
Diet-Induced Early Inflammatory Response of Visceral Adipose Tissue in Healthy Male Wistar Rats

 , ,
, ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
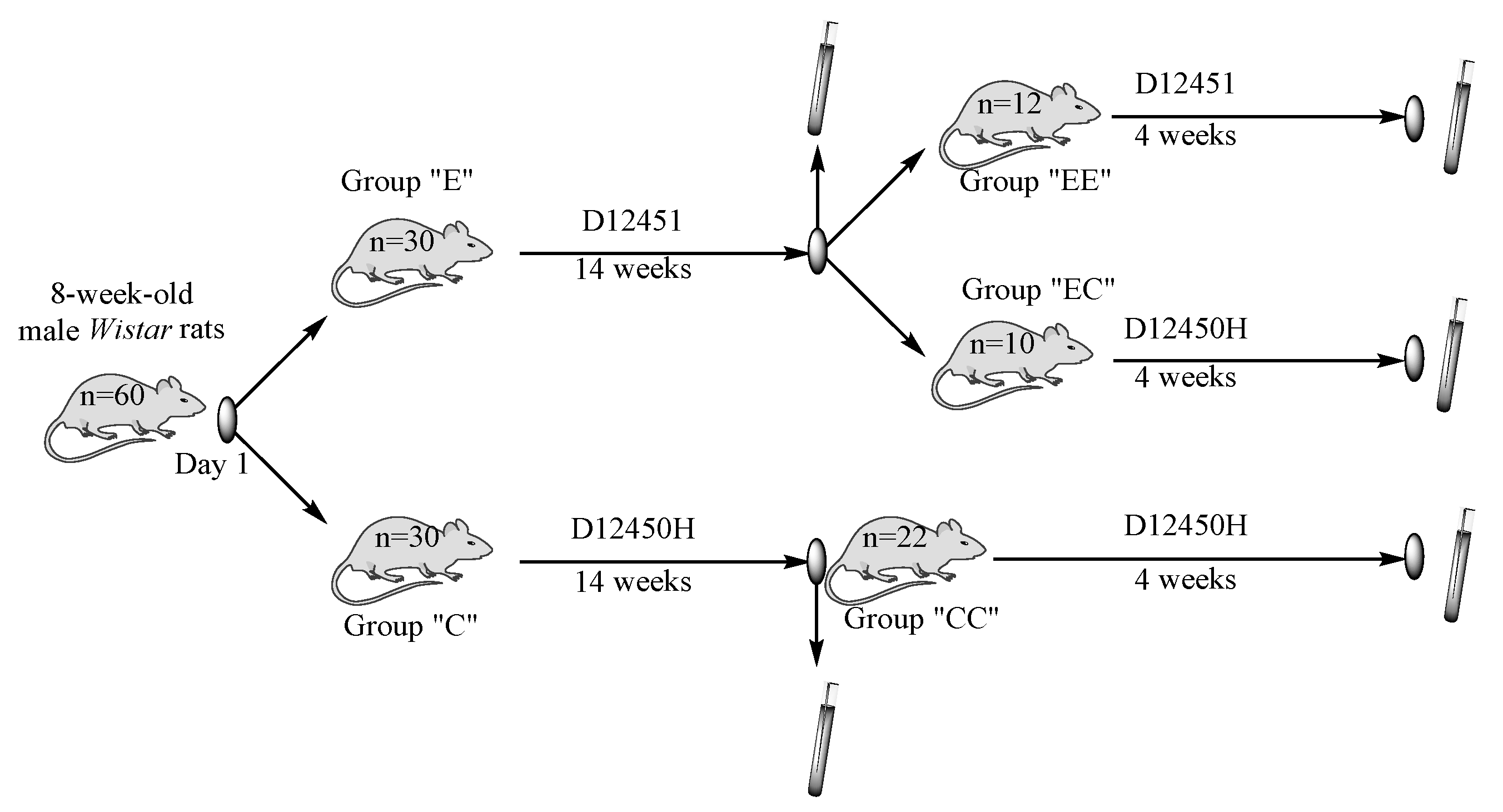
2.1. Animal Care
2.2. Study Design and Sample Collection
2.3. Statistical Analysis
3. Results
3.1. Body Weight, Body Mass Index (BMI), Relative Fat Mass (RFM), Conicity Index (CI), and Lee Index
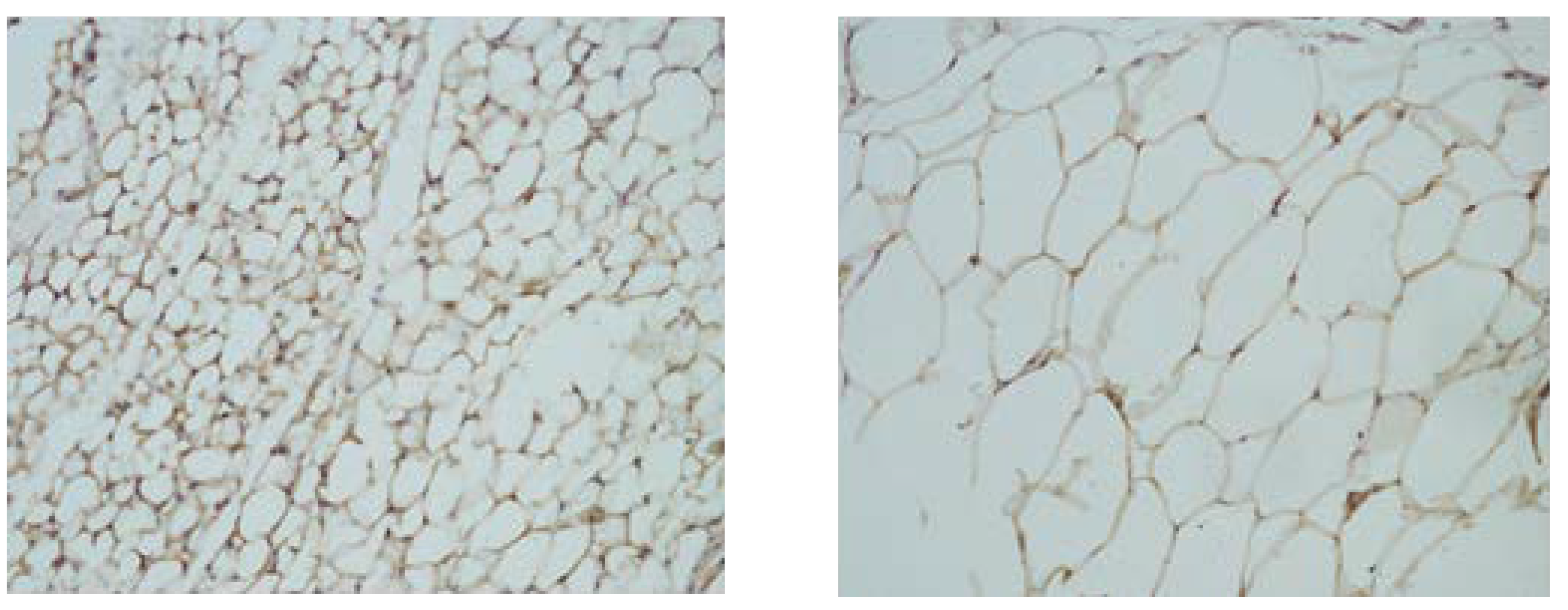
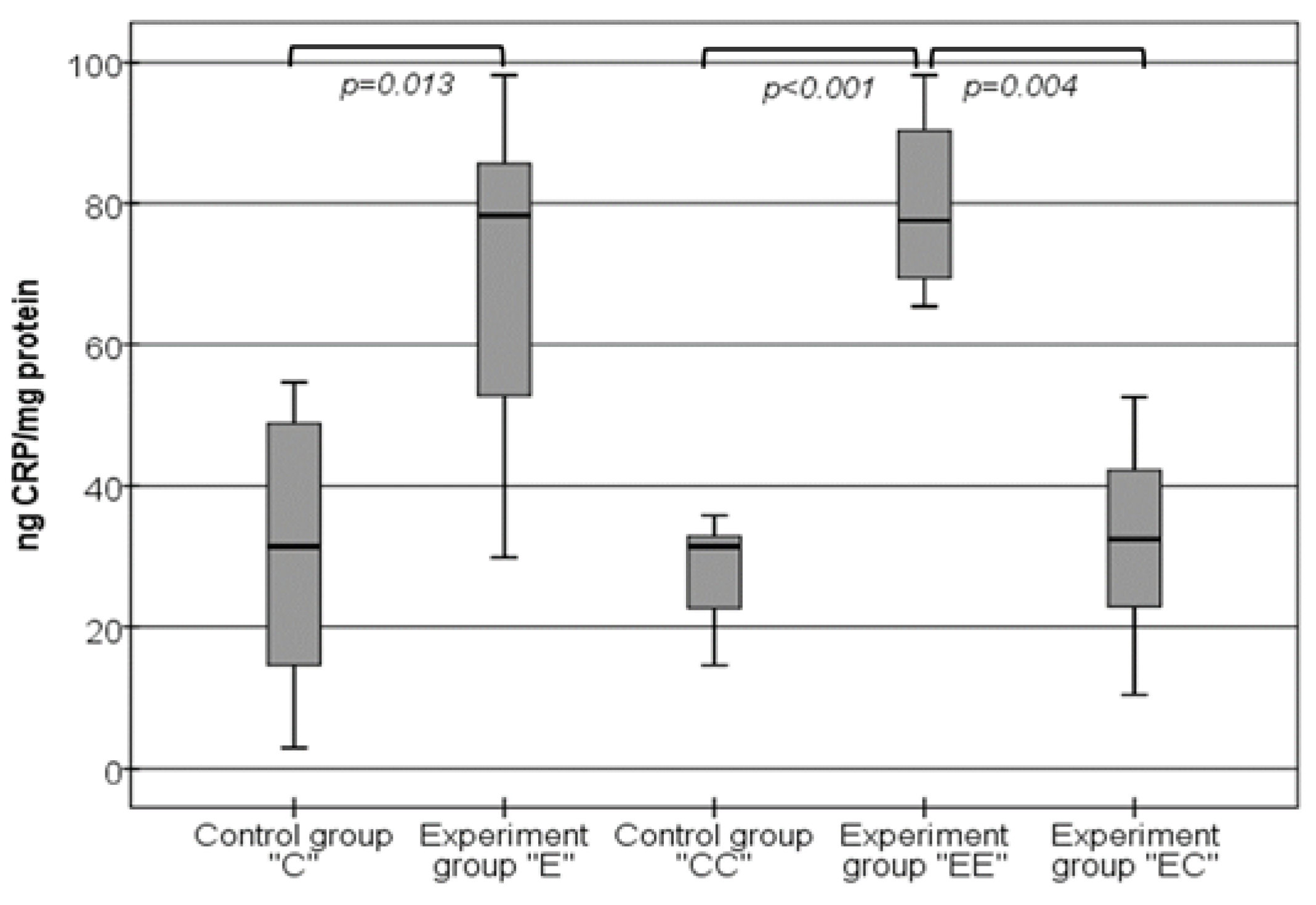
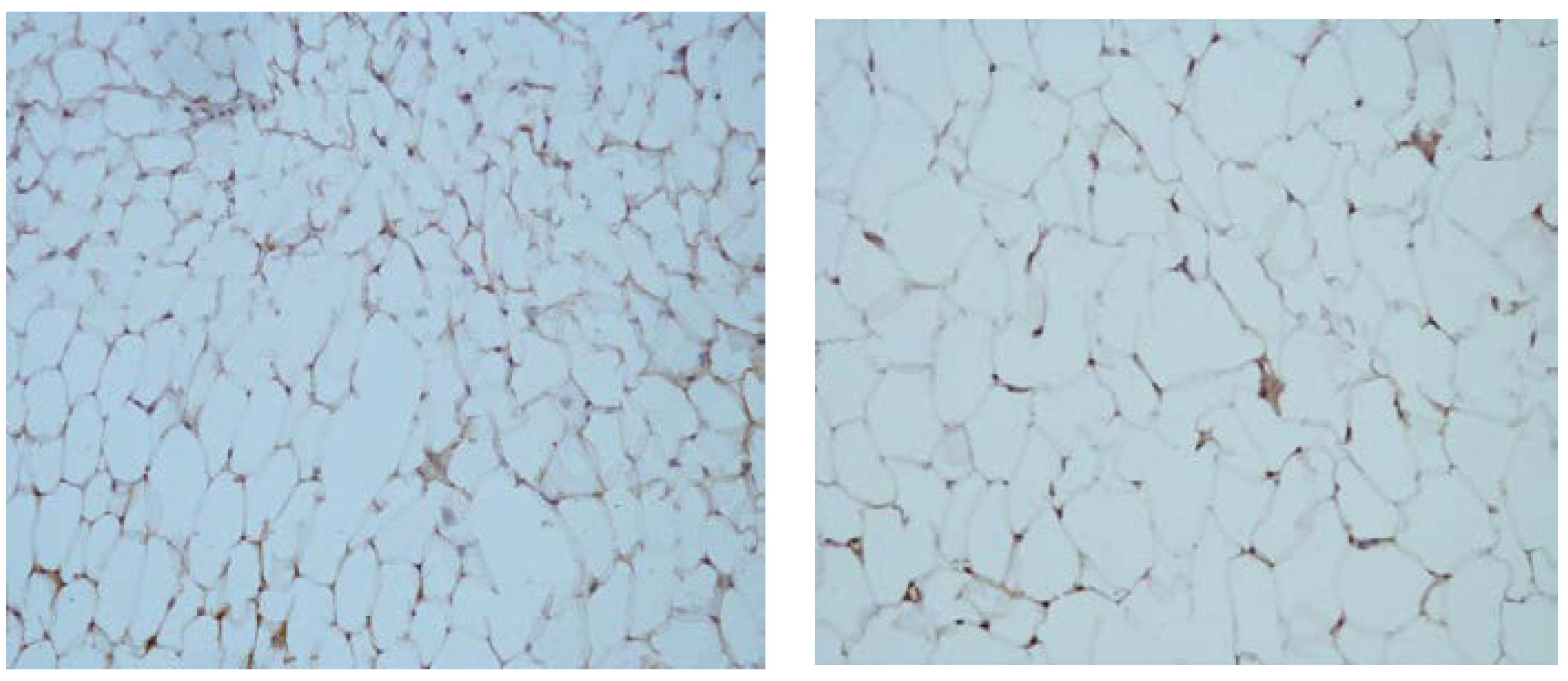
3.2. Qualitative Changes in the Levels of CRP in Tissue Homogenates from VAT
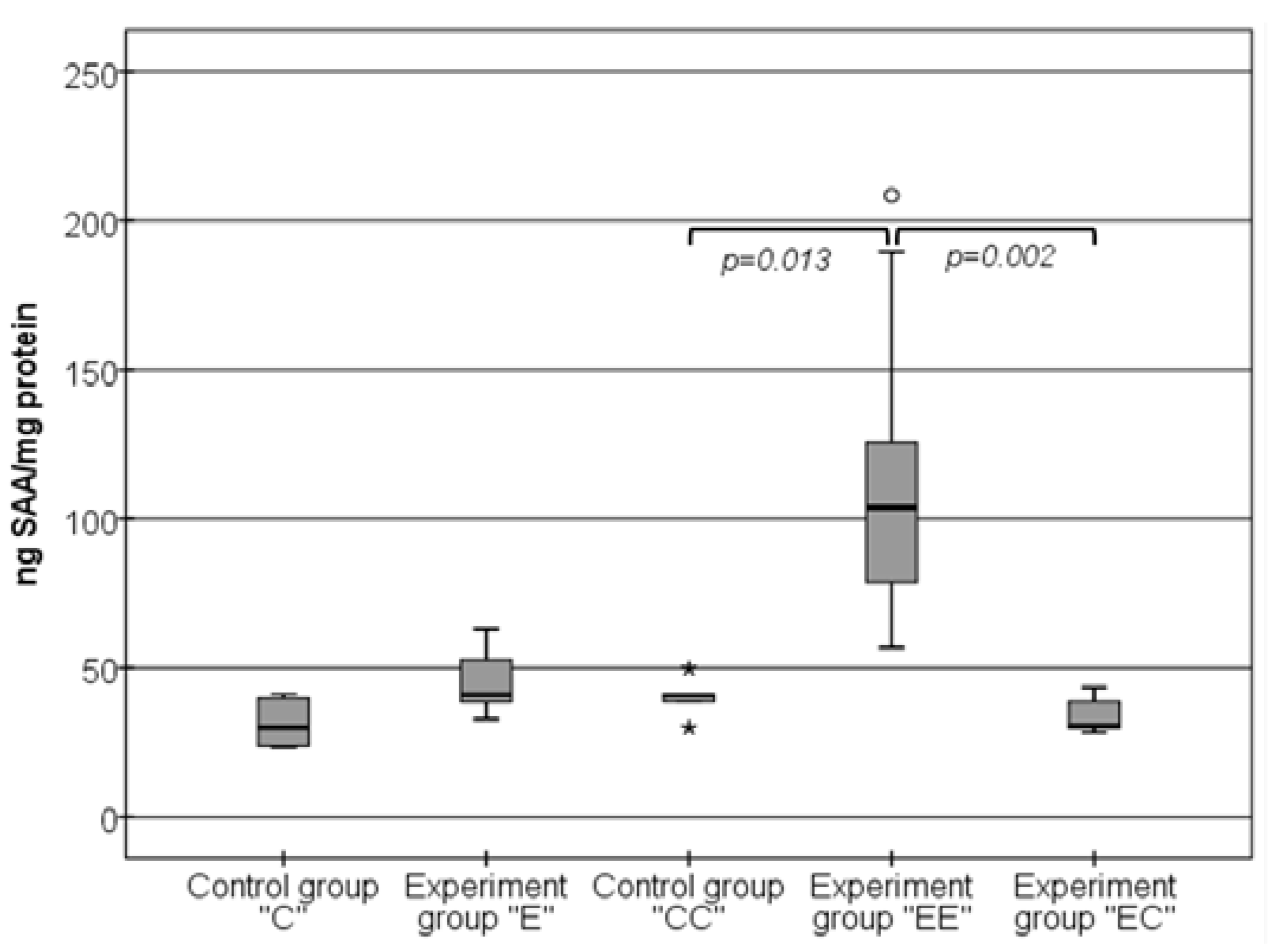
3.3. Changes in the Levels of SAA in Visceral Adipose Tissue Samples
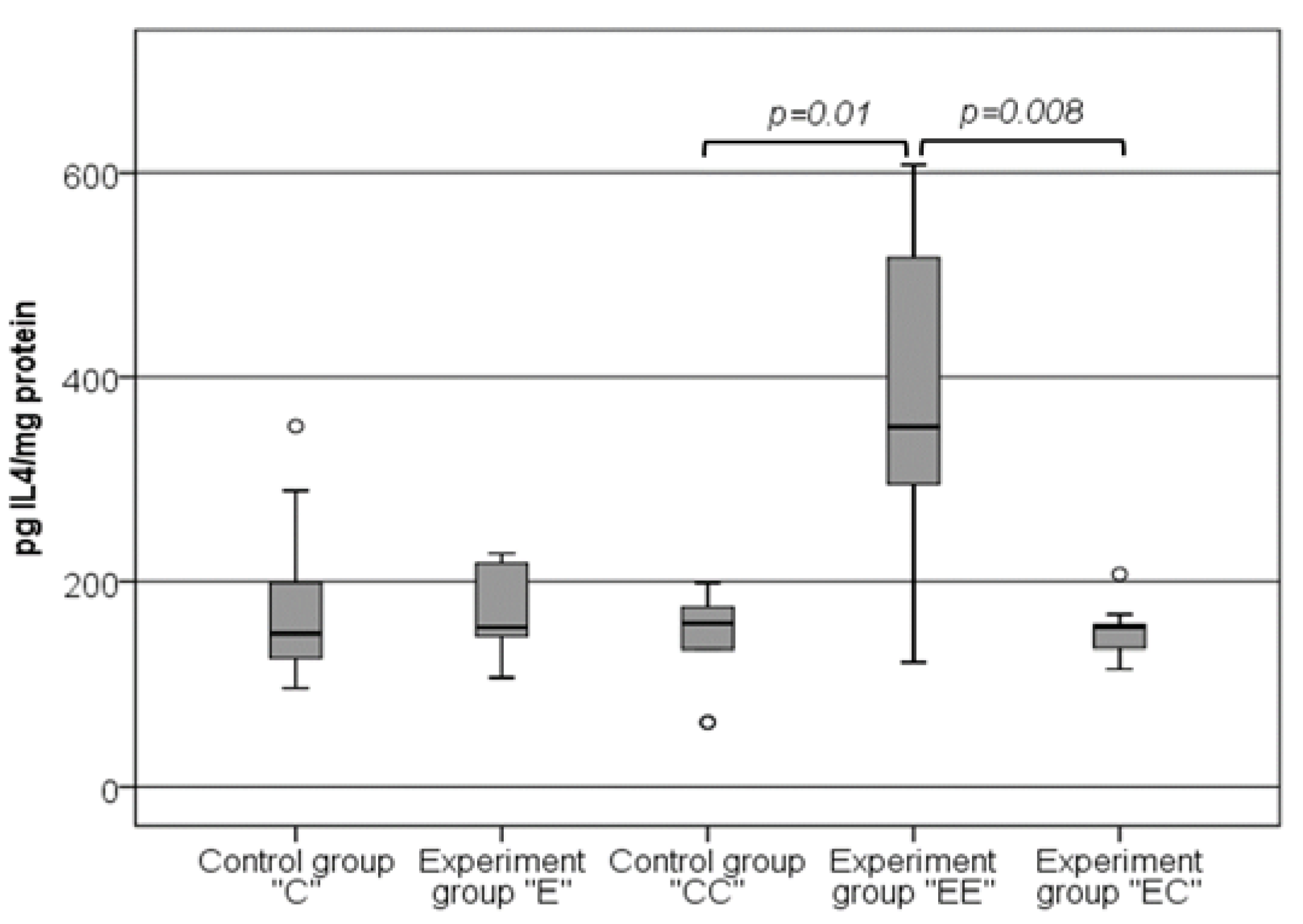
3.4. The Changes in the Levels of IL-4 in VAT Samples Follow the Same Pattern as the Changes in the Levels of SAA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ahima, R.S. Adipose Tissue as an Endocrine Organ. Obesity 2006, 14, 242S–249S. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Glass, C.K. Macrophages, Inflammation, and Insulin Resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Ferrante, A.W., Jr. Obesity-induced inflammation: A metabolic dialogue in the language of inflammation. J. Intern. Med. 2007, 262, 408–414. [Google Scholar] [CrossRef]
- Nappo, F.; Esposito, K.; Cioffi, M.; Giugliano, G.; Molinari, A.M.; Paolisso, G.; Marfella, R.; Giugliano, D. Postprandial endothelial activation in healthy subjects and in type 2 diabetic patients: Role of fat and carbohydrate meals. J. Am. Coll. Cardiol. 2002, 39, 1145–1150. [Google Scholar] [CrossRef]
- Xu, H.; Barnes, G.T.; Yang, Q.; Tan, G.; Yang, D.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jönsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S1–S78. [Google Scholar] [CrossRef]
- Cildir, G.; Akıncılar, S.C.; Tergaonkar, V. Chronic adipose tissue inflammation: All immune cells on the stage. Trends Mol. Med. 2013, 19, 487–500. [Google Scholar] [CrossRef]
- Lee, B.-C.; Lee, J. Cellular and molecular players in adipose tissue inflammation in the development of obesity-induced insulin resistance. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 446–462. [Google Scholar] [CrossRef]
- Alexopoulos, N.; Katritsis, D.; Raggi, P. Visceral adipose tissue as a source of inflammation and promoter of atherosclerosis. Atherosclerosis 2014, 233, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Park, J.Y.; Yu, R. Relationship of obesity and visceral adiposity with serum concentrations of CRP, TNF-alpha and IL-6. Diabetes Res. Clin. Pract. 2005, 69, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Bahceci, M.; Gokalp, D.; Bahceci, S.; Tuzcu, A.; Atmaca, S.; Arikan, S. The correlation between adiposity and adiponectin, tumor necrosis factor alpha, interleukin-6 and high sensitivity C-reactive protein levels. Is adipocyte size associated with inflammation in adults? J. Endocrinol. Investig. 2007, 30, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Poitou, C.; Divoux, A.; Faty, A.; Tordjman, J.; Hugol, D.; Aissat, A.; Keophiphath, M.; Henegar, C.; Commans, S.; Clément, K. Role of Serum Amyloid A in Adipocyte-Macrophage Cross Talk and Adipocyte Cholesterol Efflux. J. Clin. Endocrinol. Metab. 2009, 94, 1810–1817. [Google Scholar] [CrossRef] [PubMed]
- Czimmerer, Z.; Varga, T.; Poliska, S.; Nemet, I.; Szanto, A.; Nagy, L. Identification of novel markers of alternative activation and potential endogenous PPARγ ligand production mechanisms in human IL-4 stimulated differentiating macrophages. Immunobiology 2012, 217, 1301–1314. [Google Scholar] [CrossRef]
- Dimitrov, I.; Kamenov, V.; Angelova, P.; Petrov, M. Concentration of C-reactive protein in white adipose tissue, liver and blood serum of male Wistar rats. Trakia J. Sci. 2014, 12, 29–33. [Google Scholar]
- Dimitrov, I.V.; Kamenov, V.I.; Boyadjiev, N.P.; Georgieva, K.N.; Bivolarska, A.V.; Draganova-Filipova, M.N.; Angelova-Hristova, P.A.; Delchev, S.; Daskalova, E.; Gerginska, F.; et al. Impact of a High-fat Diet on the Development of Chronic Inflammation in Heart of Wistar rats. Folia Med. 2019, 61, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Amar, J.; Iglesias, M.A.; Poggi, M.; Knauf, C.; Bastelica, D.; Neyrinck, A.M.; Fava, F.; Tuohy, K.M.; Chabo, C.; et al. Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 2007, 56, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Fox, C.S.; Massaro, J.M.; Hoffmann, U.; Pou, K.M.; Maurovich-Horvat, P.; Liu, C.-Y.; Vasan, R.S.; Murabito, J.M.; Meigs, J.B.; Cupples, L.A.; et al. Abdominal Visceral and Subcutaneous Adipose Tissue Compartments: Association with metabolic risk factors in the Framingham Heart Study. Circulation 2007, 116, 39–48. [Google Scholar] [CrossRef]
- Bernardis, L.L.; Patterson, B.D. Correlation between ‘Lee index’ and carcass fat content in weanling and adult female rats with hypothalamic lesions. J. Endocrinol. 1968, 40, 527–528. [Google Scholar] [CrossRef]
- Stephens, D.N. Does the Lee obesity index measure general obesity? Physiol. Behav. 1980, 25, 313–315. [Google Scholar] [CrossRef] [PubMed]
- Valdez, R. A simple model-based index of abdominal adiposity. J. Clin. Epidemiol. 1991, 44, 955–956. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tong, G.; Tong, W.; Lu, L.; Qin, X. Can body mass index, waist circumference, waist-hip ratio and waist-height ratio predict the presence of multiple metabolic risk factors in Chinese subjects? BMC Public Health 2011, 11, 35. [Google Scholar] [CrossRef] [PubMed]
- Motamed, N.; Perumal, D.; Zamani, F.; Ashrafi, H.; Haghjoo, M.; Saeedian, F.; Maadi, M.; Akhavan-Niaki, H.; Rabiee, B.; Asouri, M. Conicity Index and Waist-to-Hip Ratio Are Superior Obesity Indices in Predicting 10-Year Cardiovascular Risk Among Men and Women. Clin. Cardiol. 2015, 38, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Shenoy, U. Influence of Central Obesity Assessed by Conicity Index on Lung Age in Young Adults. J. Clin. Diagn. Res. 2017, 11, CC09–CC12. [Google Scholar] [CrossRef] [PubMed]
- Woolcott, O.O.; Bergman, R.N. Relative fat mass (RFM) as a new estimator of whole-body fat percentage—A cross-sectional study in American adult individuals. Sci. Rep. 2018, 8, 10980. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Sun, S.; Xu, A.; Bhargava, P.; Yang, L.; Lam, K.S.L.; Gao, B.; Lee, C.-H.; Kersten, S.; Qi, L. Activation of Natural Killer T Cells Promotes M2 Macrophage Polarization in Adipose Tissue and Improves Systemic Glucose Tolerance via Interleukin-4 (IL-4)/STAT6 Protein Signaling Axis in Obesity. J. Biol. Chem. 2012, 287, 13561–13571. [Google Scholar] [CrossRef] [PubMed]
- Murano, I.; Barbatelli, G.; Parisani, V.; Latini, C.; Muzzonigro, G.; Castellucci, M.; Cinti, S. Dead adipocytes, detected as crown-like structures, are prevalent in visceral fat depots of genetically obese mice. J. Lipid Res. 2008, 49, 1562–1568. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation, metaflammation and immunometabolic disorders. Nature 2017, 542, 177–185. [Google Scholar] [CrossRef]
- Iyengar, N.M.; Gucalp, A.; Dannenberg, A.J.; Hudis, C.A. Obesity and Cancer Mechanisms: Tumor Microenvironment and Inflammation. J. Clin. Oncol. 2016, 34, 4270–4276. [Google Scholar] [CrossRef]
- Kaplan, M.; Hamoud, S.; Tendler, Y.; Meilin, E.; Lazarovitch, A.; Nitecki, S.; Hayek, T. A significant correlation between C—Reactive protein levels in blood monocytes derived macrophages versus content in carotid atherosclerotic lesions. J. Inflamm. 2014, 11, 7. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, M.; Shur, A.; Tendler, Y. M1 Macrophages but Not M2 Macrophages Are Characterized by Upregulation of CRP Expression via Activation of NFκB: A Possible Role for Ox-LDL in Macrophage Polarization. Inflammation 2018, 41, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Calabro, P.; Chang, D.W.; Willerson, J.T.; Yeh, E.T. Release of C-Reactive Protein in Response to Inflammatory Cytokines by Human Adipocytes: Linking Obesity to Vascular Inflammation. J. Am. Coll. Cardiol. 2005, 46, 1112–1113. [Google Scholar] [CrossRef]
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-sensitive obesity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E506–E515. [Google Scholar] [CrossRef]
- Sjöholm, K.; Palming, J.; Olofsson, L.E.; Gummesson, A.; Svensson, P.-A.; Lystig, T.C.; Jennische, E.; Brandberg, J.; Torgerson, J.S.; Carlsson, B.; et al. A Microarray Search for Genes Predominantly Expressed in Human Omental Adipocytes: Adipose Tissue as a Major Production Site of Serum Amyloid A. J. Clin. Endocrinol. Metab. 2005, 90, 2233–2239. [Google Scholar] [CrossRef]
- Ridker, P.M. Clinical Application of C-Reactive Protein for Cardiovascular Disease Detection and Prevention. Circulation 2003, 107, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Braig, D.; Nero, T.L.; Koch, H.-G.; Kaiser, B.; Wang, X.; Thiele, J.R.; Morton, C.J.; Zeller, J.; Kiefer, J.; Potempa, L.A.; et al. Transitional changes in the CRP structure lead to the exposure of proinflammatory binding sites. Nat. Commun. 2017, 8, 14188. [Google Scholar] [CrossRef]
- Thiele, J.R.; Habersberger, J.; Braig, D.; Schmidt, Y.; Goerendt, K.; Maurer, V.; Bannasch, H.; Scheichl, A.; Woollard, K.J.; von Dobschütz, E.; et al. Dissociation of pentameric to monomeric C-reactive protein localizes and aggravates inflammation: In vivo proof of a powerful proinflammatory mechanism and a new anti-inflammatory strategy. Circulation 2014, 130, 35–50. [Google Scholar] [CrossRef]
- van der Westhuyzen, D.R.; de Beer, F.C.; Webb, N.R. HDL cholesterol transport during inflammation. Curr. Opin. Infect. Lipidol. 2007, 18, 147–151. [Google Scholar] [CrossRef]
- Poitou, C.; Coussieu, C.; Rouault, C.; Coupaye, M.; Cancello, R.; Bedel, J.; Gouillon, M.; Bouillot, J.; Oppert, J.; Basdevant, A.; et al. Serum Amyloid A: A Marker of Adiposity-induced Low-grade Inflammation but Not of Metabolic Status. Obesity 2006, 14, 309–318. [Google Scholar] [CrossRef]
- Poitou, C.; Viguerie, N.; Cancello, R.; De Matteis, R.; Cinti, S.; Stich, V.; Coussieu, C.; Gauthier, E.; Courtine, M.; Zucker, J.D.; et al. Serum amyloid A: Production by human white adipocyte and regulation by obesity and nutrition. Diabetologia 2005, 48, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.; Trumbauer, A.C.; Noffsinger, V.P.; Jeon, H.; Patrick, A.C.; De Beer, F.C.; Webb, N.R.; Tannock, L.R.; Shridas, P. Serum Amyloid A is not obligatory for high-fat, high-sucrose, cholesterol-fed diet-induced obesity and its metabolic and inflammatory complications. PLoS ONE 2022, 17, e0266688. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.-Z.; Lee, M.-J.; Hu, H.; I Pollin, T.; Ryan, A.S.; Nicklas, B.J.; Snitker, S.; Horenstein, R.B.; Hull, K.; Goldberg, N.H.; et al. Acute-Phase Serum Amyloid A: An Inflammatory Adipokine and Potential Link between Obesity and Its Metabolic Complications. PLoS Med. 2006, 3, e287. [Google Scholar] [CrossRef] [PubMed]
- Ahlin, S.; Olsson, M.; Olsson, B.; Svensson, P.-A.; Sjöholm, K. No Evidence for a Role of Adipose Tissue-Derived Serum Amyloid A in the Development of Insulin Resistance or Obesity-Related Inflammation in hSAA1(+/−) Transgenic Mice. PLoS ONE 2013, 8, e72204. [Google Scholar] [CrossRef] [PubMed]
- Lynch, L.; O’Shea, D.; Winter, D.C.; Geoghegan, J.; Doherty, D.G.; O’Farrelly, C. Invariant NKT cells and CD1d+ cells amass in human omentum and are depleted in patients with cancer and obesity. Eur. J. Immunol. 2009, 39, 1893–1901. [Google Scholar] [CrossRef] [PubMed]
- Tsao, C.-H.; Shiau, M.-Y.; Chuang, P.-H.; Chang, Y.-H.; Hwang, J. Interleukin-4 regulates lipid metabolism by inhibiting adipogenesis and promoting lipolysis. J. Lipid Res. 2014, 55, 385–397. [Google Scholar] [CrossRef]
- Kang, K.; Reilly, S.M.; Karabacak, V.; Gangl, M.R.; Fitzgerald, K.; Hatano, B.; Lee, C.-H. Adipocyte-Derived Th2 Cytokines and Myeloid PPARdelta Regulate Macrophage Polarization and Insulin Sensitivity. Cell Metab. 2008, 7, 485–495. [Google Scholar] [CrossRef] [PubMed]
- Mito, N.; Hosoda, T.; Kato, C.; Sato, K. Change of cytokine balance in diet-induced obese mice. Metabolism 2000, 49, 1295–1300. [Google Scholar] [CrossRef] [PubMed]
- Borst, S.E.; Conover, C.F.; Bagby, G.J. Association of resistin with visceral fat and muscle insulin resistance. Cytokine 2005, 32, 39–44. [Google Scholar] [CrossRef]
- Poret, J.M.; Souza-Smith, F.; Marcell, S.J.; Gaudet, D.A.; Tzeng, T.H.; Braymer, H.D.; Harrison-Bernard, L.M.; Primeaux, S.D. High fat diet consumption differentially affects adipose tissue inflammation and adipocyte size in obesity-prone and obesity-resistant rats. Int. J. Obes. 2018, 42, 535–541. [Google Scholar] [CrossRef]
- McKernan, K.; Varghese, M.; Patel, R.; Singer, K. Role of TLR4 in the induction of inflammatory changes in adipocytes and macrophages. Adipocyte 2020, 9, 212–222. [Google Scholar] [CrossRef]
- Fessler, M.B.; Rudel, L.L.; Brown, J.M. Toll-like receptor signaling links dietary fatty acids to the metabolic syndrome. Curr. Opin. Lipidol. 2009, 20, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Kokoeva, M.V.; Inouye, K.; Tzameli, I.; Yin, H.; Flier, J.S. TLR4 links innate immunity and fatty acid–induced insulin resistance. J. Clin. Investig. 2006, 116, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Tanimoto-Koyama, K.; Nishida, J.; Itoh, M.; Yuan, X.; Mizuarai, S.; Kotani, H.; Yamaoka, S.; Miyake, K.; Aoe, S.; et al. Role of the Toll-like receptor 4/NF-κB pathway in saturated fatty acid-induced inflammatory changes in the interaction between adipocytes and macrophages. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 84–91. [Google Scholar] [CrossRef]
- Weatherill, A.R.; Lee, J.Y.; Zhao, L.; Lemay, D.G.; Youn, H.S.; Hwang, D.H. Saturated and polyunsaturated fatty acids recip-rocally modulate dendritic cell functions mediated through TLR4. J. Immunol. 2005, 174, 5390–5397. [Google Scholar] [CrossRef] [PubMed]
- Nanri, A.; Moore, M.A.; Kono, S. Impact of C-reactive protein on disease risk and its relation to dietary factors. Asian Pac. J. Cancer Prev. 2007, 8, 167–177. [Google Scholar] [PubMed]
- Joffe, Y.T.; Collins, M.; Goedecke, J.H. The Relationship between Dietary Fatty Acids and Inflammatory Genes on the Obese Phenotype and Serum Lipids. Nutrients 2013, 5, 1672–1705. [Google Scholar] [CrossRef]
- Vila, I.K.; Badin, P.-M.; Marques, M.-A.; Monbrun, L.; Lefort, C.; Mir, L.; Louche, K.; Bourlier, V.; Roussel, B.; Gui, P.; et al. Immune Cell Toll-like Receptor 4 Mediates the Development of Obesity- and Endotoxemia-Associated Adipose Tissue Fibrosis. Cell Rep. 2014, 7, 1116–1129. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| n | Weight (g): Mean ± SD | BMI (kg/m2) Mean ± SD | Lee Index (g/mm) Mean ± SD | CI (kg/m) Mean ± SD | RFM (kg/m) Mean ± SD | |
|---|---|---|---|---|---|---|
| Initial | 60 | 145.42 ± 15.26 | – | – | – | – |
| Group “C” | 30 | 295.88 ± 31.88 | 7.26 ± 0.74 | 329.80 ± 14 | 1.89 ± 0.15 | 40.3 ± 1.06 |
| Group “E” | 30 | 321.50 ± 24.02 ** | 7.85 ± 0.87 * | 332.05 ± 20.62 | 2.18 ± 0.15 **# | 40.52 ± 1.31 |
| Group “CC” | 8 | 331.38 ± 27.51 | 6.45 ± 0.32 | 306.92 ± 5.77 | 1.92 ± 0.13 | 38.37 ± 1.1 |
| Group “EE” | 8 | 360.13 ± 23.30 | 7.03 ± 0.65 | 317.16 ± 9.42 | 2.05 ± 0.07 | 39.41 ± 1.11 |
| Group “EC” | 8 | 341.63 ± 51.60 | 6.80 ± 0.28 | 313.59 ± 6.18 | 1.98 ± 0.15 | 38.95 ± 0.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimitrov, I.; Stankova, T.; Angelova, P.; Boyadjiev, N.; Georgieva, K.; Dimov, I.; Bivolarska, A.; Draganova, M.; Gerginska, F.; Daskalova, E.; et al. Diet-Induced Early Inflammatory Response of Visceral Adipose Tissue in Healthy Male Wistar Rats. Nutrients 2024, 16, 1184. https://doi.org/10.3390/nu16081184
Dimitrov I, Stankova T, Angelova P, Boyadjiev N, Georgieva K, Dimov I, Bivolarska A, Draganova M, Gerginska F, Daskalova E, et al. Diet-Induced Early Inflammatory Response of Visceral Adipose Tissue in Healthy Male Wistar Rats. Nutrients. 2024; 16(8):1184. https://doi.org/10.3390/nu16081184
Chicago/Turabian StyleDimitrov, Iliyan, Teodora Stankova, Penka Angelova, Nikolay Boyadjiev, Katerina Georgieva, Ivica Dimov, Anelia Bivolarska, Milena Draganova, Fanka Gerginska, Elena Daskalova, and et al. 2024. "Diet-Induced Early Inflammatory Response of Visceral Adipose Tissue in Healthy Male Wistar Rats" Nutrients 16, no. 8: 1184. https://doi.org/10.3390/nu16081184
APA StyleDimitrov, I., Stankova, T., Angelova, P., Boyadjiev, N., Georgieva, K., Dimov, I., Bivolarska, A., Draganova, M., Gerginska, F., Daskalova, E., Gramatikov, V., & Delchev, S. (2024). Diet-Induced Early Inflammatory Response of Visceral Adipose Tissue in Healthy Male Wistar Rats. Nutrients, 16(8), 1184. https://doi.org/10.3390/nu16081184