Detailed Distribution of Lipids in Greenshell™ Mussel (Perna canaliculus)

Abstract
:1. Introduction
2. Experimental Section
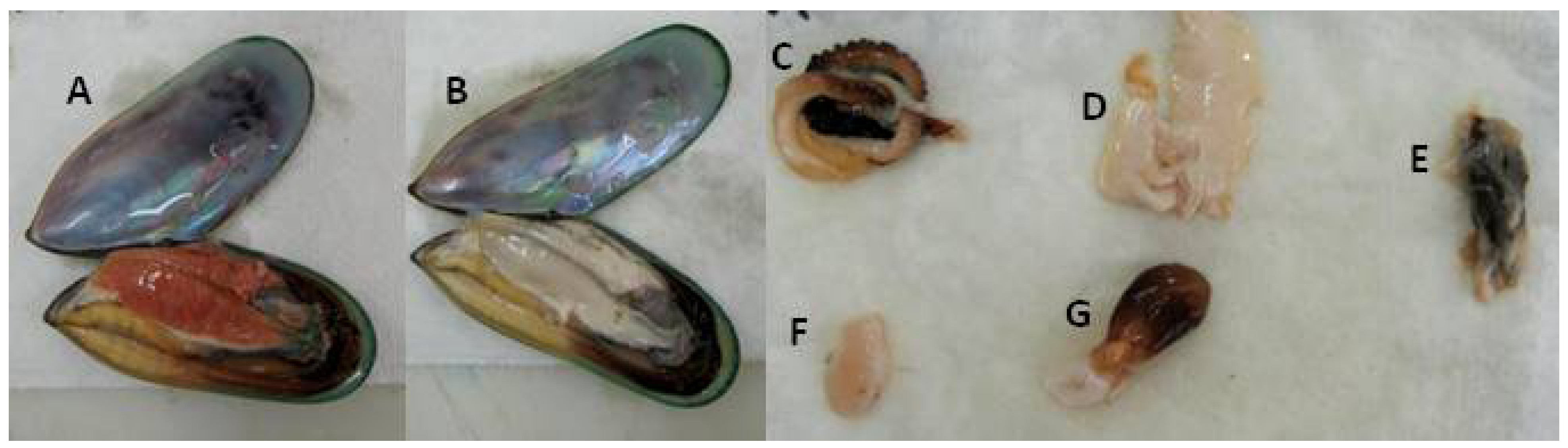
2.1. Sampling

2.2. Lipid Extraction, Fractionation, and Fatty Acid Analysis
2.3. Sterol Analysis
2.4. Statistical Analysis
3. Results
3.1. Lipid Content of Whole GSM
3.2. Lipid Classes of GSM

{kind=link}
{kind=link}
| Fatty acids (mg/g mussel ww) | Female | Male | F | p |
|---|---|---|---|---|
| 14:0 | 10.6 ± 2.1 | 7.3 ± 1.2 | 18.2 | <0.001 |
| 16:0 | 47.3 ± 6.9 | 42.2 ± 6.1 | ||
| 17:0 | 2.4 ± 0.4 | 2.6 ± 0.4 | ||
| 18:0 | 11.7 ± 1.8 | 12.1 ± 2.0 | ||
| Other SFA | 5.6 ± 1.1 | 5.4 ± 0.7 | ||
| Total SFA | 77.7 ± 12.0 | 69.5 ± 9.6 | ||
| 16:1n-7 | 18.3 ± 3.1 | 12.0 ± 2.0 | 26.5 | <0.001 |
| 18:1n-9 | 3.0 ± 0.6 | 2.8 ± 0.7 | ||
| 18:1n-7 | 7.6 ± 1.5 | 5.9 ± 1.0 | 7.7 | <0.05 |
| 20:1n-9 | 9.8 ± 1.5 | 10.0 ± 2.1 | ||
| 20:1n-7 | 4.3 ± 1.5 | 3.0 ± 0.6 | 5.9 | <0.05 |
| Other MUFA | 1.7 ± 0.5 | 0.9 ± 0.3 | 15.5 | <0.001 |
| Total MUFA | 44.7 ± 7.4 | 34.6 ± 5.1 | 11.4 | <0.01 |
| 18:4n-3 | 5.7 ± 1.0 | 3.2 ± 0.7 | 37.8 | <0.001 |
| 18:2n-6 | 3.7 ± 0.7 | 3.1 ± 0.6 | ||
| 18:3n-3 | 2.7 ± 0.3 | 2.0 ± 0.5 | 14.1 | <0.01 |
| 20:4n-6 | 4.6 ± 0.7 | 4.5 ± 1.0 | ||
| 20:5n-3 | 60.9 ± 11.4 | 42.5 ± 6.3 | 18.0 | <0.001 |
| 20:2 | 5.3 ± 0.7 | 5.8 ± 1.8 | ||
| 20:2NMI | 3.2 ± 0.5 | 2.7 ± 0.5 | ||
| 22:6n-3 | 59.2 ± 10.2 | 52.9 ± 8.1 | ||
| 22:5n-3 | 4.1 ± 0.7 | 3.2 ± 0.5 | 10.05 | <0.01 |
| 22:2NMI | 5.0 ± 0.8 | 4.7 ± 1.1 | ||
| Other PUFA | 7.7 ± 1.5 | 7.6 ± 2.1 | ||
| Total PUFA | 162.1 ± 25.4 | 132.2 ± 19.3 | 7.9 | <0.01 |
| Total n-3 | 132.8 ± 22.5 | 104.3 ± 14.5 | 10.2 | <0.01 |
| Total n-6 | 12.0 ± 1.7 | 11.1 ± 2.2 | ||
| Total other | 9.7 ± 6.4 | 10.1 ± 7.8 | ||
| Lipid Classes (g/100 g TLE) | ||||
| TAG | 19.4 ± 4.9 | 13.8 ± 3.2 | 5.8 | 0.03 |
| ST | 2.8 ± 1.3 | 2.9 ± 0.5 | ||
| FFA | tr | tr | ||
| PL | 77.8 ± 4.5 | 82.3 ± 3.2 | 6.0 | 0.03 |
| Lipid content | ||||
| g lipid /100 g wet weight | 1.9 ± 0.2 | 1.4 ± 0.3 | 14.7 | 0.001 |
| g lipid/100 g dry weight | 9.2 ± 1.1 | 7.4 ± 1.5 | 8.7 | 0.01 |
3.3. FA Profiles of GSM
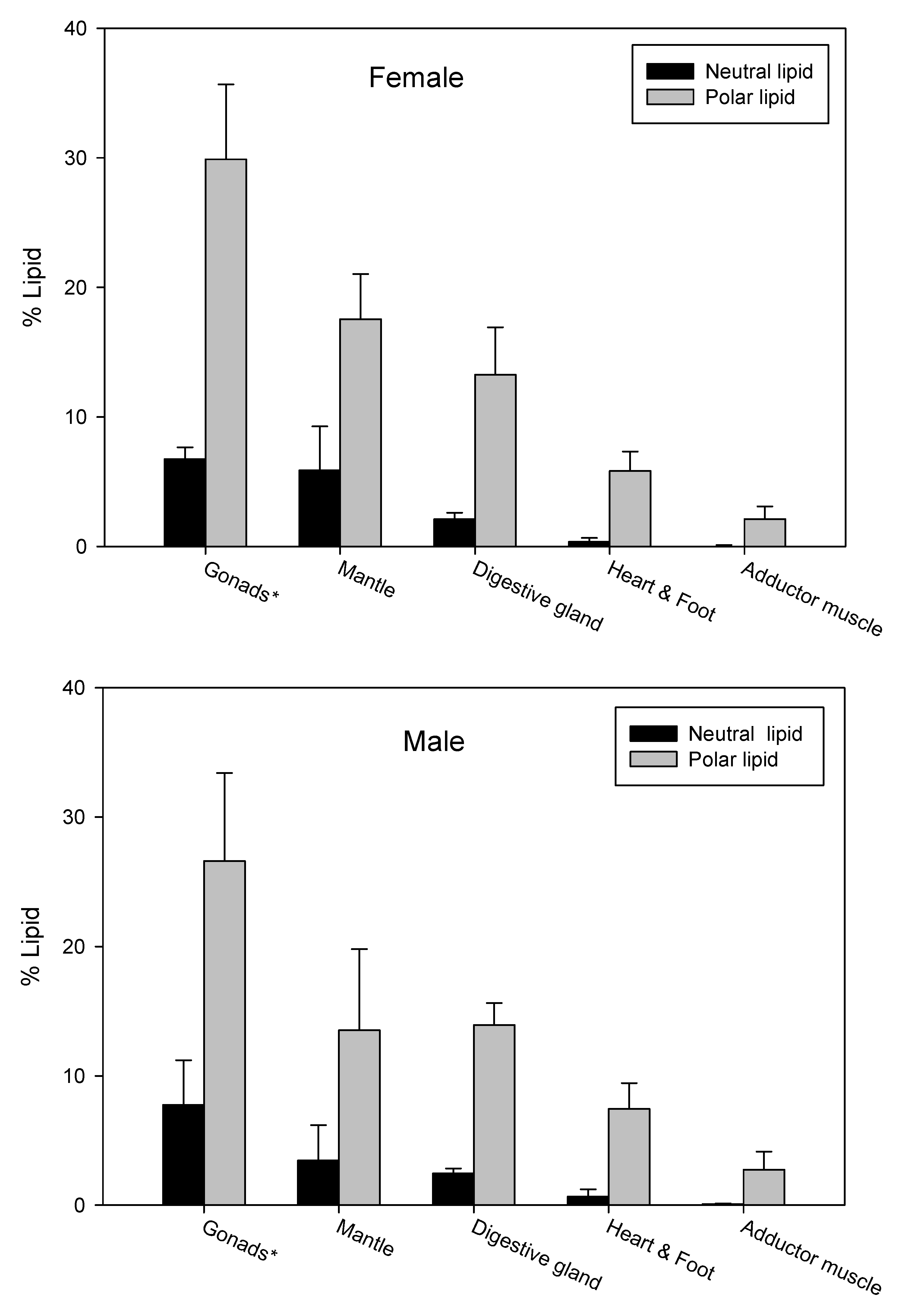
3.4. Lipid Content and Lipid Classes in Different Organs

3.5. FA Profiles of Lipids from Different Organs
3.6. Sterols
4. Discussion
4.1. Male vs. Female GSM
| Fatty acids (g/100 g TLE) | Mantle | Gonad | Digestive gland | f | p | |||||||||||||||
| Female | Male | Female | Male | Female | Male | |||||||||||||||
| 14:0 | 2.8 ± 0.2 | 3.4 ± 2.2 | 3.5 ± 0.3 | 2.9 ± 0.4 | 3.4 ± 0.7 | 3.0 ± 0.4 | ||||||||||||||
| 16:0 | 15.5 ± 0.4 | 17.1 ± 4.3 | 16.5 ± 1.1 | 16.8 ± 1.5 | 17.1 ± 3.8 | 15.6 ± 1.3 | ||||||||||||||
| 18:0 | 4.3 ± 0.4 | 4.5 ± 0.4 | 4.2 ± 0.3 | 5.2 ± 1.5 | 5.1 ± 1.6 | 4.8 ± 0.4 | ||||||||||||||
| Other SFA | 1.8 ± 0.2 ab | 1.8 ± 0.1 ab | 1.7 ± 0.2 a | 1.7 ± 0.1 a | 1.7 ± 0.2 ab | 2.4 ± 0.9 b | 2.9 | 0.03 | ||||||||||||
| Total SFA | 25.3 ± 0.4 | 27.7 ± 6.6 | 26.6 ± 1.4 | 27.5 ± 2.6 | 28.1 ± 6.3 | 26.5 ± 1.3 | ||||||||||||||
| 16:1n-7 | 5.2 ± 0.6 ab | 5.3 ± 1.8 ab | 6.3 ± 0.4 ab | 4.7 ± 1.6 a | 6.8 ± 0.6 b | 6.2 ± 0.8 ab | 3.2 | 0.02 | ||||||||||||
| 18:1n-9 | 1.3 ± 0.5 ab | 0.9 ± 0.2 a | 1.1 ± 0.3 ab | 1.0 ± 0.2 ab | 1.7 ± 0.6 b | 1.4 ± 0.3 ab | 3.0 | 0.03 | ||||||||||||
| 18:1n-7 | 2.2 ± 0.3 ab | 2.0 ± 0.2 a | 2.7 ± 0.2 c | 2.6 ± 0.4 ab | 2.5 ± 0.2 bc | 2.4 ± 0.2 abc | 6.2 | <0.001 | ||||||||||||
| 20:1n-9 | 4.2 ± 0.4 | 4.2 ± 0.7 | 3.5 ± 0.3 | 3.5 ± 0.6 | 3.8 ± 0.5 | 3.7 ± 0.3 | ||||||||||||||
| 20:1n-7 | 1.3 ± 0.1 | 1.0 ± 0.4 | 1.3 ± 0.1 | 1.2 ± 0.3 | 1.5 ± 0.4 | 1.2 ± 0.2 | ||||||||||||||
| Other MUFA | 0.5 ± 0.2 ab | 1.9 ± 1.3 b | 0.5 ± 0.1 ab | 0.3 ± 0.1 a | 0.9 ± 0.8 ab | 1.6 ± 1.6 ab | 3.1 | 0.02 | ||||||||||||
| Total MUFA | 14.6 ± 0.9 ab | 15.3 ± 1.0 bc | 15.3 ± 0.8 bc | 13.3 ± 1.2 a | 17.1 ± 1.3 c | 16.4 ± 1.2 bc | 9.4 | <0.001 | ||||||||||||
| 18:4n-3 | 1.2 ± 0.4 ab | 1.0 ± 0.3 a | 2.0 ± 0.4 bc | 1.4 ± 0.6 abc | 2.1 ± 0.6 c | 2.1 ± 0.4 c | 6.4 | <0.001 | ||||||||||||
| 18:2n-6 | 1.0 ± 0.2 a | 1.0 ± 0.1 a | 1.3 ± 0.3 ab | 1.1 ± 0.2 a | 1.5 ± 0.3 b | 1.3 ± 0.2 ab | 4.5 | 0.01 | ||||||||||||
| 18:3n-3 | 0.6 ± 0.2 a | 0.7 ± 0.2 ab | 1.0 ± 0.2 ab | 0.8 ± 0.3 ab | 1.0 ± 0.2 b | 1.0 ± 0.1 b | 4.4 | 0.01 | ||||||||||||
| 20:4n-6 | 1.9 ± 0.2 b | 1.9 ± 0.5 b | 1.3 ± 0.1 a | 1.2 ± 0.1 a | 1.3 ± 0.2 a | 1.3 ± 0.2 a | 9.3 | <0.001 | ||||||||||||
| 20:5n-3 | 18.9 ± 1.5 | 17.9 ± 1.5 | 22.0 ± 1.5 | 19.5 ± 3.7 | 18.1 ± 4.0 | 17.6 ± 3.0 | ||||||||||||||
| 20:2NMI | 2.9 ± 0.7 | 3.0 ± 0.9 | 1.6 ± 0.2 | 1.2 ± 0.2 | 1.9 ± 0.3 | 1.7 ± 0.2 | ||||||||||||||
| 20:4n-6 | 0.4 ± 0.1 | 0.3 ± 0.1 | 0.2 ± 0.1 | 0.2 ± 0.1 | 1.3 ± 0.2 | 1.3 ± 0.3 | ||||||||||||||
| 20:2n-6 | 1.5 ± 0.3 c | 1.4 ± 0.3 bc | 0.8 ± 0.1 a | 0.7 ± 0.1 a | 1.0 ± 0.1 ab | 0.9 ± 0.1 a | 14.8 | <0.001 | ||||||||||||
| 22:6n-3 | 22.0 ± 1.4 | 20.5 ± 4.0 | 19.2 ± 1.3 | 21.9 ± 2.4 | 17.9 ± 3.8 | 19.1 ± 1.5 | ||||||||||||||
| 22:5n-3 | 1.5 ± 0.3 | 1.3 ± 0.4 | 1.2 ± 0.1 | 2.0 ± 2.1 | 1.4 ± 0.3 | 1.1 ± 0.1 | ||||||||||||||
| 22:2NMI | 2.2 ± 0.4 b | 1.9 ± 1.0 ab | 1.2 ± 0.1 a | 1.2 ± 0.2 a | 1.7 ± 0.3 ab | 1.6 ± 0.3 ab | 4.4 | 0.01 | ||||||||||||
| Other PUFA | 1.8 ± 0.3 ab | 1.8 ± 0.5a | 2.9 ± 2.0 ab | 2.6 ± 1.3 ab | 3.3 ± 1.4 ab | 4.4 ± 2.2 b | 2.7 | 0.04 | ||||||||||||
| Total PUFA | 56.0 ± 2.3 | 52.8 ± 7.1 | 54.0 ± 1.8 | 54.5 ± 2.5 | 51.6 ± 7.1 | 52.5 ± 2.5 | ||||||||||||||
| Total n-3 | 44.2 ± 1.6 | 41.5 ± 4.5 | 45.4 ± 1.9 | 45.7 ± 4.1 | 40.7 ± 8.1 | 41.2 ± 4.0 | ||||||||||||||
| Total n-6 | 5.5 ± 0.5 | 5.2 ± 1.0 | 4.8 ± 0.8 | 4.9 ± 0.9 | 6.0 ± 1.1 | 6.7 ± 1.8 | ||||||||||||||
| Total other | 3.9 ± 1.8 | 3.8 ± 1.4 | 2.9 ± 2.1 | 4.4 ± 1.5 | 2.8 ± 1.1 | 4.2 ± 1.7 | ||||||||||||||
| Lipids class (g/100 g) | ||||||||||||||||||||
| TAG | 22.9 ± 8.0 ab | 18.4 ± 6.0 a | 33.7 ± 4.3 ab | 38.5 ± 17.6 b | 33.5 ± 10.8 ab | 28.8 ± 8.1 ab | 3.3 | 0.02 | ||||||||||||
| ST | 3.5 ± 0.9 | 3.2 ± 0.4 | 1.2 ± 0.6 | 6.1 ± 11.0 | 1.0 ± 1.1 | 1.7 ± 0.6 | ||||||||||||||
| PL | 73.5 ± 7.4 b | 77.3 ± 6.4 b | 64.5 ± 4.0 ab | 54.6 ± 12.7 a | 65.1 ± 10.6 ab | 68.7 ± 7.6 ab | 5.1 | 0.002 | ||||||||||||
| Lipid content | ||||||||||||||||||||
| g lipid /100 g wet weight | 2.0 ± 0.4 a | 1.5 ± 0.7 a | 4.0 ± 0.6 b | 3.1 ± 0.6 b | 3.7 ± 0.3 b | 3.1 ± 0.3 b | 19.8 | <0.001 | ||||||||||||
| g lipid/100 g dry weight | 11.8 ± 1.8 b | 7.9 ± 3.6 a | 13.3 ± 1.4 b | 11.7 ± 2.0 b | 13.2 ± 1.0 b | 11.9 ± 1.1 b | 5.6 | <0.001 | ||||||||||||
| Sterols g/100 g total sterols | Sterol name | Female | Male | f | p |
|---|---|---|---|---|---|
| 24-nordehydrocholesterol | 24-nordecholesta-5,22E-dien-3β-ol | 6.3 ± 0.9 | 6.7 ± 0.4 | ||
| 24-nordehydrocholestanol | 24-nor-5α-cholest-22E-en-3β-ol | 0.1 ± 0.0 | 0.2 ± 0.0 | ||
| Occelasterol | 27-nor-24-methylcholest-5,22E-dien-3β-ol | 4.3 ± 0.3 | 4.5 ± 0.3 | ||
| Trans-22-dehydrocholesterol | Cholesta-5,22E-dien-3β-ol | 12.4 ± 0.7 | 13.1 ± 0.8 | ||
| Trans-22-dehydrocholestanol | 5α-cholesta-22E-en-3β-ol | 0.3 ± 0.1 | 0.2 ± 0.1 | ||
| Cholesterol | Cholest-5-en-3β-ol | 28.5 ± 1.5 | 30.2 ± 0.8 | 11.1 | 0.004 |
| Cholestanol | 5α-cholestan-3β-ol | 0.3 ± 0.2 | 0.5 ± 0.4 | ||
| Brassicasterol | 24-methylcholesta-5,22E-diene-3β-ol | 23.4 ± 0.9 | 21.0 ± 1.4 | 21.3 | <0.001 |
| Brassicastanol | 24-methyl-5α-cholest-22E-en-3β-ol | 0.3 ± 0.3 | 0.3 ± 0.2 | ||
| Ergosterol | 24-methylcholesta-5,7,22E-triene-3β-ol | 0.6 ± 0.4 | 0.0 ± 0.1 | 16.6 | <0.001 |
| 24-Methylenecholesterol | 24-methylcholesta-5,24(28)-en-3β-ol | 16.2 ± 1.2 | 16.6 ± 1.6 | ||
| 24-Methylcholesterol | 4-methyl-5α-cholestan-3β-ol | 1.9 ± 0.4 | 1.6 ± 0.7 | ||
| Stigmastanol/Porifasterol | 24-ethyl-5α-cholesta-5,22E-diene-3β-ol | 0.6 ± 0.2 | 0.7 ± 0.2 | ||
| Sitosterol | 24-ethyl-5α-cholestan-3β-ol | 1.8 ± 0.4 | 2.1 ± 0.3 | ||
| Isofucosterol | 24-ethylcholesta-5,24(28)Z-dien-3β-ol | 1.7 ± 0.4 | 1.7 ± 0.4 | ||
| Unknown sterols | 1.3 ± 0.1 | 0.3 ± 0.1 | 40.8 | <0.001 |
4.2. Differences among Organs
4.3. Sterols
4.4. Implications of Results for the GSM Industry
5. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| ANOVA | 1-way analysis of variance |
| ALA | alpha-linolenic acid |
| BSTFA | N,O-bis(trimethylsilyl)trifluoroacetamide |
| CI | Condition index |
| DHA | docosahexaenoic acid |
| DPA | docosapentaenoic acid |
| EPA | eicosapentaenoic acid |
| FA | fatty acid(s) |
| FFA | free fatty acid(s) |
| FALD | dimethylacetals of aliphatic aldehydes |
| FAME | fatty acid(s) methyl ester |
| GC-MS | gas chromatography mass spectrometry |
| GSM | Greenshell™ mussel |
| HSD | honestly significant difference |
| LC | long chain (C ≥ 20) |
| MUFA | monounsaturated fatty acid(s) |
| NMI | non-methylene interrupted |
| TSN | non-saponifiable neutral |
| n-3 | Omega 3 |
| n-6 | Omega 6 |
| PUFA | polyunsaturated fatty acid(s) |
| SDA | stearidonic acid |
| SFA | saturated fatty acid(s) |
| TAG | triacylglycerol |
| TLE | total lipid extract |
| TMTD | trimethyl tetradecanoic acid |
| tr | trace amounts |
Supplementary Information
| Fatty acid (mg/g ww) | Female | Male | Female | Male | F | p |
|---|---|---|---|---|---|---|
| Polar lipid | Polar lipid | Neutral lipid | Neutral lipid | |||
| 14:0 | 5.3 ± 0.9 c | 3.5 ± 1.1 b | 1.9 ± 0.9 a | 0.9 ± 0.5 a | 50.8 | <0.001 |
| 16:0 | 23.2 ± 2.8 b | 19.8 ± 4.3 b | 5.9 ± 2.7 a | 3.0a ± 1.6 a | 110.8 | <0.001 |
| 18:0 | 5.6 ± 0.8 b | 5.8 ± 1.3 b | 1.0 ± 0.5 a | 0.7 ± 0.3 a | 113.0 | <0.001 |
| Other SFA | 2.6 ± 0.4 b | 2.5 ± 0.7 b | 0.5 ± 0.3 a | 0.3 ± 0.2a | 84.8 | <0.001 |
| Total SFA | 37.9 ± 4.5 b | 32.8 ± 7.4 b | 9.5 ± 4.4 a | 5.2 ± 2.7 a | 105.7 | <0.001 |
| 16:1n-7 | 9.2 ± 1.6 b | 5.7 ± 1.8 b | 3.3 ± 1.7 a | 1.6 ± 1.0 a | 45.5 | <0.001 |
| 18:1n-9 | 1.5 ± 0.4 b | 1.3 ± 0.3 b | 0.6 ± 0.3 a | 0.4 ± 0.2 a | 26.6 | <0.001 |
| 18:1n-7 | 3.7 ± 0.5 b | 2.9 ± 0.9b | 1.1 ± 0.6 a | 0.6 ± 0.3 a | 59.5 | <0.001 |
| 20:1n-9 | 4.7 ± 0.7 b | 4.6 ± 0.8 b | 0.7 ± 0.3 | 0.6a ± 0.2a | 223.3 | <0.001 |
| 20:1n-7 | 2.0 ± 0.6 c | 1.4 ± 0.4 b | 0.5 ± 0.2 a | 0.3 ± 0.1 a | 28.5 | <0.001 |
| Other MUFA | 0.8 ± 0.2 c | 0.4 ± 0.2 b | 0.3 ± 0.1 ab | 0.1 ± 0.1 a | 27.6 | <0.001 |
| Total MUFA | 21.7 ± 2.8 c | 16.3 ± 4.0 b | 6.5 ± 2.9 a | 3.6 ± 1.8 a | 80.8 | <0.001 |
| 18:4n-3 | 2.8 ± 0.4 d | 1.5 ± 0.5 c | 1.1 ± 0.6 b | 0.5 ± 0.3a | 42.2 | <0.001 |
| 18:2n-6 | 1.8 ± 0.3 b | 1.5 ± 0.4 b | 0.5 ± 0.3 a | 0.3 ± 0.2 a | 48.2 | <0.001 |
| 18:3n-3 | 1.3 ± 0.3 c | 0.9 ± 0.2 b | 0.4 ± 0.3 a | 0.2 ± 0.1 a | 45.9 | <0.001 |
| 20:4n-6 | 2.2 ± 0.3 b | 2.1 ± 0.4 b | 0.2 ± 0.1 a | 0.2 ± 0.1 a | 201.5 | <0.001 |
| 20:5n-3 | 30.5 ± 5.2 c | 20.5 ± 5.7 b | 7.5 ± 3.2 a | 4.1 ± 1.9 a | 80.0 | <0.001 |
| 20:2n-6 | 2.6 ± 0.5 b | 2.6 ± 0.4 b | 0.3 ± 0.1 a | 0.2 ± 0.0 a | 172.2 | <0.001 |
| 20:2NMI | 1.5 ± 0.1 b | 1.3 ± 0.3 b | 0.4 ± 0.1 a | 0.2 ± 0.1 a | 146.7 | <0.001 |
| 22:5n-6 | 0.3 ± 0.1 b | 0.3 ± 0.1 b | 0.1 ± 0.1 a | 0.1 ± 0.0 a | 6.8 | <0.001 |
| 22:6n-3 | 28.7 ± 3.7 b | 24.8 ± 4.9 b | 6.3 ± 3.3 a | 3.9 ± 1.7 a | 121.9 | <0.001 |
| 22:5n-3 | 2.0 ± 0.2 c | 1.5 ± 0.4 b | 0.3 ± 0.2 a | 0.2 ± 0.1 a | 139.4 | <0.001 |
| 22:2NMI | 2.4 ± 0.4 b | 2.2 ± 0.4 b | 0.2 ± 0.1 a | 0.2 ± 0.0 a | 177.7 | <0.001 |
| Other PUFA | 4.3 ± 1.0 b | 4.1 ± 1.4 b | 1.3 ± 1.8a | 0.8 ± 0.6 a | 20.7 | <0.001 |
| Total PUFA | 79.5 ± 9.6 c | 62.7 ± 14.0 b | 17.8 ± 7.9 a | 10.4 ± 4.5 a | 112.7 | <0.001 |
| Total n-3 | 65.5 ± 8.6 c | 49.5 ± 11.6 b | 15.6 ± 6.9 a | 9.0 ± 3.9 a | 107.5 | <0.001 |
| Total n-6 | 5.8 ± 1.1 b | 5.5 ± 1.7 b | 1.1 ± 0.6 a | 0.8 ± 0.3 a | 62.73 | <0.001 |
| Total other | 4.3 ± 2.9 b | 3.9 ± 2.5 b | 0.2 ± 0.1 a | 0.2 ± 0.1 a | 11.2 | <0.001 |
| Fatty acids (g/100 g TLE) | Adductor Muscle | Heart & foot | ||
|---|---|---|---|---|
| Female | Male | Female | Male | |
| 14:0 | 1.9 ± 0.6 | 2.4 ± 0.5 | 2.6 ± 0.5 | 2.3 ± 0.7 |
| 16:0 | 18.6 ± 6.1 | 26.3 ± 10.1 | 19.0 ± 1.8 | 16.1 ± 4.1 |
| 17:0 | 1.4 ± 0.5 | 1.5 ± 0.4 | 1.5 ± 0.2 | 1.3 ± 0.2 |
| 18:0 | 5.6 ± 0.9 | 6.8 ± 2.7 | 5.2 ± 0.3 | 5.0 ± 0.4 |
| Other SFA | 1.7 ± 0.5 | 1.3 ± 0.5 | 2.6 ± 0.2 | 2.4 ± 0.5 |
| Total SFA | 29.2 ± 7.9 | 38.1 ± 13.4 | 30.8 ± 2.4 | 27.1 ± 5.3 |
| 16:1n-7 | 2.0 ± 1.2 | 2.2 ± 0.6 | 4.4 ± 0.7 | 4.0 ± 1.2 |
| 18:1n-9 | 0.7 ± 0.2 | 0.7 ± 0.3 | 0.8 ± 0.1 | 0.8 ± 0.2 |
| 18:1n-7 | 2.6 ± 0.8 | 2.1 ± 0.7 | 1.9 ± 0.2 | 1.9 ± 0.3 |
| 20:1n-9 | 4.3 ± 0.7 | 3.5 ± 0.9 | 4.7 ± 0.6 | 4.8 ± 0.3 |
| 20:1n-7 | 0.9 ± 0.1 | 0.6 ± 0.3 | 1.1 ± 0.1 | 1.2 ± 0.2 |
| Other MUFA | 0.4 ± 0.4 | 0.6 ± 1.0 | 0.4 ± 0.0 | 0.4 ± 0.1 |
| Total MUFA | 10.8 ± 1.1 | 9.8 ± 2.3 | 13.2 ± 0.7 | 13.2 ± 1.1 |
| 18:4n-3 | 0.8 ± 0.5 | 0.3 ± 0.4 | 0.7 ± 0.3 | 0.8 ± 0.2 |
| 18:2n-6 | 1.5 ± 0.5 | 1.1 ± 0.6 | 1.5 ± 0.2 | 1.5 ± 0.1 |
| 18:3n-3 | 0.8 ± 0.3 | 0.6 ± 0.4 | 0.8 ± 0.1 | 0.8 ± 0.1 |
| 20:4n-6 | 1.9 ± 0.3 | 1.2 ± 0.6 | 3.1 ± 0.4 | 3.2 ± 0.2 |
| 20:5n-3 | 13.5 ± 1.8 | 12.0 ± 1.0 | 11.3 ± 1.5 | 12.9 ± 3 |
| 20:2NMI | 1.0 ± 0.3 | 0.5 ± 0.4 | 1.8 ± 0.3 | 1.7 ± 0.2 |
| 20:4n-6 | 0.4 ± 0.2 | 0.2 ± 0.2 | 0.5 ± 0.1 | 0.6 ± 3.2 |
| 20:2n-6 | 0.1 ± 0.1 | 0.1 ± 0.1 | 0.2 ± 0.0 | 0.2 ± 0.1 |
| 22:6n-3 | 25.3 ± 6.4 | 23.7 ± 10.8 | 19.6 ± 3.0 | 21.7 ± 0.2 |
| 22:5n-3 | 3.2 ± 0.9 | 2.4 ± 1.8 | 1.7 ± 0.4 | 1.9 ± 0.1 |
| 22:3 | 0.5 ± 0.1 | 0.3 ± 0.3 | 0.8 ± 0.2 | 0.9 ± 0.4 |
| 22:2NMI | 2.1 ± 0.8 | 1.7 ± 1.5 | 2.3 ± 0.4 | 2.4 ± 0.2 |
| Other PUFA | 1.5 ± 0.5 | 1.0 ± 0.5 | 1.8 ± 0.2 | 2.2 ± 0.2 |
| Total PUFA | 55.9 ± 9.6 | 47.6 ± 13.0 | 50.5 ± 3.8 | 54.5 ± 6.6 |
| Total n-3 | 43.6 ± 8.0 | 39.1 ± 11.5 | 34.1 ± 3.5 | 38.1 ± 5.8 |
| Total n-6 | 4.9 ± 1.3 | 3.2 ± 1.3 | 6.5 ± 1.2 | 6.4 ± 0.3 |
| Total other | 4.0 ± 1.2 | 4.4 ± 2.8 | 5.2 ± 1.5 | 4.9 ± 1.9 |
| Lipids | ||||
| TAG | 1.9 ± 2.2 | 2.7 ± 1.7 | 4.9 ± 3.3 | 7.5 ± 5.8 |
| ST | 7.1 ± 1.6 | 4.7 ± 2.7 | 8.1 ± 1.8 | 6.9 ± 0.7 |
| PL | 87.8 ± 6.0 | 91.3 ± 2.5 | 86.8 ± 1.8 | 83.1 ± 7 |
| Lipid content | ||||
| g lipid /100 g wet weight | 0.8 ± 0.4 | 1.2 ± 0.6 | 1.1 ± 0.1 | 1.3 ± 0.2 |
| g lipid/100 g dry weight | 3.2 ± 1.7 | 4.6 ± 2.7 | 5.0 ± 0.7 | 5.8 ± 1 |
Conflicts of Interest
References
- Jenkins, R.J. Mussel Cultivation in the Marlborough Sounds, 2nd ed.; Wright & Carman Ltd.: Wellington, New Zealand, 1985. [Google Scholar]
- Aquacultue New Zealand. Available online: http://aquaculture.org.nz/industry/ (accessed on 26 Novemeber 2013).
- Whitehouse, M.W.; Macrides, T.A.; Kalafatis, N.; Betts, W.H.; Haynes, D.R.; Broadbent, J. Anti-inflammatory activity of a lipid fraction (lyprinol) from the NZ green-lipped mussel. Inflammopharmacology 1997, 5, 237–246. [Google Scholar] [CrossRef]
- Shiels, I.A.; Whitehouse, M.W. Lyprinol: Anti-inflammatory and uterine-relaxant activities in rats, with special reference to a model for dysmenorrhoea. Allerg. Immunol. 2000, 32, 279–283. [Google Scholar]
- Halpern, G.M. Anti-inflammatory effects of a stabilized lipid extract of Perna canaliculus (lyprinol). Allerg. Immunol. 2000, 32, 272–278. [Google Scholar]
- Gruenwald, J.; Graubaum, H.J.; Hansen, K.; Grube, B. Efficacy and tolerability of a combination of lyprinol (r) and high concentrations of EPA and DHA in inflammatory rheumatoid disorders. Adv. Ther. 2004, 21, 197–201. [Google Scholar] [CrossRef]
- Treschow, A.P.; Hodges, L.D.; Wright, P.F.A.; Wynne, P.M.; Kalafatis, N.; Macrides, T.A. Novel anti-inflammatory omega-3 PUFAs from the new zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B 2007, 147, 645–656. [Google Scholar] [CrossRef]
- Tenikoff, D.; Murphy, K.J.; Le, M.; Howe, P.R.; Howarth, G.S. Lyprinol (stabilised lipid extract of new zealand green-lipped mussel): A potential preventative treatment modality for inflammatory bowel disease. J. Gastroenterol. 2005, 40, 361–365. [Google Scholar] [CrossRef]
- McPhee, S.; Hodges, L.D.; Wright, P.F.A.; Wynne, P.M.; Kalafatis, N.; Harney, D.W.; Macrides, T.A. Anti-cyclooxygenase effects of lipid extracts from the New Zealand green-lipped mussel, Perna canaliculus. Comp. Biochem. Physiol. B 2007, 146, 346–356. [Google Scholar] [CrossRef]
- Halpern, G.M. Novel anti-inflammatory mechanism of action of Lyprinol (r) in the aia rat model. Prog. Nutr. 2008, 10, 146–152. [Google Scholar]
- Wood, L.G.; Hazlewood, L.C.; Foster, P.S.; Hansbro, P.M. Lyprinol reduces inflammation and improves lung function in a mouse model of allergic airways disease. Clin. Exp. Allergy 2010, 40, 1785–1793. [Google Scholar] [CrossRef]
- Pumpa, K.L.; Fallon, K.E.; Bensoussan, A.; Papalia, S. The effects of Lyprinol (r) on delayed onset muscle soreness and muscle damage in well trained athletes: A double-blind randomised controlled trial. Complement. Ther. Med. 2011, 19, 311–318. [Google Scholar] [CrossRef]
- Lee, C.-H.; Lum, J.H.-K.; Ng, C.K.-C.; McKay, J.; Butt, Y.K.-C.; Wong, M.-S.; Lo, S.C.-L. Pain controlling and cytokine-regulating effects of lyprinol, a lipid extract of Perna canaliculus, in a rat adjuvant-induced arthritis model. Evid.Based Complement. Altern. Med. 2009, 6, 239–245. [Google Scholar] [CrossRef]
- Miller, M.R.; Perry, N.; Burgess, E.; Marshall, S. Regiospecific analyses of triacylglycerols of hoki (Macruronus novaezelandiae) and Greenshell™ mussel (Perna canaliculus). J. Am. Oil Chem. Soc. 2011, 88, 509–515. [Google Scholar] [CrossRef]
- Wakimoto, T.; Kondo, H.; Nii, H.; Kimura, K.; Egami, Y.; Oka, Y.; Yoshida, M.; Kida, E.; Ye, Y.P.; Akahoshi, S.; et al. Furan fatty acid as an anti-inflammatory component from the green-lipped mussel Perna canaliculus. Proc. Natl. Acad. Sci. USA 2011, 108, 17533–17537. [Google Scholar] [CrossRef]
- Barnathan, G. Non-methylene-interrupted fatty acids from marine invertebrates: Occurrence, characterization and biological properties. Biochimie 2009, 91, 671–678. [Google Scholar] [CrossRef]
- Braverman, N.E.; Moser, A.B. Functions of plasmalogen lipids in health and disease. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1442–1452. [Google Scholar] [CrossRef]
- Ostlund, R.E. Phytosterols in human nutrition. Annu. Rev. Nutr. 2002, 22, 533–549. [Google Scholar] [CrossRef]
- Murphy, K.J.; Mooney, B.D.; Mann, N.J.; Nichols, P.D.; Sinclair, A.J. Lipid, FA, and sterol composition of New Zealand green lipped mussel (Perna canaliculus) and Tasmanian blue mussel (Mytilus edulis). Lipids 2002, 37, 587–595. [Google Scholar] [CrossRef]
- MacLean, C.H.; Newberry, S.J.; Mojica, W.A.; Khanna, P.; Issa, A.M.; Suttorp, M.J.; Lim, Y.W.; Traina, S.B.; Hilton, L.; Garland, R.; et al. Effects of omega-3 fatty acids on cancer risk-A systematic review. JAMA 2006, 295, 403–415. [Google Scholar] [CrossRef]
- Horrocks, L.A.; Yeo, Y.K. Health benefits of docosahexaenoic acid (DHA). Pharm. Res. 1999, 40, 211–225. [Google Scholar] [CrossRef]
- Nettleton, J.A.; Katz, R. ω3 long-chain polyunsaturated fatty acids in type 2 diabetes: A review. J. Am. Diet. Assoc. 2005, 105, 428–440. [Google Scholar] [CrossRef]
- Kris-Etherton, P.M.; Hecker, K.D.; Binkoski, A.E. Polyunsaturated fatty acids and cardiovascular health. Nutr. Rev. 2004, 62, 414–426. [Google Scholar] [CrossRef]
- Zeldis, J.; Robinson, K.; Ross, A.; Hayden, B. First observations of predation by New Zealand greenshell mussels (Perna canaliculus) on zooplankton. J. Exp. Mar. Biol. Ecol. 2004, 311, 287–299. [Google Scholar] [CrossRef]
- Safi, K.A.; Gibbs, M.M. Importance of different size classes of phytoplankton in Beatrix Bay, Marlborough Sounds, New Zealand, and the potential implications for the aquaculture of the mussel, Perna canaliculus. N. Z. J. Mar. Freshw. Res. 2003, 37, 267–272. [Google Scholar] [CrossRef]
- Taylor, A.G.; Savage, C. Fatty acid composition of New Zealand green-lipped mussels, Perna canaliculus: Implications for harvesting for n-3 extracts. Aquaculture 2006, 261, 430–439. [Google Scholar] [CrossRef]
- Wolyniak, C.J.; Brenna, J.T.; Murphy, K.J.; Sinclair, A.J. Gas chromatography-chemical ionization-mass spectrometric fatty acid analysis of a commercial supercritical carbon dioxide lipid extract from New Zealand green-lipped mussel (Perna canaliculus). Lipids 2005, 40, 355–360. [Google Scholar] [CrossRef]
- McLean, C.H.; Bulling, K.R. Differences in lipid profile of New Zealand marine species over four seasons. J. Food Lipids 2005, 12, 313–326. [Google Scholar] [CrossRef]
- Hickman, R.W.; Waite, R.P.; Illingworth, J.; Meredythyoung, J.L.; Payne, G. The relationship between farmed mussels, Perna canaliculus, and available food in Pelorus-Kenepuru Sound, New Zealand, 1983–1985. Aquaculture 1991, 99, 49–68. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.G. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Kates, M. Techniques of Lipidology. Isolation, Analysis and Identification of Lipids. In Laboratory Techniques in Biochemistry and Molecular Biology; Burdon, R.H., Knippenberg, P.H.V., Eds.; Elsevier: New York, NY, USA, 1986; Volume 3, p. 464. [Google Scholar]
- Jones, G.J.; Nichols, P.D.; Shaw, P.M. Analysis of Microbial Sterols and Hopanoids. In Chemical Methods in Prokaryotic Sytematics; Goodfellow, M., O’Donnell, A.G., Eds.; John Wiley and Sons Ltd.: New York, NY, USA, 1994; pp. 163–195. [Google Scholar]
- Joseph, J.D. Lipid composition of marine and estuarine invertebrates. Part II: Mollusca. Prog. Lipid Res. 1982, 21, 109–153. [Google Scholar] [CrossRef]
- Martinez-Pita, I.; Sanchez-Lazo, C.; Ruiz-Jarabo, I.; Herrera, M.; Miguel Mancera, J. Biochemical composition, lipid classes, fatty acids and sexual hormones in the mussel Mytilus galloprovincialis from cultivated populations in South Spain. Aquaculture 2012, 358, 274–283. [Google Scholar]
- Gosling, E. Bivalve Molluscs. Biology, Ecology and Culture; Blackwell Publishing: Oxford, UK, 2003; pp. 1–443. [Google Scholar]
- Both, A.; Parrish, C.C.; Penney, R.W.; Thompson, R.J. Lipid composition of Mytilus edulis reared on organic waste from a Gadus morhua aquaculture facility. Aquat. Liv. Resour. 2011, 24, 295–301. [Google Scholar] [CrossRef]
- Both, A.; Parrish, C.C.; Penney, R.W. Growth and biochemical composition of Mytilus edulis when reared on effluent from a cod, Gadus morhua, aquaculture facility. J. Shellfish Res. 2012, 31, 79–85. [Google Scholar] [CrossRef]
- Sprecher, H.; Chen, Q. Polyunsaturated fatty acid biosynthesis: A microsomal-peroxisomal process. Prostaglands Leukot. Essent. Fatty Acids 1999, 60, 317–321. [Google Scholar] [CrossRef]
- Lesa, G.M.; Palfreyman, M.; Hall, D.H.; Clandinin, M.T.; Rudolph, C.; Jorgensen, E.M.; Schiavo, G. Long chain polyunsaturated fatty acids are required for efficient neurotransmission in c-elegans. J. Cell Sci. 2003, 116, 4965–4975. [Google Scholar] [CrossRef]
- Nelson, M.M.; Leighton, D.L.; Phleger, C.F.; Nichols, P.D. Comparison of growth and lipid composition in the green abalone, Haliotis fulgens, provided specific macroalgal diets. Comp. Biochem. Physiol. B 2002, 131, 695–712. [Google Scholar] [CrossRef]
- Lu, F.S.H.; Nielsen, N.S.; Baron, C.P.; Jacobsen, C. Oxidative degradation and non-enzymatic browning due to the interaction between oxidised lipids and primary amine groups in different marine PL emulsions. Food Chem. 2012, 135, 2887–2896. [Google Scholar] [CrossRef]
- Bayne, B.L.; Thompson, R.J.; Widdows, J. Phyiology: 1 Marine Mussels and Their Ecology and Physiology; Cambridge Univeristy Press: Cambridge, UK, 1976. [Google Scholar]
- Kerr, R.G.; Baker, B.J. Marine sterols. Nat. Prod. Rep. 1991, 8, 465–497. [Google Scholar] [CrossRef]
- Ostlund, R.E. Phytosterols and cholesterol metabolism. Curr. Opin. Lipidol. 2004, 15, 37–41. [Google Scholar] [CrossRef]
- Kritchevsky, D.; Chen, S.C. Phytosterols-health benefits and potential concerns: A review. Nutr. Res. 2005, 25, 413–428. [Google Scholar] [CrossRef]
- Teshima, S.; Kanazawa, A. Biosynthesis of sterols in abalone, Haliotis gurneri, and mussel, Mytilus edulis. Compar. Biochem. Physiol. B Compar. Biochem. 1974, 47, 555–561. [Google Scholar] [CrossRef]
- Miller, M.R.; Nichols, P.D.; Carter, C.G. The digestibility and accumulation of dietary phytosterols in Atlantic salmon (Salmo salar l.) smolt fed diets with replacement plant oils. Lipids 2008, 43, 549–557. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Miller, M.R.; Pearce, L.; Bettjeman, B.I. Detailed Distribution of Lipids in Greenshell™ Mussel (Perna canaliculus). Nutrients 2014, 6, 1454-1474. https://doi.org/10.3390/nu6041454
Miller MR, Pearce L, Bettjeman BI. Detailed Distribution of Lipids in Greenshell™ Mussel (Perna canaliculus). Nutrients. 2014; 6(4):1454-1474. https://doi.org/10.3390/nu6041454
Chicago/Turabian StyleMiller, Matthew R., Luke Pearce, and Bodhi I. Bettjeman. 2014. "Detailed Distribution of Lipids in Greenshell™ Mussel (Perna canaliculus)" Nutrients 6, no. 4: 1454-1474. https://doi.org/10.3390/nu6041454
APA StyleMiller, M. R., Pearce, L., & Bettjeman, B. I. (2014). Detailed Distribution of Lipids in Greenshell™ Mussel (Perna canaliculus). Nutrients, 6(4), 1454-1474. https://doi.org/10.3390/nu6041454