
Allergenic Shrimp Tropomyosin Distinguishes from a Non-Allergenic Chicken Homolog by Pronounced Intestinal Barrier Disruption and Downstream Th2 Responses in Epithelial and Dendritic Cell (Co)Culture

 ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Allergenic Proteins
2.2. Caco-2, HT-29MTX and HT-29 Cell Culture
2.3. Transepithelial Electrical Resistance (TEER) and Cytotoxicity Assay
2.4. Caco-2 and PBMC Coculture
2.5. HT-29-moDC or moDC and Sequential DC/T Cell Coculture
2.6. Cytokine Measurements
2.7. Flow Cytometry
2.8. Gene Transcription Analysis
2.9. Statistics
3. Results
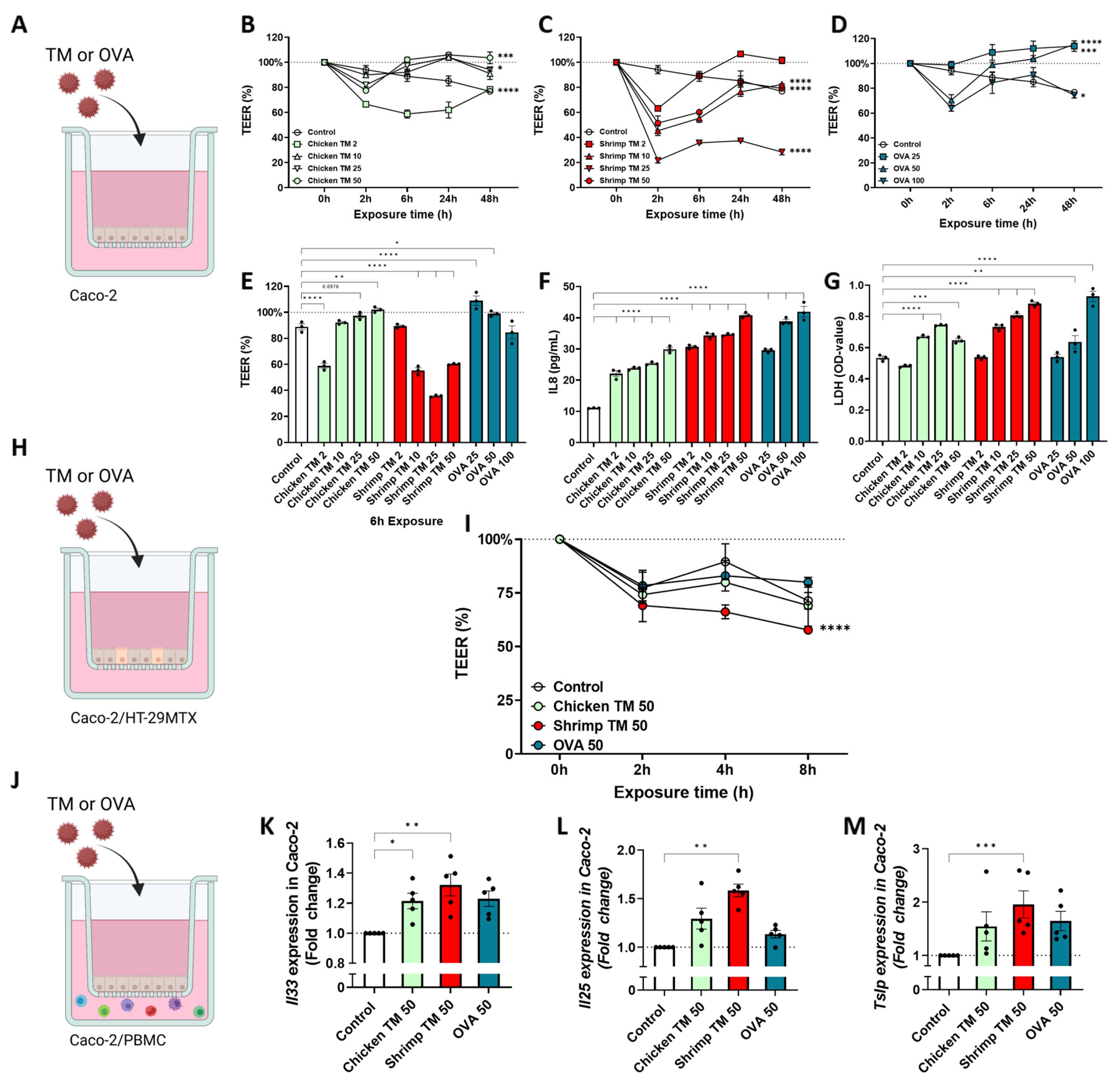
3.1. Shrimp TM Decreases Epithelial Barrier and Induces Expression of Alarmins
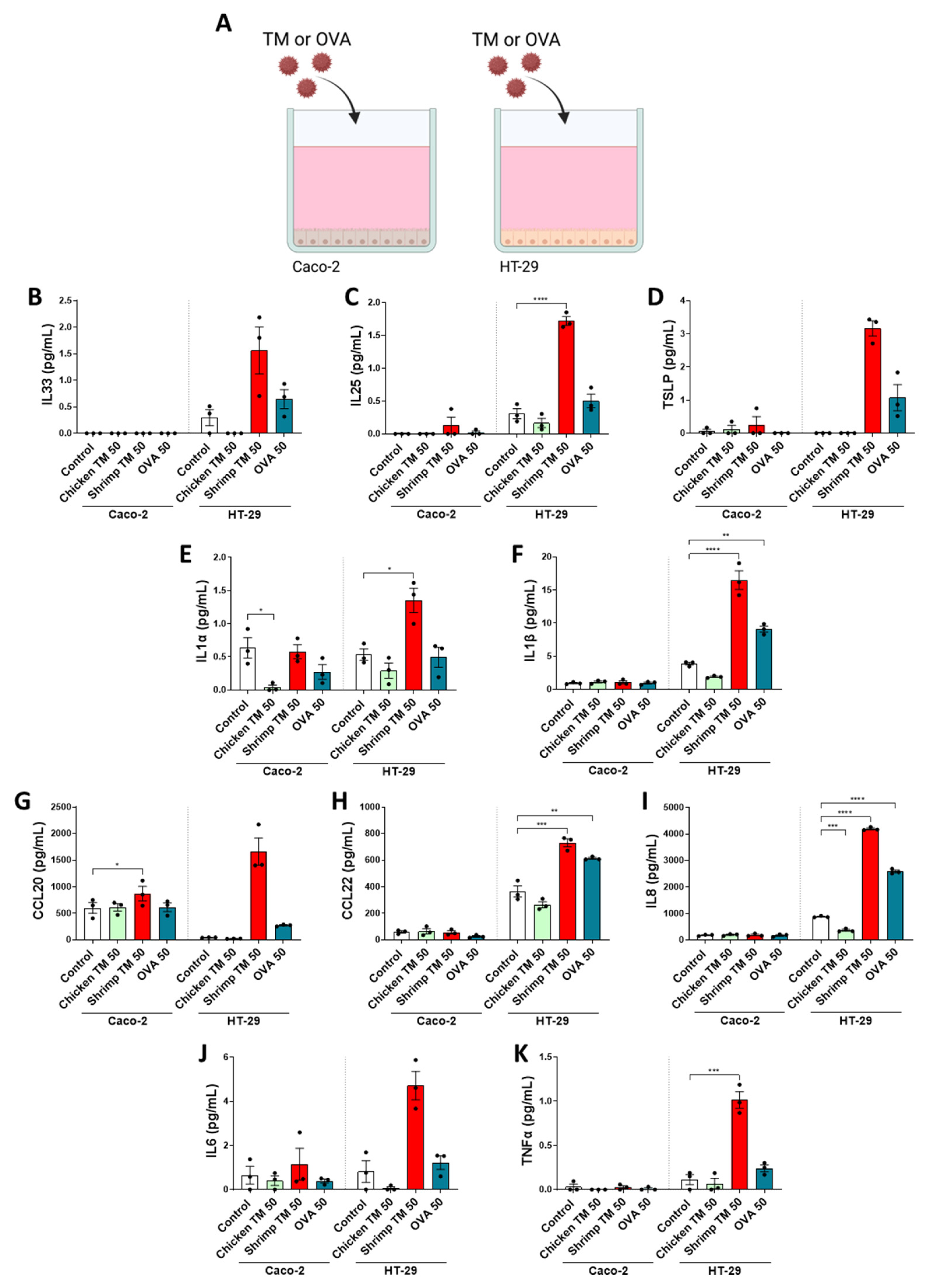
3.2. Cytokine and Chemokine Secretion Is Enhanced in HT-29 Cells after Allergen Exposure
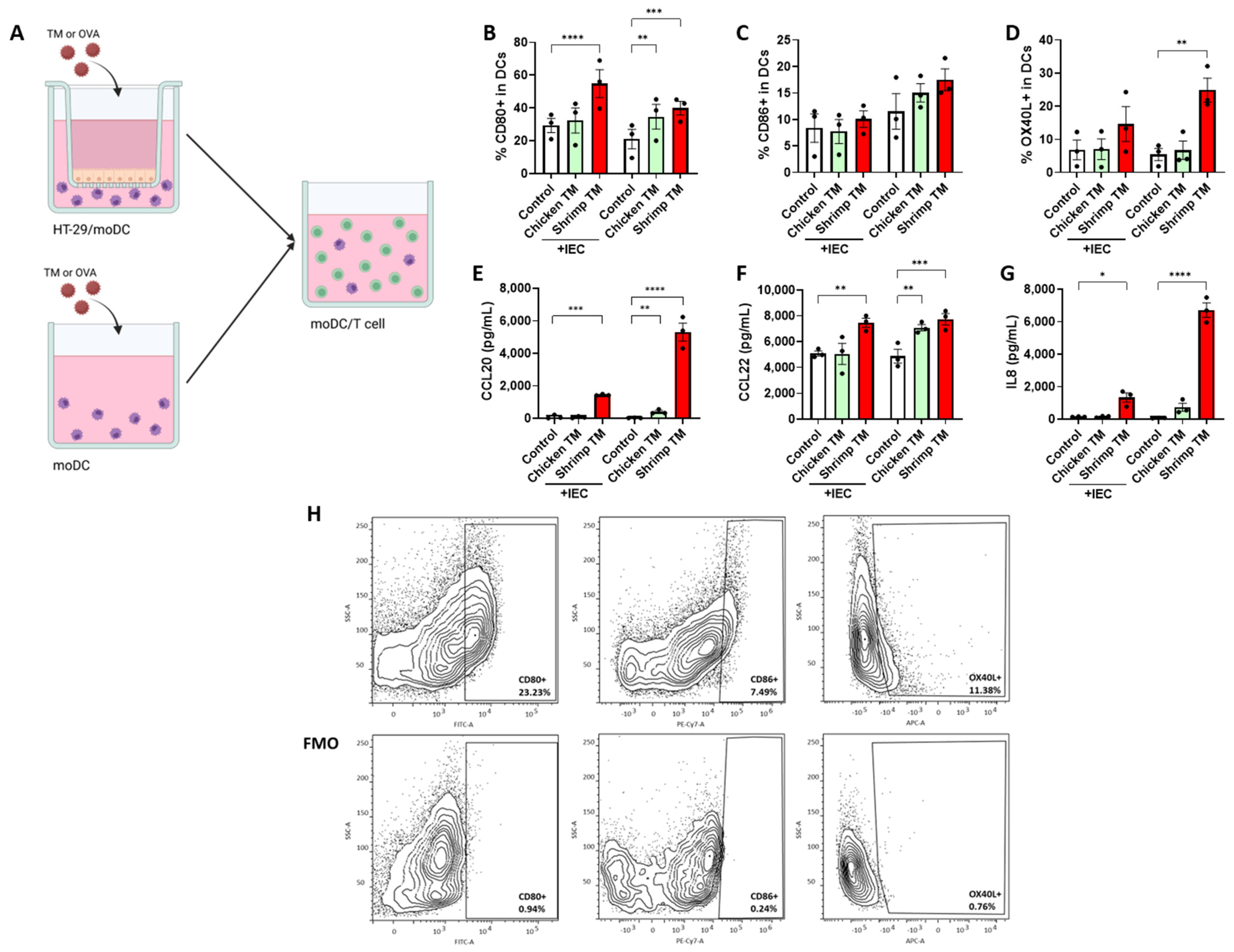
3.3. Shrimp TM Induces moDC Activation in Presence and Absence of IEC
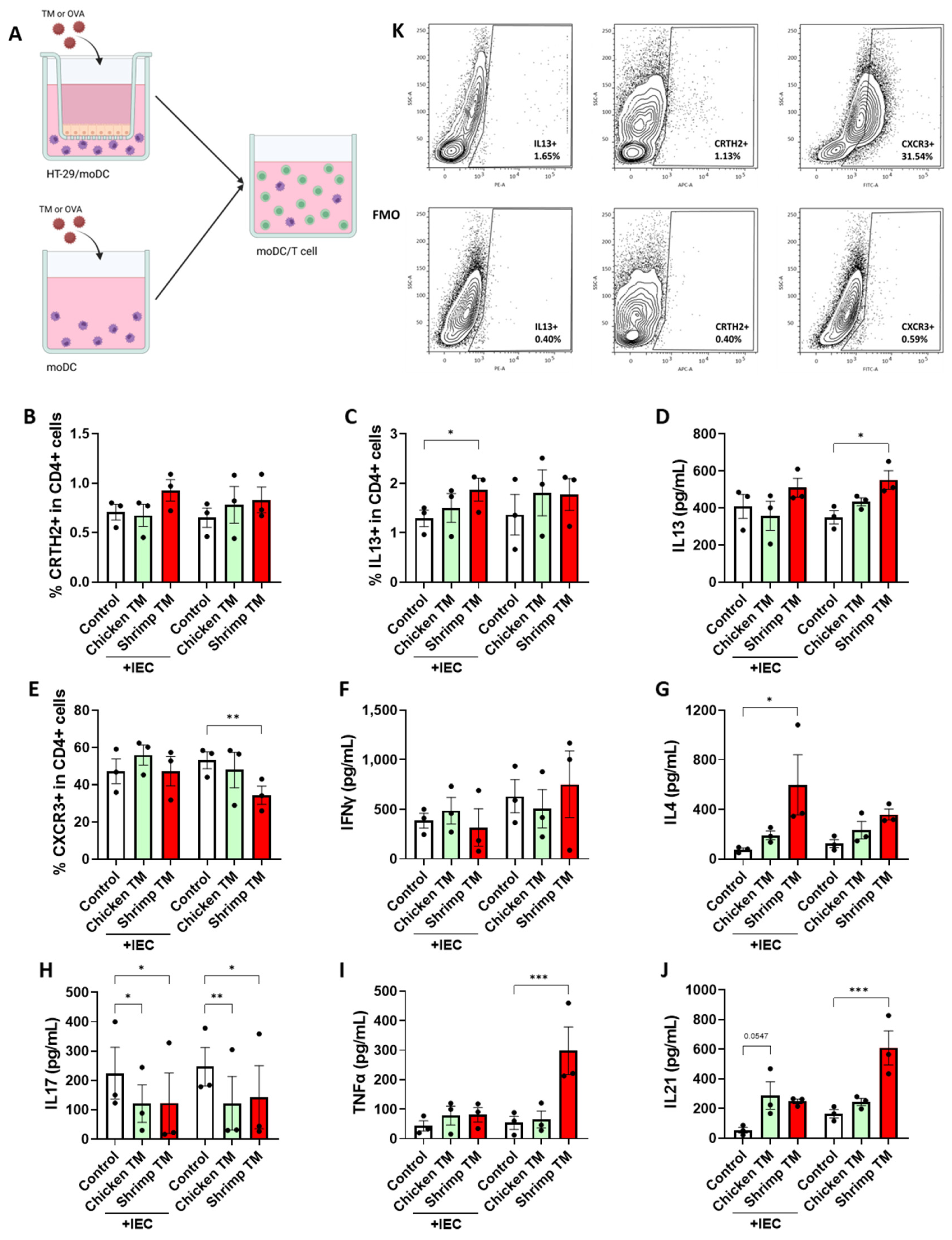
3.4. Shrimp TM Promotes a Type 2 T Cell Response via moDC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Spolidoro, G.C.I.; Amera, Y.T.; Ali, M.M.; Nyassi, S.; Lisik, D.; Ioannidou, A.; Rovner, G.; Khaleva, E.; Venter, C.; van Ree, R.; et al. Frequency of food allergy in Europe: An updated systematic review and meta-analysis. Allergy Eur. J. Allergy Clin. Immunol. 2022, 78, 351–368. [Google Scholar] [CrossRef] [PubMed]
- Loh, W.; Tang, M.L.K. The epidemiology of food allergy in the global context. Int. J. Environ. Res. Public Health 2018, 15, 2043. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, H.; Yokooji, T.; Taogoshi, T. Common food allergens and their IgE-binding epitopes. Allergol. Int. 2015, 64, 332–343. [Google Scholar] [CrossRef] [PubMed]
- Ménard, S.; Cerf-Bensussan, N.; Heyman, M. Multiple facets of intestinal permeability and epithelial handling of dietary antigens. Mucosal Immunol. 2010, 3, 247–259. [Google Scholar] [CrossRef] [PubMed]
- Tordesillas, L.; Gómez-Casado, C.; Garrido-Arandia, M.; Murua-García, A.; Palacín, A.; Varela, J.; Konieczna, P.; Cuesta-Herranz, J.; Akdis, C.A.; O’Mahony, L.; et al. Transport of Pru p 3 across gastrointestinal epithelium—An essential step towards the induction of food allergy? Clin. Exp. Allergy 2013, 43, 1374–1383. [Google Scholar] [CrossRef] [PubMed]
- Dhanapala, P.; De Silva, C.; Doran, T.; Suphioglu, C. Cracking the egg: An insight into egg hypersensitivity. Mol. Immunol. 2015, 66, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Smits, M.; Nooijen, I.; Redegeld, F.; de Jong, A.; Le, T.; Knulst, A.; Houben, G.; Verhoeckx, K. Digestion and Transport across the Intestinal Epithelium Affects the Allergenicity of Ara h 1 and 3 but Not of Ara h 2 and 6. Mol. Nutr. Food Res. 2021, 65, e2000712. [Google Scholar] [CrossRef] [PubMed]
- Niewiem, M.; Grzybowska-Chlebowczyk, U. Intestinal Barrier Permeability in Allergic Diseases. Nutrients 2022, 14, 1893. [Google Scholar] [CrossRef] [PubMed]
- Dijk, W.; Villa, C.; Benedé, S.; Vassilopoulou, E.; Mafra, I.; Garrido-Arandia, M.; Blanco, M.M.; Bouchaud, G.; Hoppenbrouwers, T.; Bavaro, S.L.; et al. Critical features of an in vitro intestinal absorption model to study the first key aspects underlying food allergen sensitization. Compr. Rev. Food Sci. Food Saf. 2023, 22, 971–1005. [Google Scholar] [CrossRef]
- Ballegaard, A.S.R.; Bøgh, K.L. Intestinal protein uptake and IgE-mediated food allergy. Food Res. Int. 2023, 163, 112150. [Google Scholar] [CrossRef] [PubMed]
- Mazzucchelli, G.; Holzhauser, T.; Velickovic, T.C.; Diaz-Perales, A.; Molina, E.; Roncada, P.; Rodrigues, P.; Verhoeckx, K.; Hoffmann-Sommergruber, K. Current (Food) Allergenic Risk Assessment: Is It Fit for Novel Foods? Status Quo and Identification of Gaps. Mol. Nutr. Food Res. 2018, 62, 1700278. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.; Broekman, H.; Knulst, A.; Houben, G. Allergenicity assessment strategy for novel food proteins and protein sources. Regul. Toxicol. Pharmacol. 2016, 79, 118–124. [Google Scholar] [CrossRef] [PubMed]
- El-Mezgueldi, M. Tropomyosin dynamics. J. Muscle Res. Cell Motil. 2014, 35, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Reese, G.; Ayuso, R.; Lehrer, S.B. Tropomyosin: An invertebrate pan-allergen. Int. Arch. Allergy Immunol. 1999, 119, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.H.; Wang, H.; Sun, D.W. An overview of tropomyosin as an important seafood allergen: Structure, cross-reactivity, epitopes, allergenicity, and processing modifications. Compr. Rev. Food Sci. Food Saf. 2022, 21, 127–147. [Google Scholar] [CrossRef] [PubMed]
- Ruethers, T.; Taki, A.C.; Johnston, E.; Nugraha, R.; Le, T.T.K.; Kalic, T.; McLean, T.; Kamath, S.D.; Lopata, A.L. Seafood allergy: A comprehensive review of fish and shellfish allergens. Mol. Immunol. 2018, 100, 28–57. [Google Scholar] [CrossRef] [PubMed]
- Klueber, J.; Costa, J.; Randow, S.; Codreanu-Morel, F.; Verhoeckx, K.; Bindslev-Jensen, C.; Ollert, M.; Hoffmann-Sommergruber, K.; Morisset, M.; Holzhauser, T.; et al. Homologous tropomyosins from vertebrate and invertebrate: Recombinant calibrator proteins in functional biological assays for tropomyosin allergenicity assessment of novel animal foods. Clin. Exp. Allergy 2020, 50, 105. [Google Scholar] [CrossRef] [PubMed]
- Van Bilsen, J.H.; Sienkiewicz-Szłapka, E.; Lozano-Ojalvo, D.; Willemsen, L.E.; Antunes, C.M.; Molina, E.; Smit, J.J.; Wróblewska, B.; Wichers, H.J.; Knol, E.F.; et al. Application of the adverse outcome pathway (AOP) concept to structure the available in vivo and in vitro mechanistic data for allergic sensitization to food proteins. Clin. Transl. Allergy 2017, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Zuurveld, M.; Díaz, C.B.; Redegeld, F.; Folkerts, G.; Garssen, J.; van’t Land, B.; Willemsen, L.E. An advanced in vitro human mucosal immune model to predict food sensitizing allergenicity risk: A proof of concept using ovalbumin as model allergen. Front. Immunol. 2023, 13, 1073034. [Google Scholar] [CrossRef]
- Popp, J.; Trendelenburg, V.; Niggemann, B.; Randow, S.; Völker, E.; Vogel, L.; Reuter, A.; Spiric, J.; Schiller, D.; Beyer, K.; et al. Pea (Pisum sativum) allergy in children: Pis s 1 is an immunodominant major pea allergen and presents IgE binding sites with potential diagnostic value. Clin. Exp. Allergy 2020, 50, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Zuurveld, M.; Kiliaan, P.C.; van Grinsven, S.E.; Folkerts, G.; Garssen, J.; van‘t Land, B.; Willemsen, L.E. Ovalbumin induced epithelial activation directs moDC to instruct type 2 inflammation in T cells which is differentially modulated by 2′-fucosyllactose and 3-fucosyllactose. J. Innate Immun. 2022, 15, 222–239. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rodríguez, L.; Lozano-Ojalvo, D.; Menchén-Martínez, D.; Molina, E.; López-Fandiño, R.; Benedé, S. Egg yolk lipids induce sensitization to egg white proteins in a mouse model without adjuvant and exacerbate Th2 responses to egg white in cells from allergic patients. Food Res. Int. 2023, 172, 112669. [Google Scholar] [CrossRef] [PubMed]
- Tordesillas, L.; Goswami, R.; Benedé, S.; Grishina, G.; Dunkin, D.; Järvinen, K.M.; Maleki, S.J.; Sampson, H.A.; Berin, M.C. Skin exposure promotes a Th2-dependent sensitization to peanut allergens. J. Clin. Investig. 2014, 124, 4965–4975. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Angkasekwinai, P.; Lu, N.; Voo, K.S.; Arima, K.; Hanabuchi, S.; Hippe, A.; Corrigan, C.J.; Dong, C.; Homey, B.; et al. IL-25 augments type 2 immune responses by enhancing the expansion and functions of TSLP-DC-activated Th2 memory cells. J. Exp. Med. 2007, 204, 1837–1847. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.L.; Eigenmann, P.A.; Sicherer, S.H. Clinical Relevance of Cross-Reactivity in Food Allergy. J. Allergy Clin. Immunol. Pract. 2021, 9, 82–99. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Lin, W.; Wang, C.; Wang, Y. Establishment of a 3-Dimensional Intestinal Cell Model to Simulate the Intestinal Mucosal Immune System for Food Allergy Investigations. Front. Immunol. 2022, 13, 853443. [Google Scholar] [CrossRef]
- Yu, L.C.H. Intestinal Epithelial Barrier Dysfunction in Food Hypersensitivity. J. Allergy 2012, 2012, 596081. [Google Scholar] [CrossRef] [PubMed]
- Kubo, A.; Nagao, K.; Amagai, M. Epidermal barrier dysfunction and cutaneous sensitization in atopic diseases. J. Clin. Investig. 2012, 122, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Akdis, C.A. Does the epithelial barrier hypothesis explain the increase in allergy, autoimmunity and other chronic conditions? Nat. Rev. Immunol. 2021, 21, 739–751. [Google Scholar] [CrossRef] [PubMed]
- Goleva, E.; Berdyshev, E.; Leung, D.Y.M. Epithelial barrier repair and prevention of allergy. J. Clin. Investig. 2019, 1, 1463–1474. [Google Scholar] [CrossRef] [PubMed]
- Hellings, P.W.; Steelant, B. Epithelial barriers in allergy and asthma. J. Allergy Clin. Immunol. 2020, 145, 1499. [Google Scholar] [CrossRef] [PubMed]
- Sander, G.R.; Cummins, A.G.; Powell, B.C. Rapid disruption of intestinal barrier function by gliadin involves altered expression of apical junctional proteins. FEBS Lett. 2005, 579, 4851–4855. [Google Scholar] [CrossRef] [PubMed]
- Price, D.B.; Ackland, M.L.; Burks, W.; Knight, M.I.; Suphioglu, C. Peanut allergens alter intestinal barrier permeability and tight junction localisation in Caco-2 cell cultures. Cell. Physiol. Biochem. 2014, 33, 1758–1777. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.M.; Čavić, M.; Nešić, A.; Andjelković, U.; Akbari, P.; Smit, J.J.; Gavrović-Jankulović, M. Kiwifruit cysteine protease actinidin compromises the intestinal barrier by disrupting tight junctions. Biochim. Biophys. Acta 2016, 1860, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lin, W.; Wang, Y.; Fu, L. Suppression of Hippo Pathway by Food Allergen Exacerbates Intestinal Epithelia Instability and Facilitates Hypersensitivity. Mol. Nutr. Food Res. 2021, 65, 2000593. [Google Scholar] [CrossRef] [PubMed]
- Koda, T.; Minami, H.; Ogawa, T.; Yamano, M.; Takeda, E. Higher concentrations of interferon-gamma enhances uptake and transport of dietary antigens by human intestinal cells: A study using cultured Caco-2 cells. J. Nutr. Sci. Vitaminol. 2003, 49, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Khuda, S.E.; Nguyen, A.V.; Sharma, G.M.; Alam, M.S.; Balan, K.V.; Williams, K.M. Effects of Emulsifiers on an In vitro Model of Intestinal Epithelial Tight Junctions and the Transport of Food Allergens. Mol. Nutr. Food Res. 2022, 66, 2100576. [Google Scholar] [CrossRef] [PubMed]
- Kasakura, K.; Kawakami, Y.; Jacquet, A.; Kawakami, T. Histamine-Releasing Factor Is a Novel Alarmin Induced by House Dust Mite Allergen, Cytokines, and Cell Death. J. Immunol. 2022, 209, 1851–1859. [Google Scholar] [CrossRef] [PubMed]
- Gour, N.; Lajoie, S.; Smole, U.; White, M.; Hu, D.; Goddard, P.; Huntsman, S.; Eng, C.; Mak, A.; Oh, S.; et al. Dysregulated invertebrate tropomyosin-dectin-1 interaction confers susceptibility to allergic diseases. Sci. Immunol. 2018, 3, 9841. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.M.; Gour, N.; Sharma, D.; Khalil, S.M.; Lane, A.P.; Lajoie, S. Epigenetic regulation of epithelial dectin-1 through an IL-33-STAT3 axis in allergic disease. Allergy 2022, 77, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.; Xie, M.; Wang, C.; Qian, Y.; Huang, J.; Sun, Z.; Zhang, H.; Wang, Y. Lactobacillus Casei Zhang Alleviates Shrimp Tropomyosin-Induced Food Allergy by Switching Antibody Isotypes through the NF-κB-Dependent Immune Tolerance. Mol. Nutr. Food Res. 2020, 64, 1900496. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and Immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef] [PubMed]
- Roan, F.; Obata-Ninomiya, K.; Ziegler, S.F. Epithelial cell–derived cytokines: More than just signaling the alarm. J. Clin. Investig. 2019, 129, 1441. [Google Scholar] [CrossRef] [PubMed]
- Chantret, I.; Barbat, A.; Dussaulx, E.; Brattain, M.G.; Zweibaum, A. Epithelial Polarity, Villin Expression, and Enterocytic Differentiation of Cultured Human Colon Carcinoma Cells: A Survey of Twenty Cell Lines. Cancer Res. 1988, 48, 1936–1942. [Google Scholar] [PubMed]
- Navabi, N.; McGuckin, M.A.; Lindén, S.K. Gastrointestinal Cell Lines Form Polarized Epithelia with an Adherent Mucus Layer when Cultured in Semi-Wet Interfaces with Mechanical Stimulation. PLoS ONE 2013, 8, e68761. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Roan, F.; Ziegler, S.F. The atopic march: Current insights into skin barrier dysfunction and epithelial cell-derived cytokines. Immunol. Rev. 2017, 278, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Gilliet, M.; Soumelis, V.; Watanabe, N.; Hanabuchi, S.; Antonenko, S.; De Waal-Malefyt, R.; Liu, Y.J. Human dendritic cells activated by TSLP and CD40L induce proallergic cytotoxic T cells. J. Exp. Med. 2003, 197, 1059–1063. [Google Scholar] [CrossRef] [PubMed]
- Hoppenbrouwers, T.; Fogliano, V.; Garssen, J.; Pellegrini, N.; Willemsen, L.E.M.; Wichers, H.J. Specific Polyunsaturated Fatty Acids Can Modulate in vitro Human moDC2s and Subsequent Th2 Cytokine Release. Front. Immunol. 2020, 11, 529229. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, L.K.; Hummelshoj, L. Triggers of IgE class switching and allergy development. Ann. Med. 2007, 39, 440–456. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, M.; Ma, W.; Jin, S.; Song, W.; He, S. Cockroach Allergen Bla g 7 Promotes TIM4 Expression in Dendritic Cells Leading to Th2 Polarization. Mediat. Inflamm. 2013, 2013, 983149. [Google Scholar] [CrossRef] [PubMed]
- Newberry, R.D.; Hogan, S.P. Intestinal epithelial cells in tolerance and allergy to dietary antigens. J. Allergy Clin. Immunol. 2021, 147, 45–48. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Xu, L.; Liang, X.; Liu, X.; Zhao, Y.; Ma, C.; Gao, L. Tim-4 in Health and Disease: Friend or Foe? Front. Immunol. 2020, 11, 509660. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, K.; Suma, Y.; Ishizaki, S.; Nagashima, Y.; Shiomi, K. Molecular Cloning of Tropomyosins Identified as Allergens in Six Species of Crustaceans. 2007. Available online: https://pubs.acs.org/sharingguidelines (accessed on 3 July 2023).
- Remillard, J.F.; Gould, M.C.; Roslansky, P.F.; Novitsky, T.J. Quantitation of endotoxin in products using the LAL kinetic turbidimetric assay. Prog. Clin. Biol. Res. 1987, 231, 197–210. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Pairs | Reference | Cycling Conditions |
|---|---|---|---|
| Gapdh | fw 5′ GAAGGTGAAGGTCGGAGTCAA 3′ rv 5′ ACGTACTCAGCGCCAGCATC 3′ | [22] | Pre-incubation 2 min 50 °C Incubation 10 min 95 °C 40 cycles |
| Il33 | fw 5′ GAGCTAAGGCCACTGAGGAA 3′ rv 5′ TGGGCCTTTGAAGTTCCATA 3′ | [23] | |
| Il25 | fw 5′ CCAGGTGGTTGCATTCTTGG 3′ rv 5′ TGGCTGTAGGTGTGGGTTCC 3′ | [24] | |
| Tslp | fw 5′ CTCTGGAGCATCAGGGAGAC 3′ rv 5′ CAATTCCACCCCAGTTTCAC 3′ | [22] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuurveld, M.; Ogrodowczyk, A.M.; Benedé, S.; Czolk, R.; Lucia Bavaro, S.; Randow, S.; Markiewicz, L.H.; Wróblewska, B.; Molina, E.; Kuehn, A.; et al. Allergenic Shrimp Tropomyosin Distinguishes from a Non-Allergenic Chicken Homolog by Pronounced Intestinal Barrier Disruption and Downstream Th2 Responses in Epithelial and Dendritic Cell (Co)Culture. Nutrients 2024, 16, 1192. https://doi.org/10.3390/nu16081192
Zuurveld M, Ogrodowczyk AM, Benedé S, Czolk R, Lucia Bavaro S, Randow S, Markiewicz LH, Wróblewska B, Molina E, Kuehn A, et al. Allergenic Shrimp Tropomyosin Distinguishes from a Non-Allergenic Chicken Homolog by Pronounced Intestinal Barrier Disruption and Downstream Th2 Responses in Epithelial and Dendritic Cell (Co)Culture. Nutrients. 2024; 16(8):1192. https://doi.org/10.3390/nu16081192
Chicago/Turabian StyleZuurveld, Marit, Anna M. Ogrodowczyk, Sara Benedé, Rebecca Czolk, Simona Lucia Bavaro, Stefanie Randow, Lidia H. Markiewicz, Barbara Wróblewska, Elena Molina, Annette Kuehn, and et al. 2024. "Allergenic Shrimp Tropomyosin Distinguishes from a Non-Allergenic Chicken Homolog by Pronounced Intestinal Barrier Disruption and Downstream Th2 Responses in Epithelial and Dendritic Cell (Co)Culture" Nutrients 16, no. 8: 1192. https://doi.org/10.3390/nu16081192
APA StyleZuurveld, M., Ogrodowczyk, A. M., Benedé, S., Czolk, R., Lucia Bavaro, S., Randow, S., Markiewicz, L. H., Wróblewska, B., Molina, E., Kuehn, A., Holzhauser, T., & Willemsen, L. E. M. (2024). Allergenic Shrimp Tropomyosin Distinguishes from a Non-Allergenic Chicken Homolog by Pronounced Intestinal Barrier Disruption and Downstream Th2 Responses in Epithelial and Dendritic Cell (Co)Culture. Nutrients, 16(8), 1192. https://doi.org/10.3390/nu16081192