Fat Mass and Obesity-Associated (FTO) Gene Polymorphisms Are Associated with Physical Activity, Food Intake, Eating Behaviors, Psychological Health, and Modeled Change in Body Mass Index in Overweight/Obese Caucasian Adults

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Study Participants
2.2. Genotyping
2.3. Anthropometry
2.4. Physical Activity
2.5. Dietary Intake
2.6. Eating Behavior

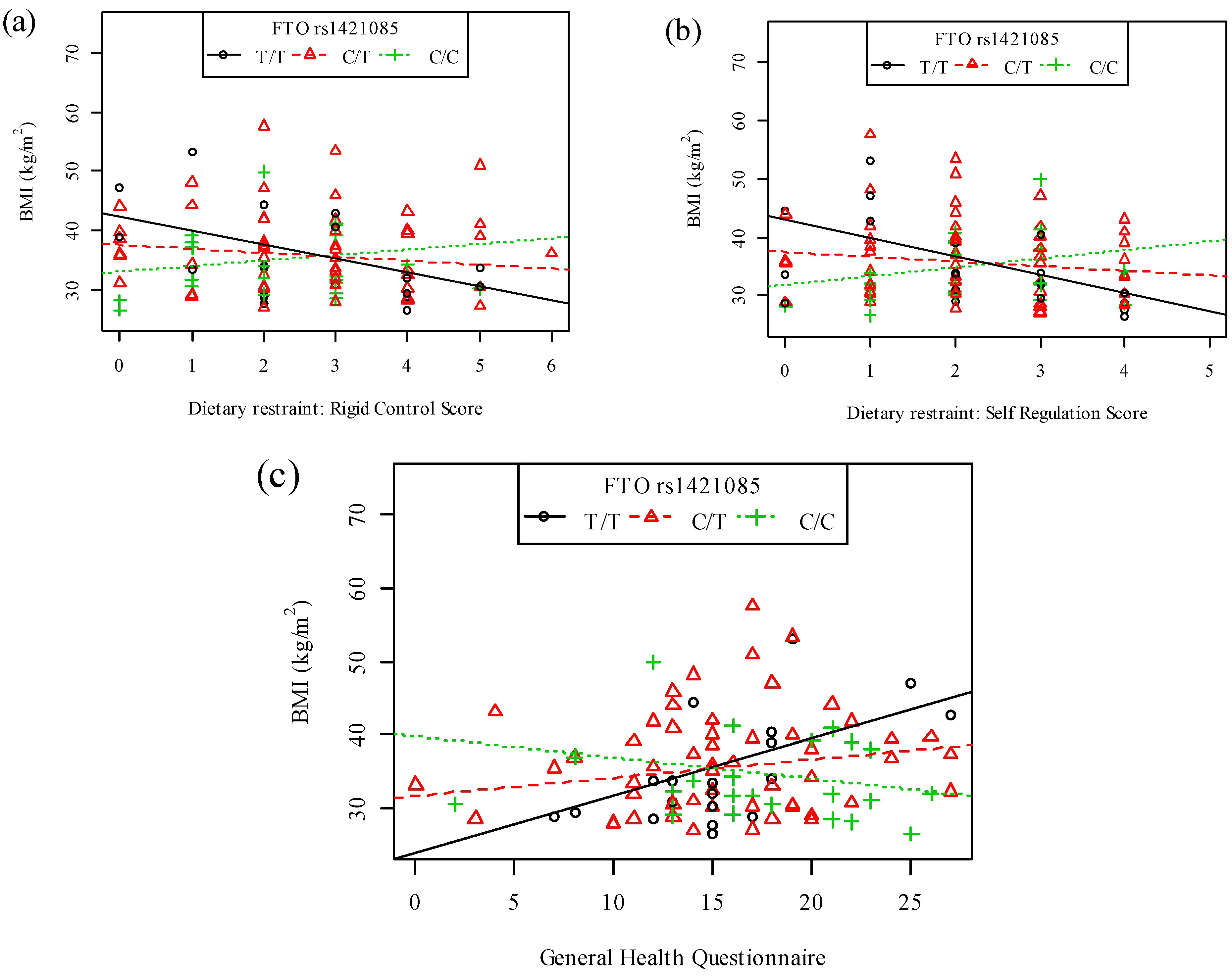{kind=link}
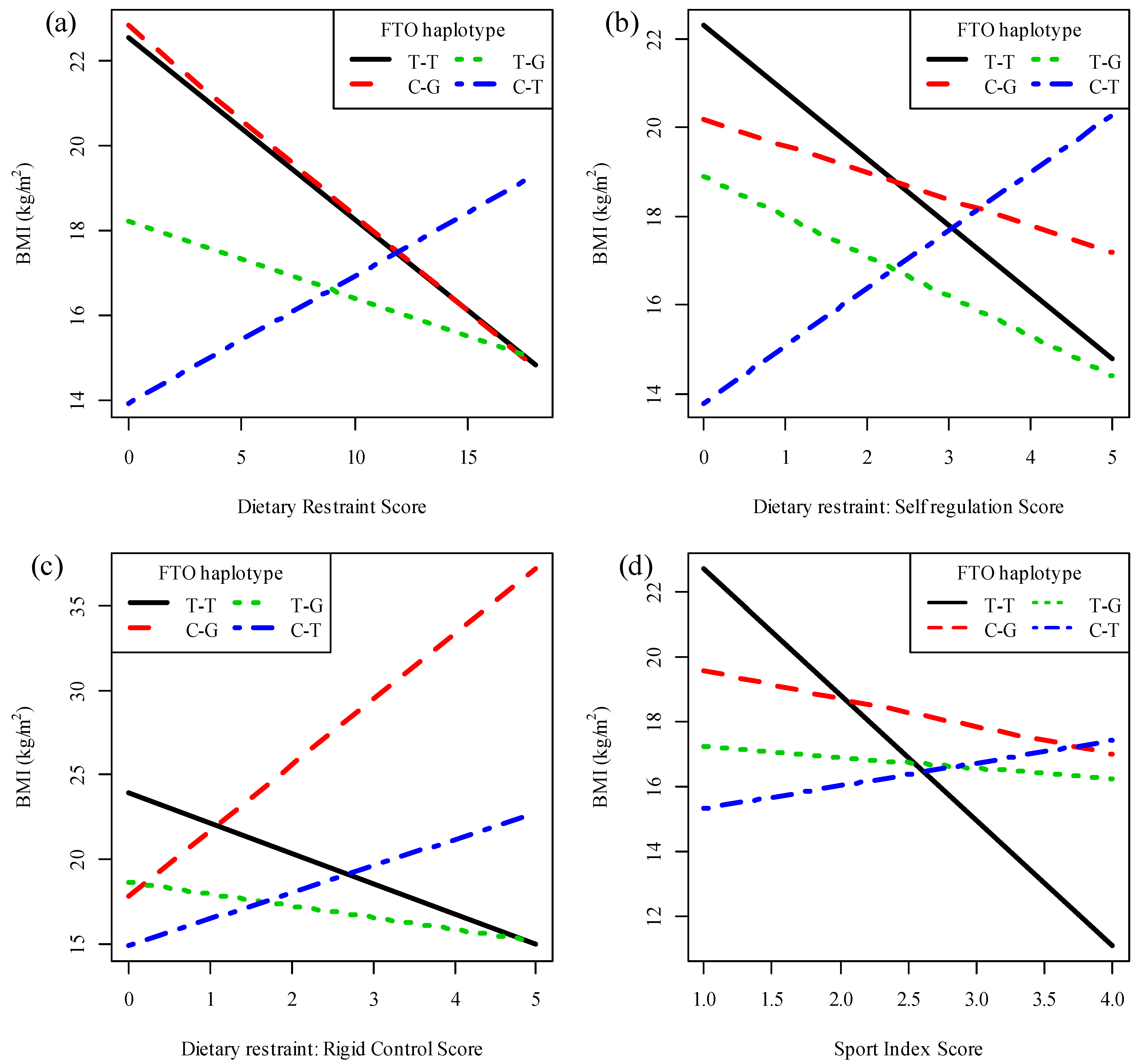{kind=link}
| Eating Behavior Scale or Subscale | Explanation of Scale or Subscale | Score Range | Reported/ Expected Association with BMI | Reference |
|---|---|---|---|---|
| Dietary restraint | The extent to which food intake is cognitively restricted (by thought and will power) in order to control body shape and weight. | 0 to 21 | − or + * | [48,49,50] |
| Flexible control (consistent restraint) | A more gradual approach towards eating and dieting. Foods like sweets and treats or fattening foods are eaten but in smaller quantities, without feelings of guilt. | 0 to 7 | − | [49] |
| Rigid control (inconsistent restraint) | A dichotomous (all-or-nothing) approach towards eating and dieting. All sweets, treats, and fattening foods are avoided. | 0 to 7 | + | [49] |
| Strategic dieting behavior | Specific behaviors employed to control weight | 0 to 4 | − or + § | [50] |
| Attitude to self-regulation (of eating) | Subjects’ general view on dietary intake and weight control. | 0 to 5 | + | [50] |
| Avoidance of fattening foods | Deliberate efforts to decrease fat content in the diet. | 0 to 4 | [50] | |
| Disinhibition | The extent of inability to control food intake in response to the presence of (1) palatable food that may result in the over-consumption of food or (2) stimuli such as emotional stress or social eating cues that may result in the inability to resist food intake when not hungry | 0 to 16 | + | [48,49,50] |
| Habitual susceptibility to disinhibition | Circumstances predispose to recurrent disinhibition | 0 to 5 | + | [50] |
| Emotional susceptibility to disinhibition | Specific negative affective states such as emotional stress result in disinhibition | 0 to 3 | + | [50] |
| Situational susceptibility to disinhibition | Specific environmental cues result in disinhibition | 0 to 5 | + | [50] |
| Perceived hunger | The extent of food intake in response to susceptibility to general subjective feelings and perceptions of hunger and the behavioral consequences thereof | 0 to 16 | + | [48,52] |
| Internal locus for hunger | Type of hunger that is interpreted and regulated internally | 0 to 6 | + | [50] |
| External locus for hunger | Type of hunger that is regulated by external cues | 0 to 6 | + | [50] |
2.7. Psychological Health
2.8. Statistical Analyses
3. Results
3.1. Socio-Demographic and Weight Profile of the Cross-Sectional Sample
| Socio-Demographic Variables and Polymorphisms | n | % | BMI (kg/m2) Mean ± SD | p-Value |
|---|---|---|---|---|
| Gender | ||||
| Female | 112 | 84.2 | 35.3 ± 6.9 | 0.818 |
| Male | 21 | 15.8 | 34.9 ± 4.5 | |
| Marital status | ||||
| Married/living together | 86 | 64.7 | 35.1 ± 6.3 | 0.703 |
| Unmarried (including separated/divorced) | 47 | 35.3 | 35.5 ± 7.1 | |
| Home language | ||||
| Afrikaans | 120 | 90.2 | 35.2 ± 6.5 | 0.729 |
| English | 13 | 9.8 | 35.8 ± 7.2 | |
| Level of education | ||||
| Completed Grade 10 or Matric | 43 | 32.3 | 35.1 ± 7.3 | 0.887 |
| Tertiary qualification | 90 | 67.7 | 35.3 ± 6.2 | |
| Living | ||||
| alone | 22 | 16.5 | 37.4 ± 8.0 | 0.156 |
| with friends/parents | 20 | 15.0 | 33.7 ± 6.0 | |
| with a partner | 30 | 22.6 | 33.8 ± 5.6 | |
| with a partner and child (ren) | 61 | 45.9 | 35.7 ± 6.5 | |
| FTO rs1421085 polymorphism | ||||
| Genotype frequencies: TT | 21 | 20.8 | 34.5 ± 7.3 | 0.1566 |
| TC | 55 | 54.5 | 36.6 ± 7.0 | |
| CC | 25 | 24.7 | 33.6 ± 5.4 | |
| Allele frequencies: Risk allele: C | 52.0 | 0.5831 | ||
| FTO rs17817449 polymorphism | ||||
| Genotype frequencies: TT | 31 | 29.5 | 35.9 ± 7.0 | 0.7814 |
| TG | 32 | 30.5 | 34.9 ± 6.8 | |
| GG | 42 | 40.0 | 36.0 ± 7.2 | |
| Allele frequencies: Risk allele: G | 55.2 | 0.9178 | ||
| FTO rs1421085-rs17817449 haplotype frequencies | ||||
| Haplotypes: C-G | * | 27.3 | * | 0.6010 |
| C-T | 24.7 | |||
| T-G | 27.9 | |||
| T-T | 20.0 |
3.2. Genotype and Allele Frequencies
3.3. Dietary Intake of Indicator Food Groups
| Lifestyle/Psychological Health Variable | Polymorphisms | Genotypes | n * | Mean ± SD | Effect Size ± SE | Model | p-Value |
|---|---|---|---|---|---|---|---|
| Food groups | Frequency of intake/day | ||||||
| High fat foods | FTO rs17817449 | T-allele carriers | 59 | 4.9 ± 3.6 | 1.74 ± 0.87 | Recessive G | 0.0494 |
| GG | 38 | 6.6 ± 4.9 | |||||
| Refined starches | FTO rs17817449 | T-allele carriers | 59 | 0.94 ± 1.36 | 0.67 ± 0.30 | Recessive G | 0.0287 |
| GG | 38 | 1.59 ± 1.55 | |||||
| TFEQ | Questionnaire score | ||||||
| Perceived hunger | FTO rs1421085 | TT | 20 | 5.9 ± 3.4 | 1.43 ± 0.52 | Additive C | 0.0072 |
| TC | 54 | 7.3 ± 3.0 | |||||
| CC | 23 | 8.8 ± 3.9 | |||||
| Hunger: Internal locus of control | FTO rs1421085 | TT | 20 | 2.2 ± 1.7 | 0.84 ± 0.28 | Additive C | 0.0038 |
| TC | 54 | 2.8 ± 1.8 | |||||
| CC | 23 | 3.9 ± 2.0 | |||||
| Emotional disinhibition | FTO rs1421085 | TT | 20 | 2.0 ± 1.3 | 0.58 ± 0.26 | Dominant C | 0.0281 |
| C-allele carriers | 77 | 2.4 ± 1.1 | |||||
| Restraint scale: | FTO rs1421085 | TT | 20 | 1.7 ± 1.6 | 0.89 ± 0.43 | Dominant C | 0.0392 |
| Flexible control | C-allele carriers | 77 | 2.5 ± 1.7 | ||||
| BDI | FTO rs17817449 | T-allele carriers | 61 | 12.7 ± 9.4 | 4.58 ± 2.02 | Recessive G | 0.0256 |
| GG | 40 | 17.6 ± 10.7 |
3.4. Eating Behavior

3.5. Physical Activity

3.6. Psychological Health
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization. Diet, Nutrition and the Prevention of Chronic Diseases. In Report for a Joint Who/Fao Expert Consultation; WHO Technical Report Series, No. 916; WHO: Geneva, Switzerland, 2003. [Google Scholar]
- Obesity: Situation and Trends. Available online: http://www.who.int/gho/ncd/risk_factors/obesity_text/en/index.html (accessed on 19 March 2014).
- Marti, A.; Martinez-González, M.A.; Martinez, J.A. Interaction between genes and lifestyle factors on obesity. Proc. Nutr. Soc. 2008, 67, 1–8. [Google Scholar] [CrossRef]
- Romao, I.; Roth, J. Genetic and environmental interactions in obesity and type 2 diabetes. J. Am. Diet. Assoc. 2008, 108, S24–S28. [Google Scholar] [CrossRef]
- Larder, R.; Cheung, M.K.; Tung, Y.C.; Yeo, G.S.; Coll, A.P. Where to go with FTO? Trends Endocrinol. Metab. 2011, 22, 53–59. [Google Scholar] [CrossRef]
- Speliotes, E.K.; Willer, C.J.; Berndt, S.I.; Monda, K.L.; Thorleifsson, G.; Jackson, A.U.; Lango Allen, H.; Lindgren, C.M.; Luan, J.; Mägi, R.; et al. Association analyses of 249,796 individuals reveal 18 new loci associated with body mass index. Nat. Genet. 2010, 42, 937–948. [Google Scholar] [CrossRef]
- Dina, C.; Meyre, D.; Gallina, S.; Durand, E.; Körner, A.; Jacobson, P.; Carlsson, L.M.; Kiess, W.; Vatin, V.; Lecoeur, C.; et al. Variation in FTO contributes to childhood obesity and severe adult obesity. Nat. Genet. 2007, 39, 724–726. [Google Scholar] [CrossRef]
- Frayling, T.M.; Timpson, N.J.; Weedon, M.N.; Zeggini, E.; Freathy, R.M.; Lindgren, C.M.; Perry, J.R.; Elliott, K.S.; Lango, H.; Rayner, N.W.; et al. A common variant in the FTO gene is associated with body mass index and predisposes to childhood and adult obesity. Science 2007, 316, 889–894. [Google Scholar] [CrossRef]
- Scuteri, A.; Sanna, S.; Chen, W.M.; Uda, M.; Albai, G.; Strait, J.; Najjar, S.; Nagaraja, R.; Orru, M.; Usala, G.; et al. Genome-wide association scan shows genetic variants in the FTO gene are associated with obesity-related traits. PLoS Genet. 2007, 3, e115. [Google Scholar] [CrossRef]
- Frayling, T.M.; Ong, K. Piecing together the FTO jigsaw. Genome Biol. 2011, 12, 104. [Google Scholar] [CrossRef]
- Gerken, T.; Girard, C.A.; Tung, Y.C.; Webby, C.J.; Saudek, V.; Hewitson, K.S.; Yeo, G.S.; McDonough, M.A.; Cunliffe, S.; McNeill, L.A.; et al. The obesity-associated FTO gene encodes a 2-oxoglutarate-dependent nucleic acid demethylase. Science 2007, 318, 1469–1472. [Google Scholar] [CrossRef]
- Sanchez-Pulido, L.; Andrade-Navarro, M.A. The FTO (fat mass and obesity associated) gene codes for a novel member of the non-heme dioxygenase superfamily. BMC Biochem. 2007, 8, 23. [Google Scholar] [CrossRef]
- Han, Z.; Niu, T.; Chang, J.; Lei, X.; Zhao, M.; Wang, Q.; Cheng, W.; Wang, J.; Feng, Y.; Chai, J. Crystal structure of the FTO protein reveals basis for its substrate specificity. Nature 2010, 464, 1205–1209. [Google Scholar] [CrossRef]
- Fawcett, K.A.; Barroso, I. The genetics of obesity: FTO leads the way. Trends Genet. 2010, 26, 266–274. [Google Scholar] [CrossRef]
- Jia, G.; Yang, C.; Yang, S.; Jian, X.; Yi, C.; Zhou, Z.; He, C. Oxidative demethylation of 3-methylthymine and 3-methyluracil in single-stranded DNA and RNA by mouse and human FTO. FEBS Lett. 2008, 582, 3313–3319. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, YG.; He, C. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Chem. Biol. 2011, 7, 885–887. [Google Scholar]
- Fredriksson, R.; Hägglund, M.; Olszewski, P.K.; Stephansson, O.; Jacobsson, J.A.; Olszewska, A.M.; Levine, A.S.; Lindblom, J.; Schiöth, H.B. The obesity gene, FTO, is of ancient origin, up-regulated during food deprivation and expressed in neurons of feeding-related nuclei of the brain. Endocrinology 2008, 149, 2062–2071. [Google Scholar] [CrossRef]
- Stratigopoulos, G.; Padilla, S.L.; LeDuc, C.A.; Watson, E.; Hattersley, A.T.; McCarthy, M.I.; Zeltser, L.M.; Chung, W.K.; Leibel, R.L. Regulation of Fto/Ftm gene expression in mice and humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 294, R1185–R1196. [Google Scholar] [CrossRef]
- Tung, Y.C.; Ayuso, E.; Shan, X.; Bosch, F.; O’Rahilly, S.; Coll, A.P.; Yeo, G.S. Hypothalamic-specific manipulation of Fto, the ortholog of the human obesity gene FTO, affects food intake in rats. PLoS One 2010, 5, e8771. [Google Scholar]
- Poritsanos, N.J.; Lew, P.S.; Fischer, J.; Mobbs, C.V.; Nagy, J.I.; Wong, D.; Rüther, U.; Mizuno, T.M. Impaired hypothalamic Fto expression in response to fasting and glucose in obese mice. Nutr. Diabetes 2011, 1, e19. [Google Scholar] [CrossRef]
- Wåhlén, K.; Sjölin, E.; Hoffstedt, J. The common rs9939609 gene variant of the fat mass- and obesity-associated gene FTO is related to fat cell lipolysis. J. Lipid Res. 2008, 49, 607–611. [Google Scholar] [CrossRef]
- Zabena, C.; González-Sánchez, J.L.; Martínez-Larrad, M.T.; Torres-García, A.; Alvarez-Fernández-Represa, J.; Corbatón-Anchuelo, A.; Pérez-Barba, M.; Serrano-Ríos, M. The FTO obesity gene. Genotyping and gene expression analysis in morbidly obese patients. Obes. Surg. 2009, 19, 87–95. [Google Scholar] [CrossRef]
- Fischer, J.; Koch, L.; Emmerling, C.; Vierkotten, J.; Peters, T.; Brüning, J.C.; Rüther, U. Inactivation of the Fto gene protects from obesity. Nature 2009, 458, 894–988. [Google Scholar] [CrossRef]
- Cheung, M.K.; Gulati, P.; O’Rahilly, S.; Yeo, G.S. FTO expression is regulated by availability of essential amino acids. Int. J. Obes. 2013, 37, 744–747. [Google Scholar] [CrossRef]
- Pitman, R.T.; Fong, J.T.; Billman, P.; Puri, N. Knockdown of the fat mass and obesity gene disrupts cellular energy balance in a cell-type specific manner. PLoS One 2012, 7, e38444. [Google Scholar]
- Church, C.; Moir, L.; McMurray, F.; Girard, C.; Banks, G.T.; Teboul, L.; Wells, S.; Brüning, J.C.; Nolan, P.M.; Ashcroft, F.M.; et al. Overexpression of Fto leads to increased food intake and results in obesity. Nat. Genet. 2010, 42, 1086–1092. [Google Scholar] [CrossRef]
- McMurray, F.; Church, C.D.; Larder, R.; Nicholson, G.; Wells, S.; Teboul, L.; Tung, Y.C.; Rimmington, D.; Bosch, F.; Jimenez, V.; et al. Adult onset global loss of the Fto gene alters body composition and metabolism in the mouse. PLoS Genet. 2013, 9, e1003166. [Google Scholar] [CrossRef]
- Gao, X.; Shin, Y.H.; Li, M.; Wang, F.; Tong, Q.; Zhang, P. The fat mass and obesity associated gene FTO functions in the brain to regulate postnatal growth in mice. PLoS One 2010, 5, e14005. [Google Scholar]
- Cecil, J.E.; Tavendale, R.; Watt, P.; Hetherington, M.M.; Palmer, C.N. An obesity-associated FTO gene variant and increased energy intake in children. N. Engl. J. Med. 2008, 359, 2558–2566. [Google Scholar] [CrossRef]
- Do, R.; Bailey, S.D.; Desbiens, K.; Belisle, A.; Montpetit, A.; Bouchard, C.; Pérusse, L.; Vohl, M.C.; Engert, J.C. Genetic variants of FTO influence adiposity, insulin sensitivity, leptin levels, and resting metabolic rate in the Quebec Family Study. Diabetes 2008, 57, 1147–1150. [Google Scholar] [CrossRef]
- Berentzen, T.; Kring, S.I.; Holst, C.; Zimmermann, E.; Jess, T.; Hansen, T.; Pedersen, O.; Toubro, S.; Astrup, A.; Sørensen, T.I. Lack of association of fatness-related FTO gene variants with energy expenditure or physical activity. J. Clin. Endocrinol. Metab. 2008, 93, 2904–2908. [Google Scholar] [CrossRef]
- Speakman, J.R.; Rance, K.A.; Johnstone, A.M. Polymorphisms of the FTO gene are associated with variation in energy intake, but not energy expenditure. Obesity 2008, 16, 1961–1965. [Google Scholar] [CrossRef]
- Hakanen, M.; Raitakari, O.T.; Lehtimäki, T.; Peltonen, N.; Pahkala, K.; Sillanmäki, L.; Lagström, H.; Viikari, J.; Simell, O.; Rönnemaa, T. FTO genotype is associated with body mass index after the age of seven years but not with energy intake or leisure-time physical activity. J. Clin. Endocrinol. Metab. 2009, 94, 1281–1287. [Google Scholar] [CrossRef]
- Haupt, A.; Thamer, C.; Staiger, H.; Tschritter, O.; Kirchhoff, K.; Machicao, F.; Häring, H.U.; Stefan, N.; Fritsche, A. Variation in the FTO gene influences food intake but not energy expenditure. Exp. Clin. Endocrinol. Diabetes 2009, 117, 194–197. [Google Scholar] [CrossRef]
- Tanofsky-Kraff, M.; Han, J.C.; Anandalingam, K.; Shomaker, L.B.; Columbo, K.M.; Wolkoff, L.E.; Kozlosky, M.; Elliott, C.; Ranzenhofer, L.M.; Roza, C.A.; et al. The FTO gene rs9939609 obesity-risk allele and loss of control over eating. Am. J. Clin. Nutr. 2009, 90, 1483–1488. [Google Scholar] [CrossRef]
- Wardle, J.; Llewellyn, C.; Sanderson, S.; Plomin, R. The FTO gene and measured food intake in children. Int. J. Obes. 2009, 33, 42–45. [Google Scholar] [CrossRef]
- McCaffery, J.M.; Papandonatos, G.D.; Peter, I.; Huggins, G.S.; Raynor, H.A.; Delahanty, L.M.; Cheskin, L.J.; Balasubramanyam, A.; Wagenknecht, L.E.; Wing, R.R. et al. Obesity susceptibility loci and dietary intake in the Look AHEAD Trial. Am. J. Clin. Nutr. 2012, 95, 1477–1486. [Google Scholar] [CrossRef]
- Stutzmann, F.; Cauchi, S.; Durand, E.; Calvacanti-Proença, C.; Pigeyre, M.; Hartikainen, A.L.; Sovio, U.; Tichet, J.; Marre, M.; Weill, J.; et al. Common genetic variation near MC4R is associated with eating behavior patterns in European populations. Int. J. Obes. 2009, 33, 373–378. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Norton, K.; Olds, T. Anthropometrica: A Textbook of Body Measurement for Sports and Health Courses; UNSW: Sydney, Australia, 1996; pp. 35–37. [Google Scholar]
- Albanes, D.; Conway, J.M.; Taylor, P.R.; Moe, P.W.; Judd, J. Validation and comparison of eight physical activity questionnaires. Epidemiology 1990, 1, 65–71. [Google Scholar] [CrossRef]
- Miller, D.J.; Freedson, P.S.; Kline, G.M. Comparison of activity levels using the Caltrac accelerometer and five questionnaires. Med. Sci. Sports Exerc. 1994, 26, 376–382. [Google Scholar]
- Richardson, M.T.; Ainsworth, B.E.; Wu, H.; Jacobs, D.R.; Leon, A.S. Ability of the atherosclerosis risk in communities (ARIC)/Baecke questionnaire to assess leisure-time physical activity. Int. J. Epidemiol. 1995, 24, 685–693. [Google Scholar] [CrossRef]
- Baecke, J.A.H.; Burema, J.; Frijters, J.E.R. A short questionnaire for the measurement of habitual physical activity in epidemiological studies. Am. J. Clin. Nutr. 1982, 36, 936–942. [Google Scholar]
- Steyn, N.P.; Senekal, M. A Guide for the Use of the Dietary Assessment and Education Kit (DAEK); Medical Research Council: Cape Town, South Africa, 2005. [Google Scholar]
- Laessle, R.G.; Tuschl, R.J.; Kotthaus, B.C.; Pirke, K.M. A comparison of the validity of three scales for the assessment of dietary restraint. J. Abnorm. Psychol. 1989, 98, 504–507. [Google Scholar] [CrossRef]
- Provencher, V.; Drapeau, V.; Tremblay, A.; Després, J.P.; Lemieux, S. Eating behaviors and indexes of body composition in men and women from the Québec Family Study. Obes. Res. 2003, 11, 783–792. [Google Scholar] [CrossRef]
- Stunkard, A.J.; Messick, S. The three-factor eating questionnaire to measure dietary restraint, disinhibition and hunger. J. Psychosom. Res. 1985, 29, 71–83. [Google Scholar] [CrossRef]
- Westenhoefer, J.; Stunkard, A.J.; Pudel, V. Validation of the flexible and rigid control dimensions of dietary restraint. Int. J. Eat. Disord. 1999, 26, 53–64. [Google Scholar] [CrossRef]
- Bond, M.J.; McDowell, A.J.; Wilkinson, J.Y. The measurement of dietary restraint, disinhibition and hunger: An examination of the factor structure of the Three Factor Eating Questionnaire (TFEQ). Int. J. Obes. 2001, 25, 900–906. [Google Scholar]
- Bas, M.; Donmez, S. Self-efficacy and restrained eating in relation to weight loss among overweight men and women in Turkey. Appetite 2009, 52, 209–216. [Google Scholar] [CrossRef]
- Chaput, J.P.; Leblanc, C.; Perusse, L.; Despres, J.P.; Bouchard, C.; Tremblay, A. Risk factors for adult overweight and obesity in the Quebec Family Study: Have we been barking up the wrong tree? Obesity 2009, 17, 1964–1970. [Google Scholar] [CrossRef]
- Beck, A.T.; Steer, R.A. Manual for the Beck Depression Inventory; The Psychological Corporation: San Antonio, TX, USA, 1987. [Google Scholar]
- Beck, A.T.; Steer, R.A.; Brown, G.K. BDI-II Manual; The Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Goldberg, D.P. The Detection of Psychiatric Illness by Questionnaire; Oxford University Press: London, UK, 1972. [Google Scholar]
- Goldberg, D.P.; Rickels, K.; Downing, R.; Hesbacher, P. A comparison of two psychiatric screening tests. Br. J. Psychiatry 1976, 129, 61–67. [Google Scholar] [CrossRef]
- Banks, M.H. Validation of the General Health Questionnaire in a young community sample. Psychol. Med. 1983, 13, 349–353. [Google Scholar] [CrossRef]
- Lorr, M.; Wunderlich, R.A. Two objective measures of self-esteem. J. Pers. Assess. 1986, 50, 18–23. [Google Scholar] [CrossRef]
- Reynolds, W.M. Measurement of academic self-concept in college students. J. Pers. Assess. 1988, 52, 223–240. [Google Scholar] [CrossRef]
- Rosenberg, M. Society and the Adolescent Self-Image; Princeton University Press: Princeton, NJ, USA, 1965. [Google Scholar]
- R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 18 July 2014).
- Andersson, U.; McKean-Cowdin, R.; Hjalmars, U.; Malmer, B. Genetic variants in association studies-review of strengths and weaknesses in study design and current knowledge of impact on cancer risk. Acta Oncol. 2009, 48, 948–954. [Google Scholar] [CrossRef]
- Nyholt, D.R. A simple correction for multiple testing for single-nucleotide polymorphisms in linkage disequilibrium with each other. Am. J. Hum. Genet. 2004, 74, 765–769. [Google Scholar] [CrossRef]
- Perneger, T.V. What’s wrong with Bonferroni adjustments. BMJ 1998, 316, 1236–1238. [Google Scholar] [CrossRef]
- Wardle, J.; Carnell, S.; Haworth, C.M.; Farooqi, I.S.; O’Rahilly, S.; Plomin, R. Obesity associated genetic variation in FTO is associated with diminished satiety. J. Clin. Endocrinol. Metab. 2008, 93, 3640–3643. [Google Scholar] [CrossRef]
- Liu, G.; Zhu, H.; Lagou, V.; Gutin, B.; Stallmann-Jorgensen, I.S.; Treiber, F.A.; Dong, Y.; Snieder, H. FTO variant rs9939609 is associated with body mass index and waist circumference, but not with energy intake or physical activity in European- and African-American youth. BMC Med. Genet. 2010, 11, 57. [Google Scholar]
- Sonestedt, E.; Roos, C.; Gullberg, B.; Ericson, U.; Wirfält, E.; Orho-Melander, M. Fat and carbohydrate intake modify the association between genetic variation in the FTO genotype and obesity. Am. J. Clin. Nutr. 2009, 90, 1418–1425. [Google Scholar] [CrossRef]
- Hainer, V.; Kunesova, M.; Bellisle, F.; Parizkova, J.; Braunerova, R.; Wagenknecht, M.; Lajka, J.; Hill, M.; Stunkard, A. The Eating Inventory, body adiposity and prevalence of diseases in a quota sample of Czech adults. Int. J. Obes. 2006, 30, 830–836. [Google Scholar] [CrossRef]
- Rideout, C.A.; Barr, S.I. “Restrained eating” vs. “trying to lose weight”: How are they associated with body weight and tendency to overeat among postmenopausal women? J. Am. Diet. Assoc. 2009, 109, 890–893. [Google Scholar] [CrossRef]
- Savage, J.S.; Hoffman, L.; Birch, L.L. Dieting, restraint, and disinhibition predict women’s weight change over 6 y. Am. J. Clin. Nutr. 2009, 90, 33–40. [Google Scholar] [CrossRef]
- Foster, G.D.; Wadden, T.A.; Swain, R.M.; Stunkard, A.J.; Platte, P.; Vogt, R.A. The eating inventory in obese women: Clinical correlates and relationship to weight loss. Int. J. Obes. 1998, 22, 778–785. [Google Scholar]
- Vogels, N.; Diepvens, K.; Westerterp-Plantenga, M. Predictors of long-term weight maintenance. Obes. Res. 2005, 13, 2162–2168. [Google Scholar] [CrossRef]
- Hill, J.O.; Wyatt, H.R.; Reed, G.W.; Peters, J.C. Obesity and the environment: Where do we go from here? Science 2003, 299, 853–855. [Google Scholar] [CrossRef]
- Bryant, E.J.; King, N.A.; Blundell, J.E. Disinhibition: Its effects on appetite and weight regulation. Obes. Rev. 2008, 9, 409–419. [Google Scholar] [CrossRef]
- Kilpeläinen, T.O.; Qi, L.; Brage, S.; Sharp, S.J.; Sonestedt, E.; Demerath, E.; Ahmad, T.; Mora, S.; Kaakinen, M.; Sandholt, C.H.; et al. Physical activity attenuates the influence of FTO variants on obesity risk: A meta-analysis of 218,166 adults and 19,268 children. PLoS Med. 2011, 8, e1001116. [Google Scholar] [CrossRef]
- Ahlberg, A.C.; Ljung, T.; Rosmond, R.; McEwen, B.; Holm, G.; Akesson, H.O.; Björntorp, P. Depression and anxiety symptoms in relation to anthropometry and metabolism in men. Psychiatry Res. 2002, 112, 101–110. [Google Scholar] [CrossRef]
- Davis, C. Psychobiological traits in the risk profile for overeating and weight gain. Int. J. Obes. 2009, 33 (Suppl. 2), S49–S53. [Google Scholar] [CrossRef]
- Rivera, M.; Cohen-Woods, S.; Kapur, K.; Breen, G.; Ng, M.Y.; Butler, A.W.; Craddock, N.; Gill, M.; Korszun, A.; Maier, W.; et al. Depressive disorder moderates the effect of the FTO gene on body mass index. Mol. Psychiatry 2012, 17, 604–611. [Google Scholar] [CrossRef]
- Samaan, Z.; Anand, S.; Zhang, X.; Desai, D.; Rivera, M.; Pare, G.; Thabane, L.; Xie, C.; Gerstein, H.; Engert, J.C.; et al. The protective effect of the obesity-associated rs9939609A variant in fat mass and obesity associated gene on depression. Mol. Psychiatry 2012, 18, 1281. [Google Scholar]
- Peeters, A.; Beckers, S.; Verrijken, A.; Roevens, P.; Peeters, P.; van Gaal, L.; van Hul, W. Variants in the FTO gene are associated with common obesity in the Belgian population. Mol. Genet. Metab. 2008, 93, 481–484. [Google Scholar] [CrossRef]
- Price, R.A.; Li, W.D.; Zhao, H. FTO gene SNPs associated with extreme obesity in cases, controls and extremely discordant sister pairs. BMC Med. Genet. 2008, 9, 4. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Harbron, J.; Van der Merwe, L.; Zaahl, M.G.; Kotze, M.J.; Senekal, M. Fat Mass and Obesity-Associated (FTO) Gene Polymorphisms Are Associated with Physical Activity, Food Intake, Eating Behaviors, Psychological Health, and Modeled Change in Body Mass Index in Overweight/Obese Caucasian Adults. Nutrients 2014, 6, 3130-3152. https://doi.org/10.3390/nu6083130
Harbron J, Van der Merwe L, Zaahl MG, Kotze MJ, Senekal M. Fat Mass and Obesity-Associated (FTO) Gene Polymorphisms Are Associated with Physical Activity, Food Intake, Eating Behaviors, Psychological Health, and Modeled Change in Body Mass Index in Overweight/Obese Caucasian Adults. Nutrients. 2014; 6(8):3130-3152. https://doi.org/10.3390/nu6083130
Chicago/Turabian StyleHarbron, Janetta, Lize Van der Merwe, Monique G. Zaahl, Maritha J. Kotze, and Marjanne Senekal. 2014. "Fat Mass and Obesity-Associated (FTO) Gene Polymorphisms Are Associated with Physical Activity, Food Intake, Eating Behaviors, Psychological Health, and Modeled Change in Body Mass Index in Overweight/Obese Caucasian Adults" Nutrients 6, no. 8: 3130-3152. https://doi.org/10.3390/nu6083130
APA StyleHarbron, J., Van der Merwe, L., Zaahl, M. G., Kotze, M. J., & Senekal, M. (2014). Fat Mass and Obesity-Associated (FTO) Gene Polymorphisms Are Associated with Physical Activity, Food Intake, Eating Behaviors, Psychological Health, and Modeled Change in Body Mass Index in Overweight/Obese Caucasian Adults. Nutrients, 6(8), 3130-3152. https://doi.org/10.3390/nu6083130