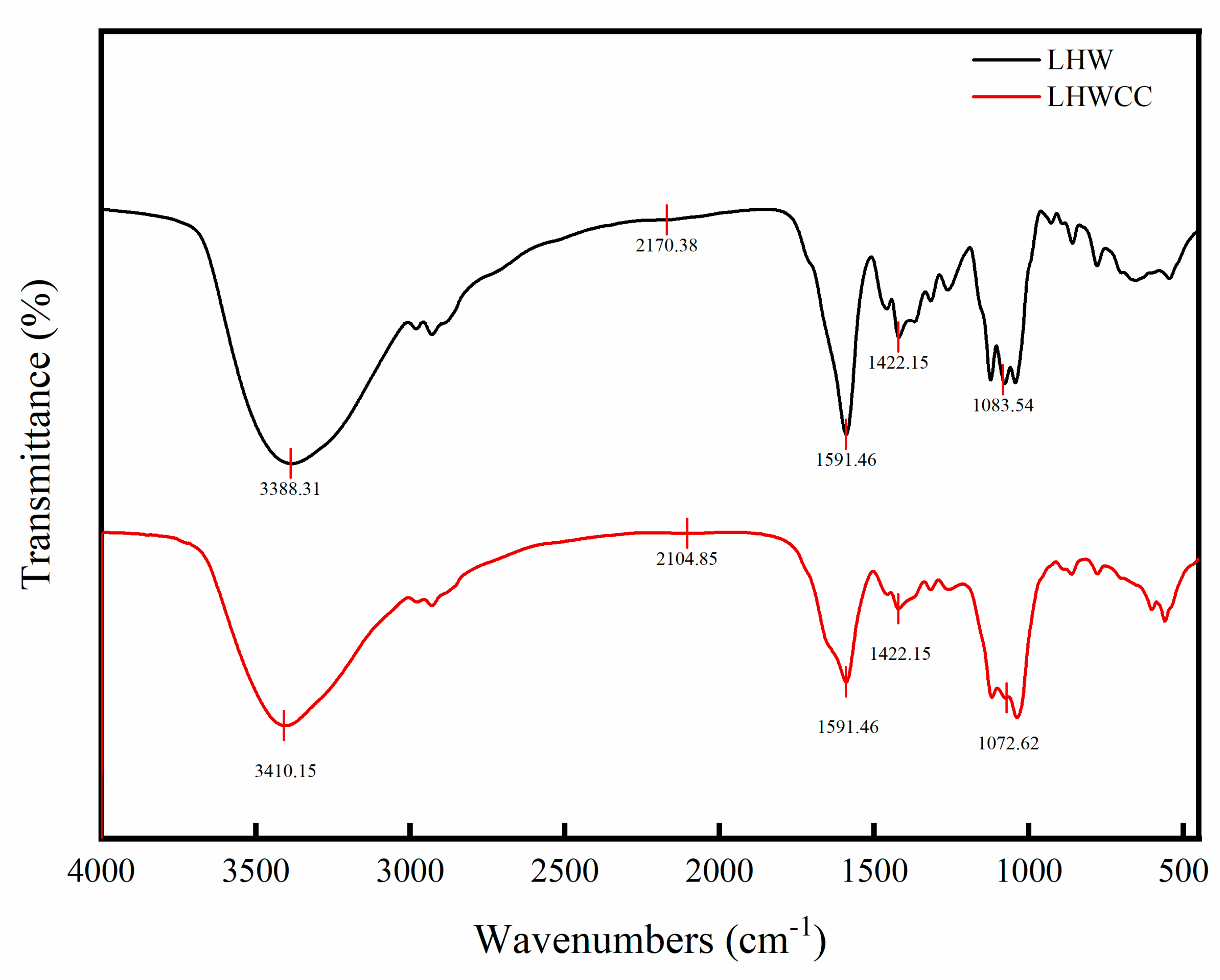
3.1. Fourier Transform-Infrared Spectroscopy of LHWCC
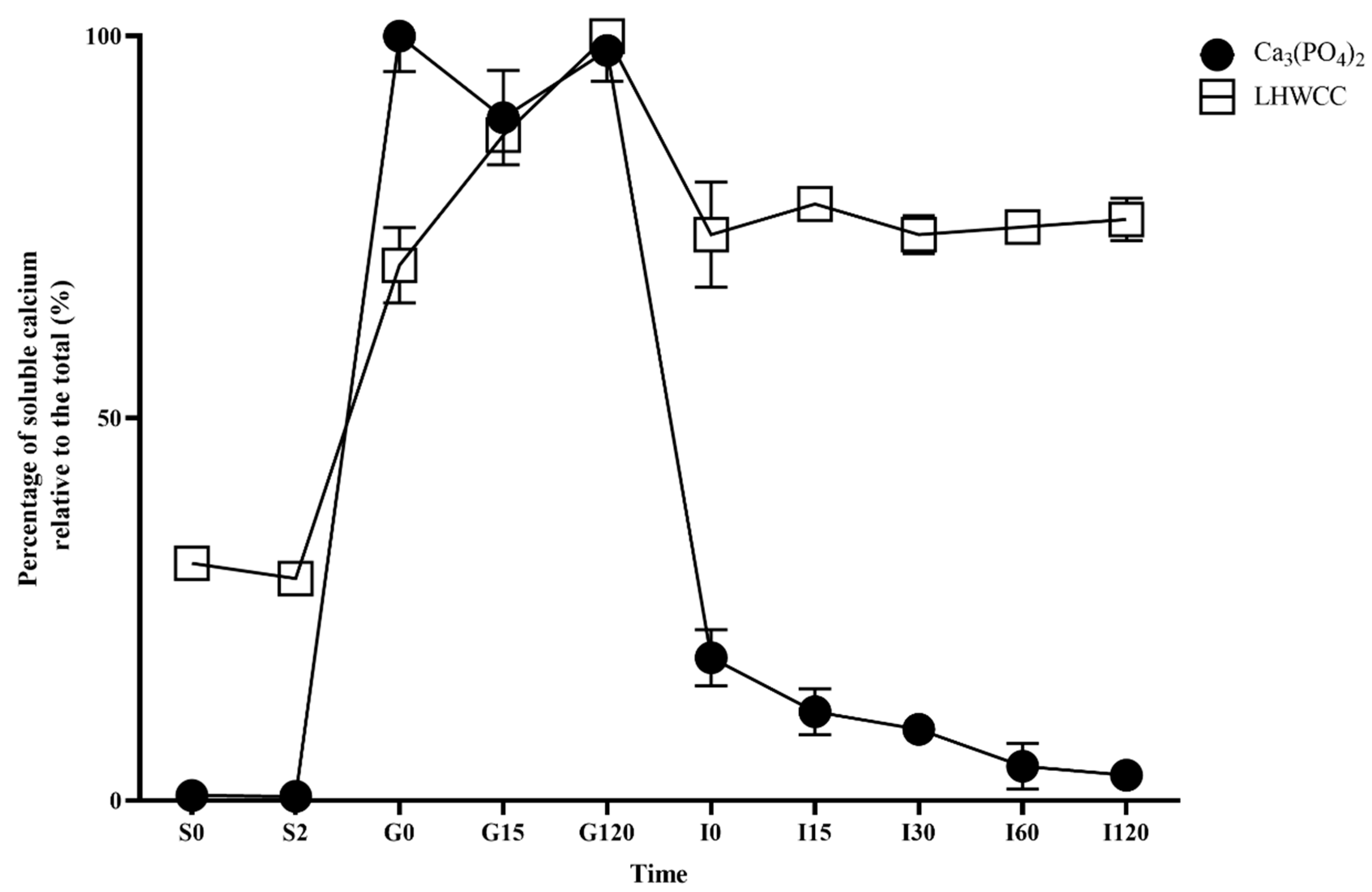
3.2. Soluble Calcium Content of LHWCC In Vitro Digestion
3.3. Effect of LHWCC on Weight, Body Length, and Tail Length Gain
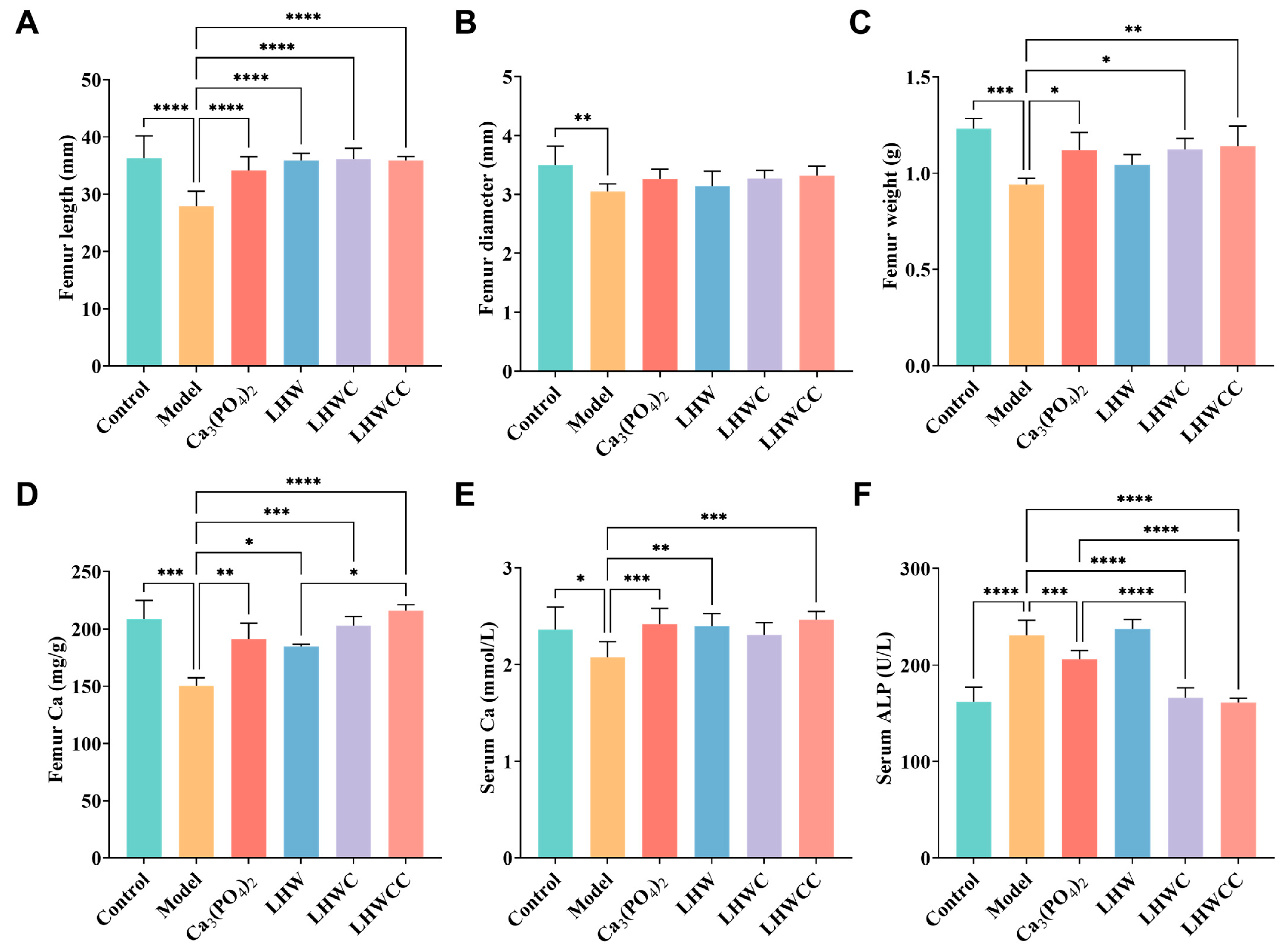
3.4. Effect of LHWCC on Femoral and Serum Indicators
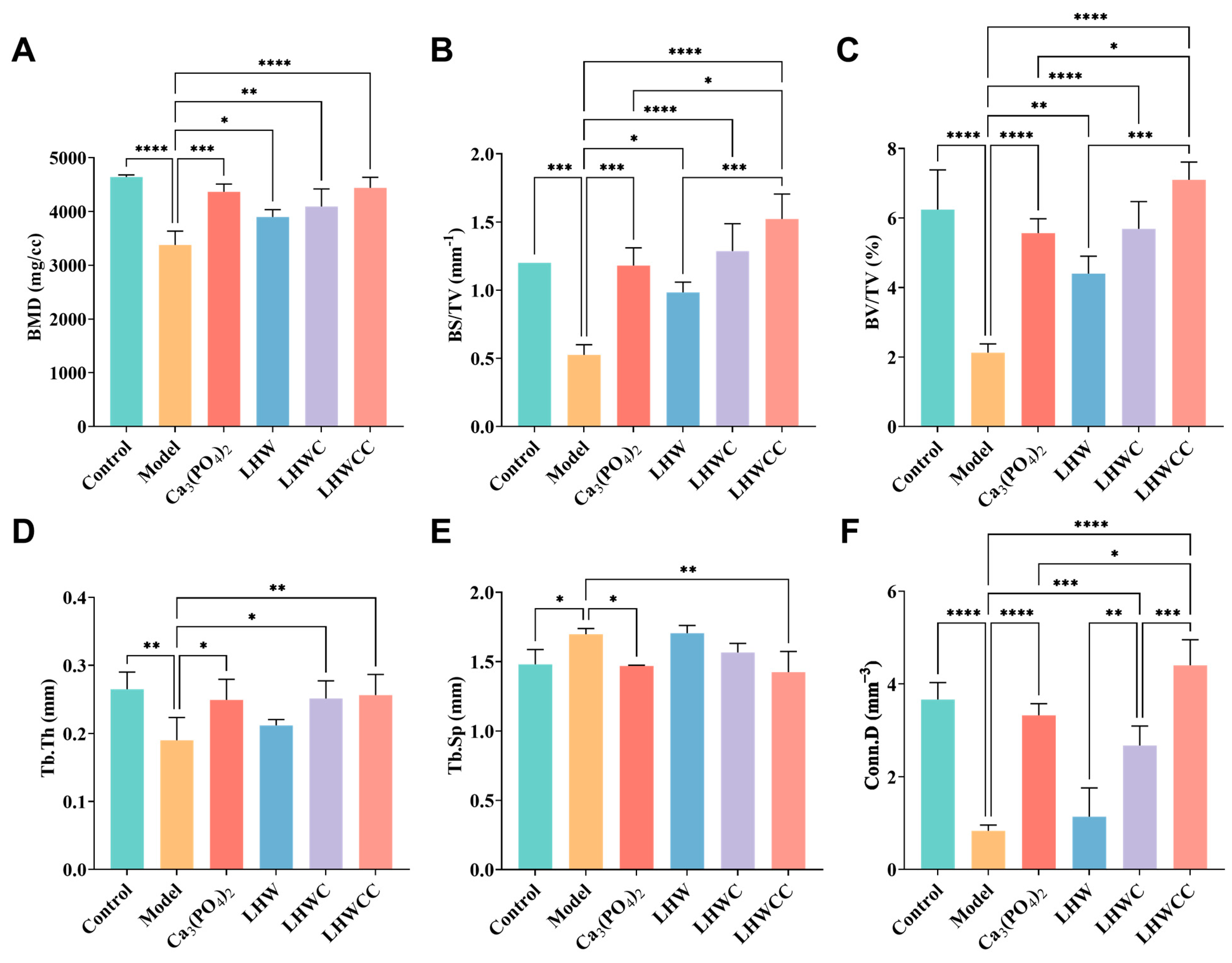
3.5. Bone Biomechanical Parameters and Histomorphometry
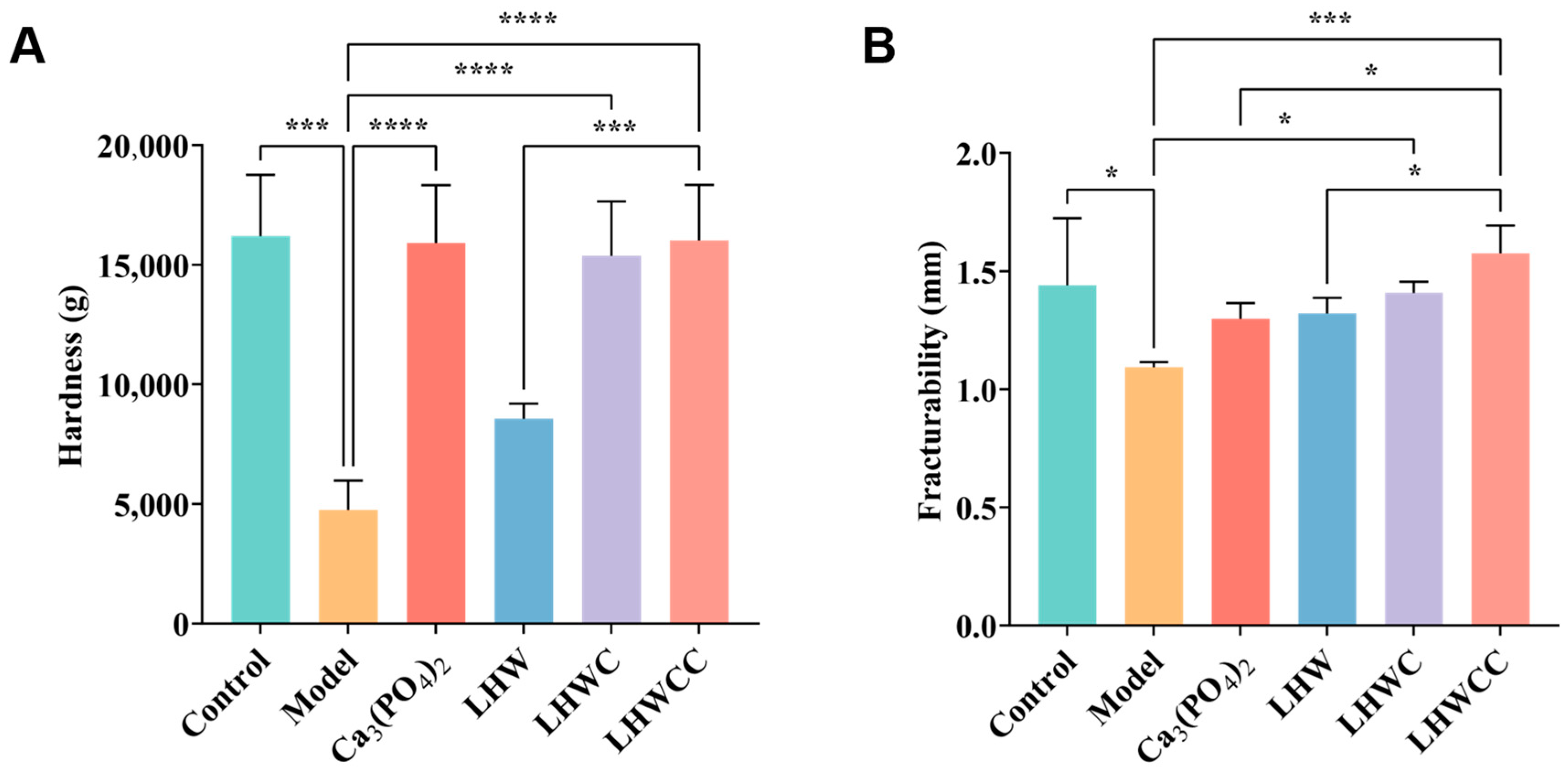
3.6. Effects on Bone Biomechanical Strength
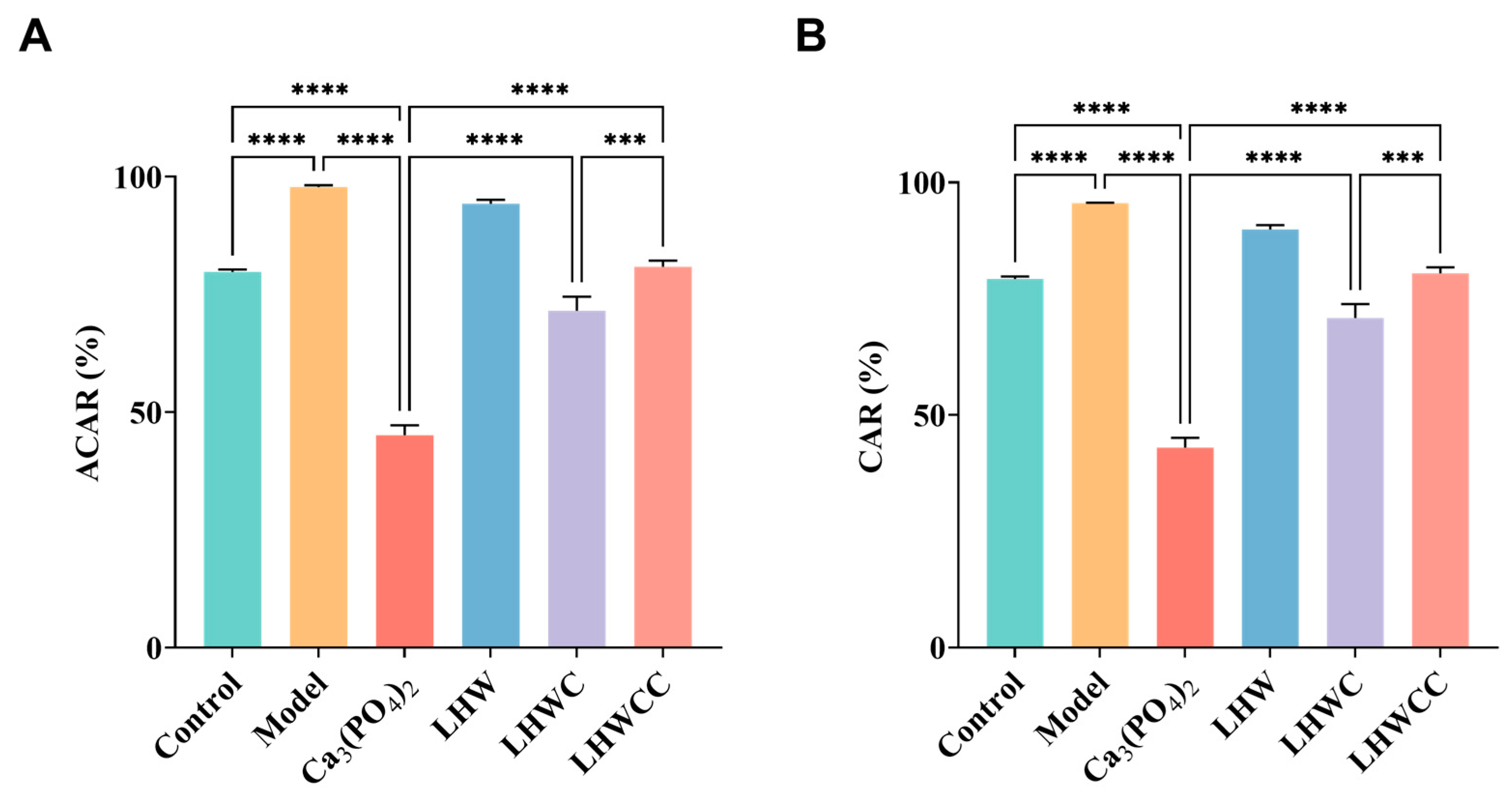
3.7. Effect of LHWCC on Calcium Balance
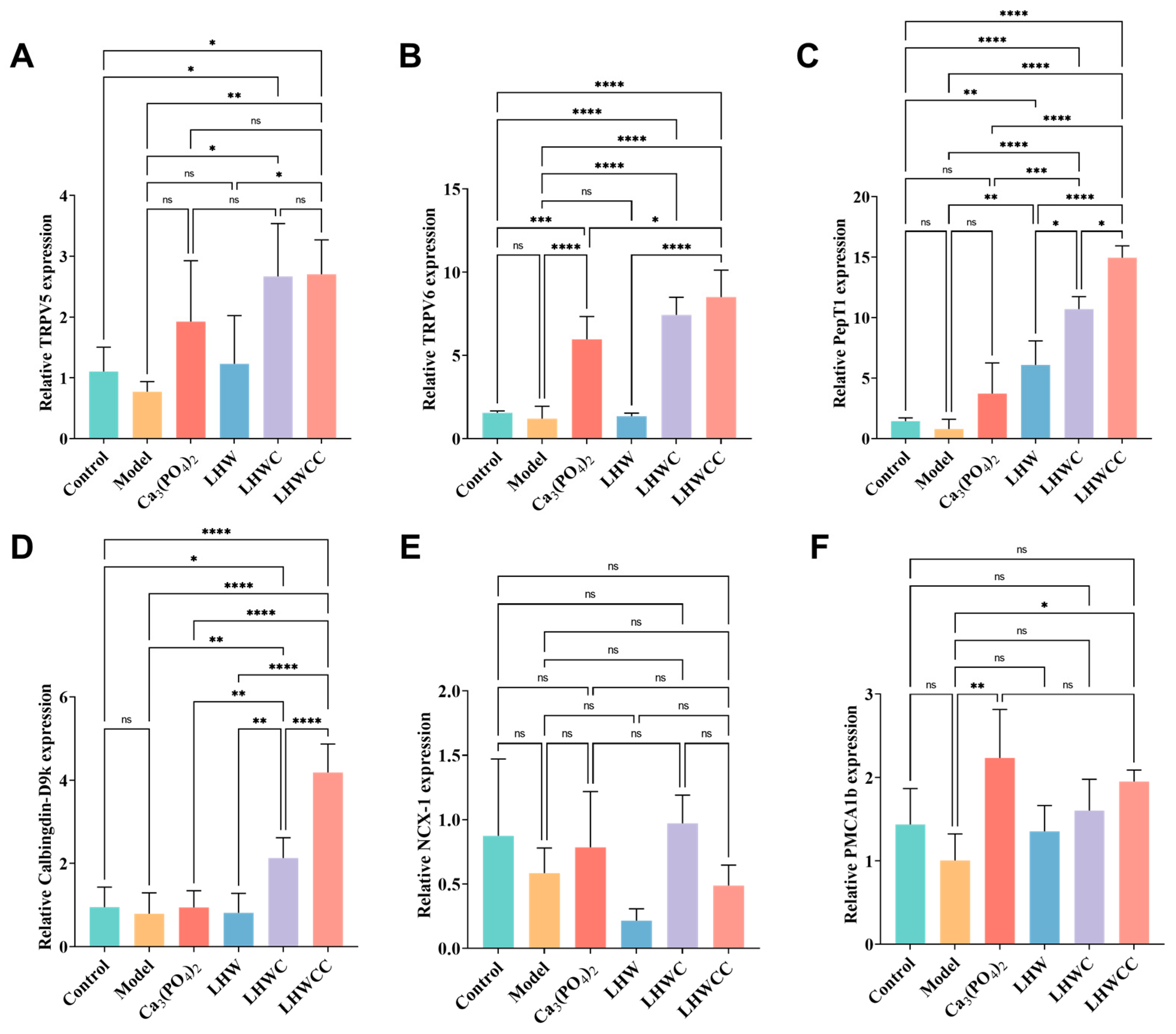
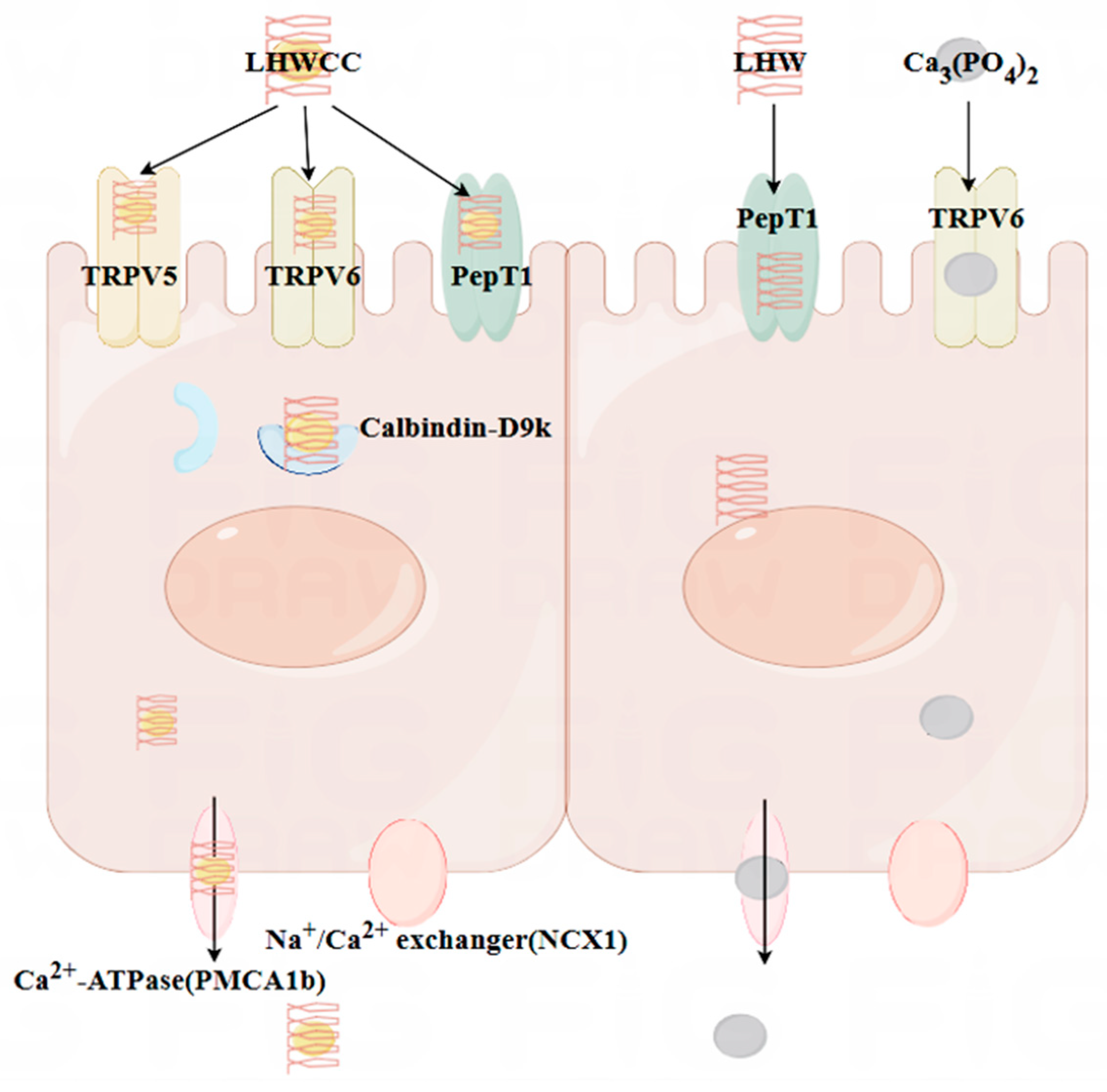
3.8. Gene Expression of Corresponding Receptors in the Intestines of Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flynn, A. The Role of Dietary Calcium in Bone Health. Proc. Nutr. Soc. 2003, 62, 851–858. [Google Scholar] [CrossRef]
- Sun, N.; Wu, H.; Du, M.; Tang, Y.; Liu, H.; Fu, Y.; Zhu, B. Food Protein-Derived Calcium Chelating Peptides: A Review. Trends Food Sci. Technol. 2016, 58, 140–148. [Google Scholar] [CrossRef]
- Welch, A.A.; Fransen, H.; Jenab, M.; Boutron-Ruault, M.C.; Tumino, R.; Agnoli, C.; Ericson, U.; Johansson, I.; Ferrari, P.; Engeset, D.; et al. Variation in Intakes of Calcium, Phosphorus, Magnesium, Iron and Potassium in 10 Countries in the European Prospective Investigation into Cancer and Nutrition Study. Eur. J. Clin. Nutr. 2009, 63, S101–S121. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Zhang, Y.; Ying, Z.; Li, H.; Liu, W.; Wang, J.; Liu, X. The Formation, Structural Characteristics, Absorption Pathways and Bioavailability of Calcium–Peptide Chelates. Foods 2022, 11, 2762. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.; Liu, H.; Ren, J.; Li, X.; Guo, S. The Positive Effect of Soybean Protein Hydrolysates—Calcium Complexes on Bone Mass of Rapidly Growing Rats. Food Funct. 2013, 4, 1245. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Gao, A.; Chen, Y.; Zhang, X.; Li, S.; Chen, Y. Preparation of Cucumber Seed Peptide-Calcium Chelate by Liquid State Fermentation and Its Characterization. Food Chem. 2017, 229, 487–494. [Google Scholar] [CrossRef]
- Li, H.; Duan, S.; Yuan, P.; Liu, J.; Wang, X.; Liu, Y.; Peng, Y.; Pan, C.; Xia, K. Preparation of Casein Phosphopeptides Calcium Complex and the Promotion in Calcium Cellular Uptake through Transcellular Transport Pathway. J. Food Biochem. 2021, 45, e14001. [Google Scholar] [CrossRef]
- Tian, L.; Xiong, D.; Jia, J.; Liu, X.; Zhang, Y.; Duan, X. Mechanism Study on Enhanced Emulsifying Properties of Phosvitin and Calcium-Binding Capacity of Its Phosphopeptides by Lactic Acid Bacteria Fermentation. LWT 2022, 155, 113002. [Google Scholar] [CrossRef]
- Lorieau, L.; Le Roux, L.; Gaucheron, F.; Ligneul, A.; Hazart, E.; Dupont, D.; Floury, J. Bioaccessibility of Four Calcium Sources in Different Whey-Based Dairy Matrices Assessed by in Vitro Digestion. Food Chem. 2018, 245, 454–462. [Google Scholar] [CrossRef]
- Hua, P.; Xiong, Y.; Yu, Z.; Liu, B.; Zhao, L. Effect of Chlorella Pyrenoidosa Protein Hydrolysate-Calcium Chelate on Calcium Absorption Metabolism and Gut Microbiota Composition in Low-Calcium Diet-Fed Rats. Mar. Drugs 2019, 17, 348. [Google Scholar] [CrossRef]
- Giraffa, G.; Gatti, M.; Rossetti, L.; Senini, L.; Neviani, E. Molecular Diversity within Lactobacillus Helveticus as Revealed by Genotypic Characterization. Appl. Environ. Microbiol. 2000, 66, 1259–1265. [Google Scholar] [CrossRef] [PubMed]
- Sipola, M.; Finckenberg, P.; Santisteban, J.; Korpela, R.; Vapaatalo, H.; Nurminen, M.L. Long-Term Intake of Milk Peptides Attenuates Development of Hypertension in Spontaneously Hypertensive Rats. J. Physiol. Pharmacol. 2001, 52, 745–754. [Google Scholar]
- Jauhiainen, T.; Vapaatalo, H.; Poussa, T.; Kyrönpalo, S.; Rasmussen, M.; Korpela, R. Lactobacillus Helveticus Fermented Milk Lowers Blood Pressure in Hypertensive Subjects in 24-h Ambulatory Blood Pressure Measurement. Am. J. Hypertens 2005, 18, 1600–1605. [Google Scholar] [CrossRef]
- Ohsawa, K.; Uchida, N.; Ohki, K.; Nakamura, Y.; Yokogoshi, H. Lactobacillus Helveticus-Fermented Milk Improves Learning and Memory in Mice. Nutr. Neurosci. 2015, 18, 232–240. [Google Scholar] [PubMed]
- Gao, S.; Jiang, Y.; Zhang, X.; Cui, S.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Comparative Peptidomics Analysis of Milk Fermented by Lactobacillus Helveticus. Foods 2022, 11, 3885. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Jiang, Y.; Yu, P.; Lee, Y.K.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Effect of Carbon Catabolite Repression on Lactose and Galactose Catabolism in Lacticaseibacillus Paracasei. Food Biosci. 2021, 40, 100912. [Google Scholar] [CrossRef]
- Lao, L.; He, J.; Liao, W.; Zeng, C.; Liu, G.; Cao, Y.; Miao, J. Casein Calcium-Binding Peptides: Preparation, Characterization, and Promotion of Calcium Uptake in Caco-2 Cell Monolayers. Process Biochem. 2023, 130, 78–86. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carrière, F.; Boutrou, R.; Corredig, M.; Dupont, D.; et al. A Standardised Static In Vitro Digestion Method Suitable for Food—An International Consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Chen, J.; Qiu, X.; Hao, G.; Zhang, M.; Weng, W. Preparation and Bioavailability of Calcium-chelating Peptide Complex from Tilapia Skin Hydrolysates. J. Sci. Food Agric. 2017, 97, 4898–4903. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Liu, D.; Tang, X.; Cui, Y.; Hu, W.; Regenstein, J.M.; Zhou, P. Sea Cucumber Enzymatic Hydrolysates Relieve Osteoporosis through OPG/RANK/RANKL System in Ovariectomized Rats. Food Biosci. 2022, 46, 101572. [Google Scholar] [CrossRef]
- Ye, M.; Zhang, C.; Jia, W.; Shen, Q.; Qin, X.; Zhang, H.; Zhu, L. Metabolomics Strategy Reveals the Osteogenic Mechanism of Yak (Bos Grunniens) Bone Collagen Peptides on Ovariectomy-Induced Osteoporosis in Rats. Food Funct. 2020, 11, 1498–1512. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, K.; Feng, Y.; Jiang, S.; Zhao, Y.; Zeng, M. Preparation, Characterization of L-Aspartic Acid Chelated Calcium from Oyster Shell Source and Its Calcium Supplementation Effect in Rats. J. Funct. Foods 2020, 75, 104249. [Google Scholar] [CrossRef]
- Sun, L.; Liu, J.; Pei, H.; Shi, M.; Chen, W.; Zong, Y.; Zhao, Y.; Li, J.; Du, R.; He, Z. Structural Characterisation of Deer Sinew Peptides as Calcium Carriers, Their Promotion of MC3T3-E1 Cell Proliferation and Their Effect on Bone Deposition in Mice. Food Funct. 2024, 15, 2587–2603. [Google Scholar] [CrossRef] [PubMed]
- Zhai, W.; Lin, D.; Mo, R.; Zou, X.; Zhang, Y.; Zhang, L.; Ge, Y. Process Optimization, Structural Characterization, and Calcium Release Rate Evaluation of Mung Bean Peptides-Calcium Chelate. Foods 2023, 12, 1058. [Google Scholar] [CrossRef]
- Zhang, X.; Jia, Q.; Li, M.; Liu, H.; Wang, Q.; Wu, Y.; Niu, L.; Liu, Z. Isolation of a Novel Calcium-Binding Peptide from Phosvitin Hydrolysates and the Study of Its Calcium Chelation Mechanism. Food Res. Int. 2021, 141, 110169. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, J.; Zhao, H.; Zhao, R.; Xu, Y.; Lyu, X. Preparation of Grape Seed Polypeptide and Its Calcium Chelate with Determination of Calcium Bioaccessibility and Structural Characterisation. Int. J. Food Sci. Tech. 2021, 56, 166–177. [Google Scholar] [CrossRef]
- Dyck, K.V.; Tas, S.; Robberecht, H.; Deelstra, H. The Influence of Different Food Components on the in Vitro Availability of Iron, Zinc and Calcium from a Composed Meal. Int. J. Food Sci. Nutr. 1996, 47, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Bronner, F.; Pansu, D. Nutritional Aspects of Calcium Absorption1. J. Nutr. 1999, 129, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Ménard, O.; Bourlieu, C.; De Oliveira, S.C.; Dellarosa, N.; Laghi, L.; Carrière, F.; Capozzi, F.; Dupont, D.; Deglaire, A. A First Step towards a Consensus Static in Vitro Model for Simulating Full-Term Infant Digestion. Food Chem. 2018, 240, 338–345. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, R.; Bai, H.; Wang, S.; Liu, T.; Zhang, X.; Wang, Z. Casein Phosphopeptide Calcium Chelation: Preparation Optimization, in Vitro Gastrointestinal Simulated Digestion, and Peptide Fragment Exploration. J. Sci. Food Agric. 2024, 104, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Xia, A.; Jiang, Y.; Lee, Y.-K.; Liang, D.; Yang, B.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Identification and Validation of Fermented Milk-Derived Osteogenic Peptides via Molecular Docking and Osteoblastic Cell Model. Food Biosci. 2024, 58, 103698. [Google Scholar] [CrossRef]
- Peng, Z.; Hou, H.; Zhang, K.; Li, B. Effect of Calcium-Binding Peptide from Pacific Cod (Gadus Macrocephalus) Bone on Calcium Bioavailability in Rats. Food Chem. 2017, 221, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R.; Heaney, R.P. Co-Dependence of Calcium and Phosphorus for Growth and Bone Development under Conditions of Varying Deficiency. Bone 2003, 32, 532–540. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Szeto, I.M.-Y.; Li, N.; Yang, H.; Zhou, Y.; Liu, B.; He, F.; Zhang, L.; Duan, S.; Chen, J. Effects of Menaquinone-7 on the Bone Health of Growing Rats under Calcium Restriction: New Insights from Microbiome-Metabolomics. Nutrients 2023, 15, 3398. [Google Scholar] [CrossRef]
- Benzvi, L.; Gershon, A.; Lavi, I.; Wollstein, R. Secondary Prevention of Osteoporosis Following Fragility Fractures of the Distal Radius in a Large Health Maintenance Organization. Arch. Osteoporos 2016, 11, 20. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, L.-L.; Ding, S.-Q.; Liu, J.-J.; Dong, Y.-H.; Li, Y.-T.; Li, N.; Zhao, X.-J.; Hu, C.-L.; Jiang, Y.; et al. Anti-Osteoporotic Activity of an Edible Traditional Chinese Medicine Cistanche Deserticola on Bone Metabolism of Ovariectomized Rats Through RANKL/RANK/TRAF6-Mediated Signaling Pathways. Front. Pharmacol. 2019, 10, 1412. [Google Scholar] [CrossRef]
- Zhao, M.; Ahn, D.U.; Li, S.; Liu, W.; Yi, S.; Huang, X. Effects of Phosvitin Phosphopeptide-Ca Complex Prepared by Efficient Enzymatic Hydrolysis on Calcium Absorption and Bone Deposition of Mice. Food Sci. Hum. Wellness 2022, 11, 1631–1640. [Google Scholar] [CrossRef]
- Keaveny, T.M.; Morgan, E.F.; Niebur, G.L.; Yeh, O.C. Biomechanics of Trabecular Bone. Annu. Rev. Biomed. Eng. 2001, 3, 307–333. [Google Scholar] [CrossRef]
- Chen, D.; Mu, X.; Huang, H.; Nie, R.; Liu, Z.; Zeng, M. Isolation of a Calcium-Binding Peptide from Tilapia Scale Protein Hydrolysate and Its Calcium Bioavailability in Rats. J. Funct. Foods 2014, 6, 575–584. [Google Scholar] [CrossRef]
- Zhao, N.; Hu, J.; Hou, T.; Ma, Z.; Wang, C.; He, H. Effects of Desalted Duck Egg White Peptides and Their Products on Calcium Absorption in Rats. J. Funct. Foods 2014, 8, 234–242. [Google Scholar] [CrossRef]
- Liu, W.-Y.; Lu, J.; Gao, F.; Gu, R.-Z.; Lin, F.; Ren, D.-F.; Cai, M.-Y. Preparation, Characterization and Identification of Calcium-Chelating Atlantic Salmon (Salmo Salar L.) Ossein Oligopeptides. Eur. Food Res. Technol. 2015, 241, 851–860. [Google Scholar] [CrossRef]
- Nijenhuis, T.; Hoenderop, J.G.J.; van der Kemp, A.W.C.M.; Bindels, R.J.M. Localization and Regulation of the Epithelial Ca2+ Channel TRPV6 in the Kidney. J. Am. Soc. Nephrol. 2003, 14, 2731–2740. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, X.-L.; Leung, P.-C.; Che, C.-T.; Wong, M.-S. Fructus Ligustri Lucidi Extract Improves Calcium Balance and Modulates the Calciotropic Hormone Level and Vitamin D-Dependent Gene Expression in Aged Ovariectomized Rats. Menopause 2008, 15, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef]
- Goda, T.; Kishi, K.; Ezawa, I.; Takase, S. The Maltitol-Induced Increase in Intestinal Calcium Transport Increases the Calcium Content and Breaking Force of Femoral Bone in Weanling Rats. J. Nutr. 1998, 128, 2028–2031. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence |
|---|---|
| TRPV6 | forward 5′-CACCCAGTGGACGTATGGAC-3′ |
| reverse 5′-CTCGTGCGGTTATTGGTCCT-3′ | |
| TRPV5 | forward 5′-ACGTATGGACCCCTGACCTC-3′ |
| reverse 5′-GAATTTGGCGAGCCTCTCGT-3′ | |
| Calbindin-D9K | forward 5′-GGCAACCAGACACCAGAATGA-3′ |
| reverse 5′-TGACAACTGGTCTGGATCACC-3′ | |
| NCX-1 | forward 5′-TTGAGATTGGAGAACCCCGT-3′ |
| reverse 5′-ATGTGAAGCCACCAAGCTCA-3′ | |
| PMCA1b | forward 5′-AGTGATTGTTGCTTTTACGGGC-3′ |
| reverse 5′-AGAGACTCAGTGGGTGGTTCCG-3′ | |
| PepT1 | forward 5′-ATCTACCATACGTTTGTTGC-3′ |
| reverse 5′-CTGGGGCTGAAACTTCTT-3′ | |
| β-Actin | forward 5′-CACCCAGCACAATGAAGATCAAGAT-3′ |
| reverse 5′-CCAGTTTTTAAATCCTGAGTCAAGC-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, W.; Pei, Z.; Xia, A.; Jiang, Y.; Yang, B.; Liu, X.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus helveticus-Derived Whey-Calcium Chelate Promotes Calcium Absorption and Bone Health of Rats Fed a Low-Calcium Diet. Nutrients 2024, 16, 1127. https://doi.org/10.3390/nu16081127
Hu W, Pei Z, Xia A, Jiang Y, Yang B, Liu X, Zhao J, Zhang H, Chen W. Lactobacillus helveticus-Derived Whey-Calcium Chelate Promotes Calcium Absorption and Bone Health of Rats Fed a Low-Calcium Diet. Nutrients. 2024; 16(8):1127. https://doi.org/10.3390/nu16081127
Chicago/Turabian StyleHu, Wei, Zhiwen Pei, Aonan Xia, Yang Jiang, Bo Yang, Xiaoming Liu, Jianxin Zhao, Hao Zhang, and Wei Chen. 2024. "Lactobacillus helveticus-Derived Whey-Calcium Chelate Promotes Calcium Absorption and Bone Health of Rats Fed a Low-Calcium Diet" Nutrients 16, no. 8: 1127. https://doi.org/10.3390/nu16081127