Fetal Adrenal Demedullation Lowers Circulating Norepinephrine and Attenuates Growth Restriction but not Reduction of Endocrine Cell Mass in an Ovine Model of Intrauterine Growth Restriction

Abstract
:1. Introduction
2. Experimental Section
2.1. Ethical Approval
2.2. IUGR and Control Fetuses
2.3. Surgical Preparation for Adrenal Demedullation and Cannulation
2.4. Postmortem Examination
2.5. Biochemical Analysis
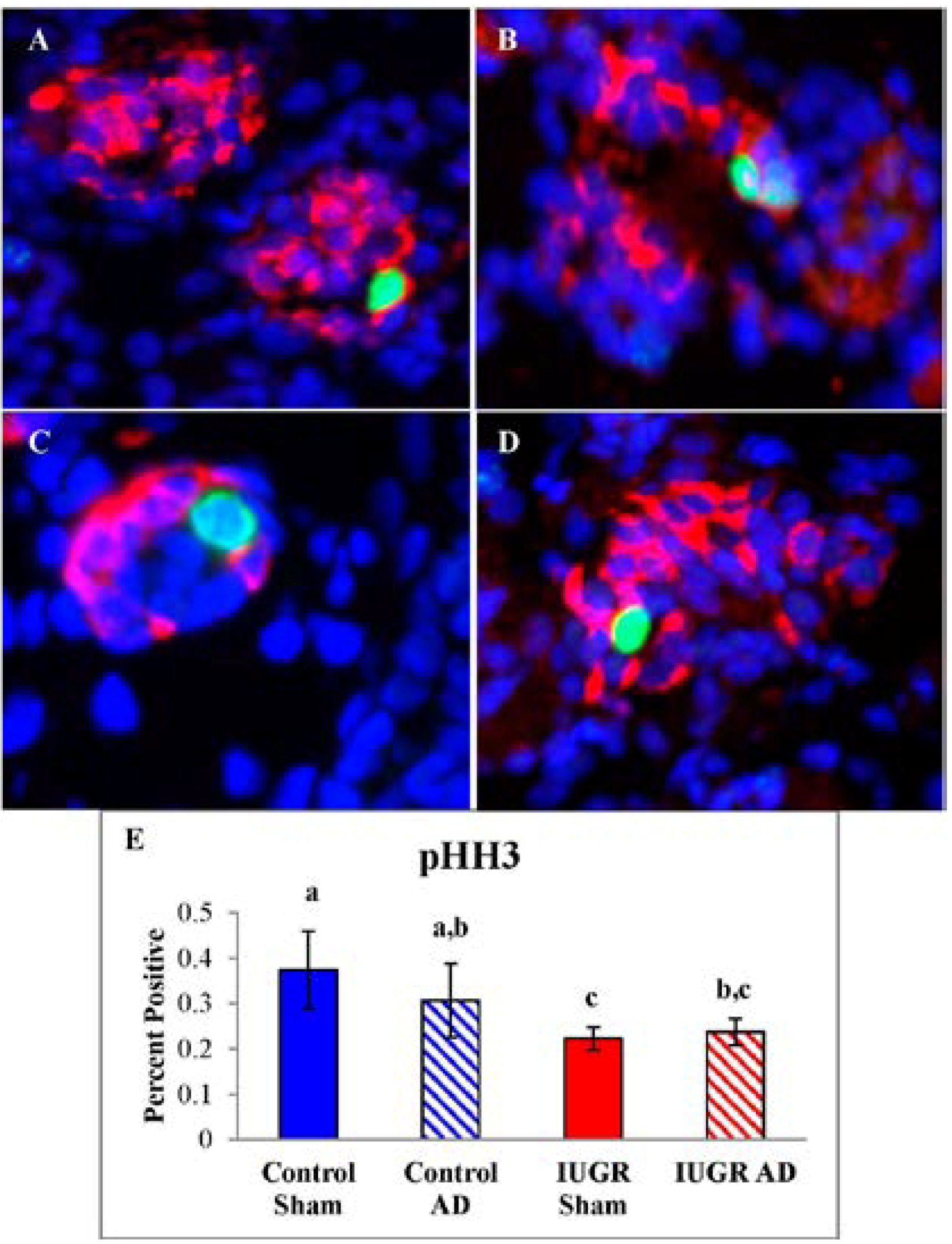
2.6. Pancreas Morphology and Pancreatic Endocrine Cell Proliferation
2.7. Statistical Analysis
3. Results
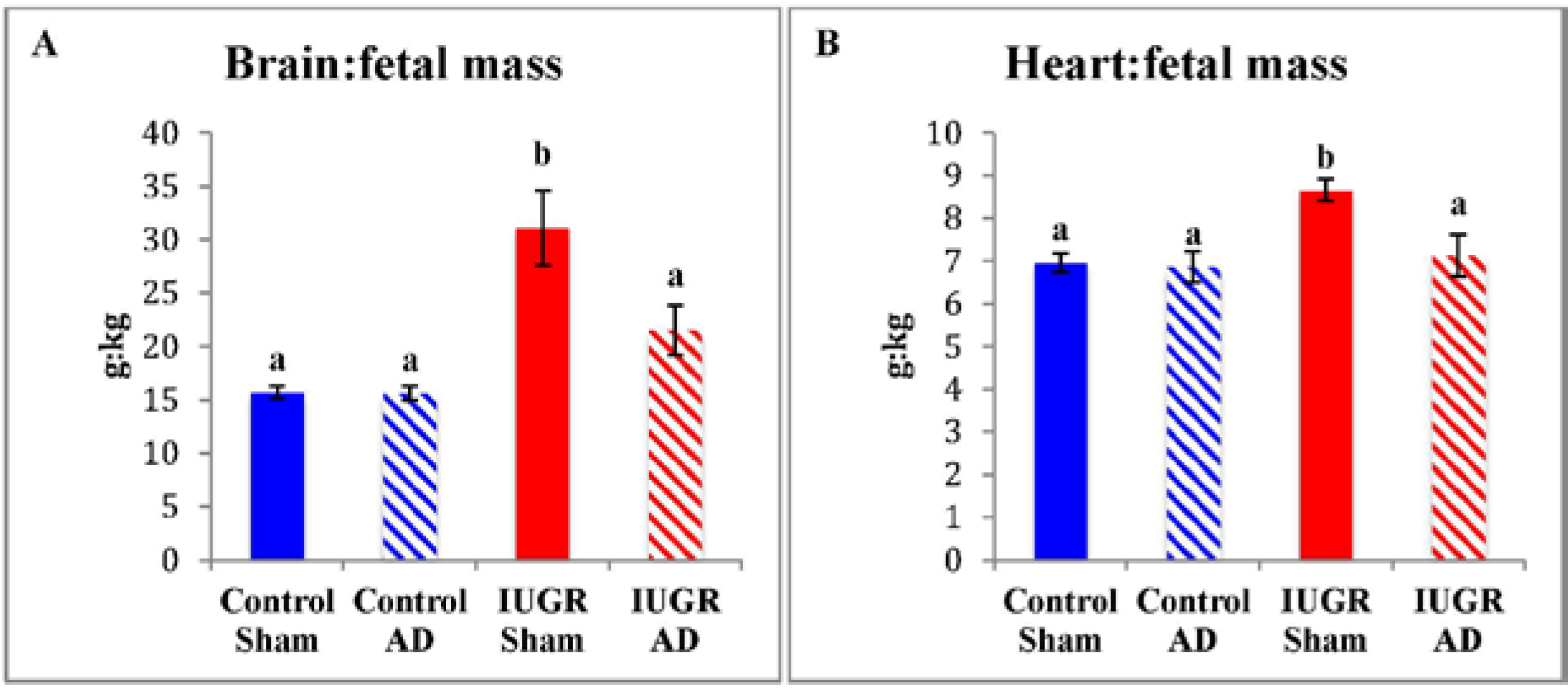
3.1. Fetal Biometry and Blood Measurements


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Control Sham (n = 8) | Control AD (n = 7) | IUGR Sham (n = 7) | IUGR AD (n = 4) |
|---|---|---|---|---|
| Fetus | 3250 ± 149 a | 3242 ± 177 a | 1577 ± 250 b | 2394 ± 414 c |
| Placenta | 319 ± 27 a | 272 ± 8.7 a | 137 ± 26 b | 160 ± 36 b |
| Brain | 50.5 ± 1.4 a | 50.0 ± 1.7 a | 40.9 ± 2.1 b | 48.8 ± 3.1 a |
| Heart | 22.6 ± 1.2 a | 22.3 ± 1.6 a | 12.2 ± 1.8 b | 16.6 ± 1.9 b |
| Kidneys | 22.4 ± 1.1 a | 23.3 ± 1.3 a | 11.7 ± 2.0 b | 17.3 ± 1.1 c |
| Liver | 96.5 ± 8.5 a | 88.3 ± 5.2 a,b | 45.0 ± 7.8 c | 63.5 ± 11 b,c |
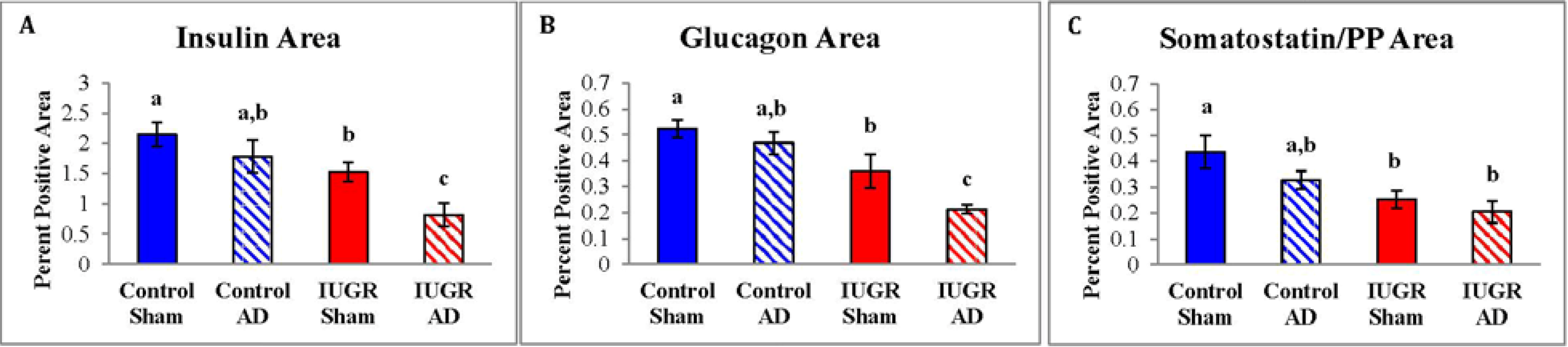
3.2. Fetal Pancreas Morphology



| Treatment Group | Control Sham (n = 7) | Control AD (n = 5) | IUGR Sham (n = 4) | IUGR AD (n = 3) |
|---|---|---|---|---|
| β-cell Mass | 75.2 ± 12 a | 51.6 ± 8.4 a,b | 30.9 ± 7.8 b | 22.2 ± 12 b |
| α-cell Mass | 18.8 ± 2.1 a | 12.2 ± 1.6 b | 8.6 ± 2.2 b | 5.5 ± 1.4 c |
| δ- and F-cell Mass | 15.7 ± 2.2 a | 9.2 ± 1.1 a | 7.0 ± 2.2 a | 6.1 ± 2.7 a |
| Total Endocrine Cell Mass | 109.7 ± 14 a | 73.0 ± 8.7 b | 46.5 ± 12 b | 33.8 ± 16 b |

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Barry, J.S.; Anthony, R.V. The pregnant sheep as a model for human pregnancy. Theriogenology 2008, 69, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Blondeau, B.; Avril, I.; Duchene, B.; Bréant, B. Endocrine pancreas development is altered in fetuses from rats previously showing intra-uterine growth retardation in response to malnutrition. Diabetologia 2002, 45, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Leos, R.; Anderson, M.; Chen, X.; Pugmire, J.; Anderson, K.A.; Limesand, S. Chronic exposure to elevated norepinephrine suppresses insulin secretion in fetal sheep with placental insufficiency and intrauterine growth restriction. Am. J. Physiol. Endrocrinol. Metab. 2010, 298, 770–778. [Google Scholar] [CrossRef]
- Limesand, S.W.; Jensen, J.; Hutton, C.J.; Hay, W.W., Jr. Diminished beta-cell replication contributes to reduced beta-cell mass in fetal sheep with intrauterine growth restriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, R1297–R1305. [Google Scholar] [CrossRef] [PubMed]
- Limesand, S.W.; Rozance, P.J.; Smith, D.; Hay, W.W., Jr. Increased insulin sensitivity and maintenance of glucose utilization rates in fetal sheep with placental insufficiency and intrauterine growth restriction. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E1716–E1725. [Google Scholar] [CrossRef] [PubMed]
- Regnault, T.R.; de Vrijer, B.; Battaglia, F.C. Transport and metabolism of amino acids in placenta. Endocrine 2002, 19, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Resnik, R. Intrauterine growth restriction. Obstet. Gynecol. 2002, 99, 490–496. [Google Scholar] [CrossRef] [PubMed]
- Setia, S; Sridhar, M.G.; Bhat, V.; Chaturvedula, L.; Vinayagamoorti, R.; John, M. Insulin sensitivity and insulin secretion at birth in intrauterine growth retarded infants. Pathology 2006, 38, 236–238. [Google Scholar]
- Simmons, R.A. Role of metabolic programming in the pathogenesis of β-cell failure in postnatal life. Rev. Endocr. Metab. Disord. 2007, 8, 95–104. [Google Scholar] [CrossRef] [PubMed]
- Thureen, P.J.; Trembler, K.A.; Meschia, G.; Makowski, E.L.; Wilkening, R.B. Placental glucose transport in heat-induced fetal growth retardation. Am. J. Physiol. 1992, 263, R578–R585. [Google Scholar] [PubMed]
- Green, A.S.; Rozance, P.J.; Limesand, S.W. Consequences of a compromised intrauterine environment on islet function. J. Endocrinol. 2010, 205, 211–224. [Google Scholar] [CrossRef] [PubMed]
- Hiraoka, T.; Kudo, T.; Kishimoto, Y. Catecholamines in experimentally growth-retarded rat fetus. AOFOG 1991, 17, 341–348. [Google Scholar]
- Jones, C.T.; Robinson, J.S. Studies on experimental growth retardation in sheep. Plasma catecholamines in fetuses with small placenta. J. Dev. Physiol. 1983, 5, 77–87. [Google Scholar] [PubMed]
- Langercrantz, H.; Sjoquist, B.; Bremme, K. Catecholamine metabolites in amniotic fluid as indicators of intrauterine stress. AJOG 1980, 136, 1067–1070. [Google Scholar]
- Rosengren, A.H.; Braun, M.; Mahdi, T.; Andersson, S.A.; Travers, M.E.; Shigeto, M.; Zhang, E.; Almgren, P.; Ladenvall, C.; Axelsson, A.S.; et al. Reduced insulin exocytosis in human pancreatic beta-cells with gene variants linked to type 2 diabetes. Diabetes 2012, 61, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.B.; Phillips, I.D.; Simonetta, G.; McMillen, I.C. Differential effects of increasing gestational age and placental restriction on tyrosine hydroxylase, phenylethanolamine N-methyltransferase, and proenkephalin A mRNA levels in the fetal sheep adrenal. J. Neurochem. 1998, 71, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Bassett, J.M.; Hanson, C. Catecholamines inhibit growth in fetal sheep in the absence of hypoxemia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 274, R1536–R1545. [Google Scholar]
- Jones, C.T.; Roebuck, M.M.; Walker, D.W.; Johnston, B.M. The role of the adrenal medulla and peripheral sympathetic nerves in the physiological responses of the fetal sheep to hypoxia. J. Dev. Physiol. 1988, 10, 17–36. [Google Scholar] [PubMed]
- Simonetta, G.; Young, I.R.; McMillen, I.C. Plasma catecholamine and met-enkephalin-Arg6-Phe7 responses to hypoxaemia after adrenalectomy in the fetal sheep. J. Auton. Nerv. Syst. 1996, 60, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.T.; Macko, A.R.; Chen, X.; Green, A.S.; Kelly, A.C.; Anderson, M.J.; Fowden, A.L.; Limesand, S.W. Hypoxaemia-induced catecholamine secretion from adrenal chromaffin cells inhibits glucose-stimulated hyperinsulinaemia in fetal sheep. J. Physiol. 2012, 590, 5439–5447. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.Y. Fetal adrenal medulla catecholamine response to hypoxia-direct and neural components. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 258, R1340–R1346. [Google Scholar]
- Cohen, W.R.; Piasecki, G.J.; Cohn, H.E.; Susa, J.B.; Jackson, B.T. Sympathoadrenal responses during hypoglycemia, hyperinsulinemia, and hypoxemia in the ovine fetus. Am. J. Physiol. Endocrinol. Metab. 1991, 261, E95–E102. [Google Scholar]
- Greenough, A.; Nicolaides, K.H.; Lagercrantz, H. Human fetal sympathoadrenal responsiveness. Early Hum. Dev. 1990, 23, 9–13. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.T.; Cohn, H.E.; Morrison, S.H.; Baker, R.M.; Piasecki, G.J. Hypoxia-induced sympathetic inhibition of the fetal plasma insulin response to hyperglycemia. Diabetes 1993, 42, 1621–1625. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.T.; Piasecki, G.J.; Cohn, H.E.; Cohen, W.R. Control of fetal insulin secretion. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R2179–R2188. [Google Scholar] [PubMed]
- Okamura, K.; Watanabe, T.; Tanigawara, S.; Endo, H.; Iwamoto, M.; Murotsuki, J.; Yajima, A. Catecholamine levels and their correlation to blood gases in umbilical venous blood obtained by cordocentesis. Fetal Diagn. Ther. 1990, 5, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Padbury, J.; Agata, Y.; Ludlow, J.; Ikegami, M.; Baylen, B.; Humme, J. Effect of fetal adrenalectomy on catecholamine release and physiologic adaptation at birth in sheep. J. Clin. Investig. 1987, 80, 1096–1103. [Google Scholar] [CrossRef] [PubMed]
- Phillippe, M.; Kitzmiller, J.L. The fetal and maternal catecholamine response to insulin-induced hypoglycemia in the rat. Am. J. Obstet. Gynecol. 1981, 139, 407–415. [Google Scholar] [PubMed]
- Yates, D.T.; Green, A.S.; Limesand, S.W. Catecholamines mediate multiple fetal adaptations during placental insufficiency that contribute to intrauterine growth restriction: Lessons from hyperthermic sheep. J. Pregnancy 2011, 10, 1–9. [Google Scholar] [CrossRef]
- Macko, A.R.; Yates, D.T.; Chen, X.; Green, A.S.; Kelly, A.C.; Brown, L.D.; Limesand, S.W. Elevated plasma norepinephrine inhibits insulin secretion, but adrenergic blockade reveals enhanced β-cell responsiveness in an ovine model of placental insufficiency at 0.7 of gestation. J. Dev. Orig. Health Dis. 2013, 4, 402–410. [Google Scholar] [CrossRef] [PubMed]
- Hay, W.W., Jr.; Meznarich, H.K.; Fowden, A.L. The effects of streptozotocin on rates of glucose utilization, oxidation, and production in the sheep fetus. Metabolism 1989, 38, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Green, A.S.; Macko, A.; Yates, D.T.; Kelly, A.C.; Limesand, S.W. Enhanced insulin secretion responsiveness and islet adrenergic desensitization after chronic norepinephrine suppression is discontinued in fetal sheep. Am. J. Physiol. Endocrinol. Metab. 2014, 306, E58–E64. [Google Scholar] [CrossRef] [PubMed]
- Limesand, S.W.; Rozance, P.J.; Macko, A.R.; Anderson, M.; Kelly, A.C.; Hay, W.W., Jr. Reductions in insulin concentrations and β-cell mass precede growth restriction in sheep fetuses with placental insufficiency. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E516–E523. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L. The role of insulin in fetal growth. Early Hum. Dev. 1992, 29, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Galan, H.L.; Anthony, R.V.; Rigano, S.; Parker, T.A.; de Vrijer, B.; Ferrazzi, E.; Wilkening, R.B.; Regnault, T.R. Fetal hypertension and abnormal Doppler velocimetry in an ovine model of intrauterine growth restriction. Am. J. Obstet. Gynecol. 2005, 192, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Yates, D.T.; Fowden, A.L.; Macko, A.R.; Chen, X.; Green, A.S.; Limesand, S.W. Adrenal demedullation abolishes hypoxemia-induced catecholamine suppression of glucose stimulated insulin secretion in fetal sheep. J. Pediatr. Res. 2010, 68, 179A. [Google Scholar]
- Yates, D.T.; Fowden, A.L.; Macko, A.R.; Chen, X.; Green, A.S.; Limesand, S.W. Elevated catecholamines are the predominant inhibitors of insulin secretion and contribute to altered metabolic phenotype during acute hypoxemia in fetal sheep. Reprod. Sci. 2011, 18, 222A. [Google Scholar]
- Limesand, S.W.; Hay, W.W., Jr. Adaptation of ovine fetal pancreatic insulin secretion to chronic hypoglycemia and euglycemic correction. J. Physiol. 2003, 547, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Cole, L.; Anderson, M.; Antin, P.B.; Limesand, S.W. One process for pancreatic beta-cell coalescence into islets involves an epithelial-mesenchymal transition. J. Endocrinol. 2009, 203, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.W.; Wilkening, R.B.; Meschia, G. Some aspects of placental function in chronically heat-stressed ewes. J. Dev. Physiol. 1987, 9, 17–29. [Google Scholar] [PubMed]
- Brown, L.D.; Rozance, P.J.; Thorn, S.R.; Friedman, J.E.; Hay, W.W., Jr. Acute supplementation of amino acids increases net protein accretion in IUGR fetal sheep. Am. J. Physiol. Endocrinol. Metab. 2012, 303, E352–E364. [Google Scholar] [CrossRef] [PubMed]
- De Vrijer, B.; Regnault, T.R.; Wilkening, R.B.; Meschia, G.; Battaglia, F.C. Placental uptake and transport of ACP, a neutral nonmetabolizable amino acid, in an ovine model of fetal growth restriction. Am. J. Physiol. Endocrinol. Metab. 2004, 287, E1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Regnault, T.R.; de Vrijer, B.; Galan, H.L.; Davidsen, M.L.; Trembler, K.A.; Battaglia, F.C.; Wilkening, R.B.; Anthony, R.V. The relationship between transplacental O2 diffusion and placental expression of PlGF, VEGF and their receptors in a placental insufficiency model of fetal growth restriction. J. Physiol. 2003, 550, 641–656. [Google Scholar] [CrossRef] [PubMed]
- Bocking, A.D.; Gagnon, R.; White, S.E.; Homan, J.; Milne, K.M.; Richardson, B.S. Circulatory responses to prolonged hypoxemia in fetal sheep. Am. J. Obstet. Gynecol. 1988, 159, 1418–1424. [Google Scholar] [CrossRef] [PubMed]
- Cohn, H.E.; Sacks, E.J.; Heymann, M.A.; Rudolph, A.M. Cardiovascular responses to hypoxemia and acidemia in fetal lambs. Am. J. Obstet. Gynecol. 1974, 120, 817–824. [Google Scholar] [PubMed]
- Fumia, F.D.; Edelstone, D.I.; Holzman, I.R. Blood flow and oxygen delivery to fetal organs as functions of fetal hematocrit. Am. J. Obstet. Gynecol. 1984, 150, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Peeters, L.L.; Sheldon, R.E.; Jones, M.D., Jr.; Makowski, E.L.; Meschia, G. Blood flow to fetal organs as a function of arterial oxygen content. Am. J. Obstet. Gynecol. 1979, 135, 637–646. [Google Scholar] [PubMed]
- Robinson, J.S.; Jones, C.T.; Thorburn, G.D. The effects of hypoxaemia in fetal sheep. J. Clin. Pathol. Suppl. 1977, 11, 127–133. [Google Scholar] [CrossRef]
- Block, B.S.; Schlafer, D.H.; Wentworth, R.A.; Kreitzer, L.A.; Nathanielsz, P.W. Regional blood flow distribution in fetal sheep with intrauterine growth retardation produced by decreased umbilical placental perfusion. J. Dev. Physiol. 1990, 13, 81–85. [Google Scholar] [PubMed]
- Kamitomo, M.; Alonso, J.G.; Okai, T.; Longo, L.D.; Gilbert, R.D. Effects of long-term, high-altitude hypoxemia on ovine fetal cardiac output and blood flow distribution. Am. J. Obstet. Gynecol. 1993, 169, 701–707. [Google Scholar] [CrossRef] [PubMed]
- Poudel, R.; McMillen, I.C.; Dunn, S.L.; Zhang, S.; Morrison, J.L. Impact of chronic hypoxemia on blood flow to the brain, heart and adrenal gland in the late gestation IUGR sheep fetus. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014. [Google Scholar] [CrossRef]
- Hoeflich, A.; Wu, M.; Mohan, S.; Foll, J.; Wanke, R.; Froehlich, T.; Arnold, G.J.; Lahm, H.; Kolb, H.J.; Wolf, E. Overexpression of insulin-like growth factor-binding protein-2 in transgenic mice reduces postnatal body weight gain. Endocrinology 1999, 140, 5488–5496. [Google Scholar] [CrossRef] [PubMed]
- Hooper, S.B.; Bocking, A.D.; White, S.E.; Fraher, L.J.; McDonald, T.J.; Han, V.K. Catecholamines stimulate the synthesis and release of insulin-like growth factor binding protein-1 (IGFBP-1) by fetal sheep liver in vivo. Endocrinology 1994, 134, 1104–1112. [Google Scholar] [PubMed]
- Hooper, S.B.; Bocking, A.D.; White, S.E.; Challis, J.R.; Han, V.K. DNA synthesis is reduced in selected fetal tissues during prolonged hypoxemia. Am. J. Physiol. 1991, 261, R508–R514. [Google Scholar] [PubMed]
- Lorlin, R.H.W.; Longo, L.D. Norepinephrine elevation in the fetal lamb: Oxygen consumption and cardiac output. Am. J. Physiol. 1980, 239, R115–R122. [Google Scholar] [PubMed]
- Milley, J.R. Ovine fetal metabolism during norepinephrine infusion. Am. J. Physiol. 1997, 273, E336–E347. [Google Scholar] [PubMed]
- Ross, J.C.; Fennessey, P.V.; Wilkening, R.B.; Battaglia, F.C.; Meschia, G. Placental transport and fetal utilization of leucine in a model of fetal growth retardation. Am. J. Physiol. 1996, 270, E491–E503. [Google Scholar]
- Carver, T.D.; Anderson, S.M.; Aldoretta, P.W.; Hay, W.W., Jr. Effect of low-level basal plus marked “pulsatile” hyperglycemia on insulin secretion in fetal sheep. Am. J. Physiol. 1996, 271, E865–E871. [Google Scholar] [PubMed]
- Andrews, S.E.; Brown, L.D.; Thorn, S.R.; Limesand, S.W.; Davis, M.; Hay, W.W., Jr.; Rozance, P.J. Increased adrenergic signaling is responsible for decreased glucose-stimulated insulin secretion in the chronically hyperinsulinemic ovine fetus. Endocrinology 2014. [Google Scholar] [CrossRef]
- Bhushan, A.; Itoh, N.; Kato, S.; Thiery, J.P.; Czernichow, P.; Bellusci, S.; Scharfmann, R. Fgf10 is essential for maintaining the proliferative capacity of epithelial progenitor cells during early pancreatic organogenesis. Development 2001, 128, 5109–5117. [Google Scholar] [PubMed]
- Jensen, J. Gene regulatory factors in pancreatic development. Dev. Dyn. 2004, 229, 176–200. [Google Scholar] [CrossRef] [PubMed]
- Petrik, J.; Arany, E.; McDonald, T.J.; Hill, D.J. Apoptosis in the pancreatic islet cells of the neonatal rat is associated with a reduced expression of insulin-like growth factor II that may act as a survival factor. Endocrinology 1998, 139, 2994–3004. [Google Scholar] [PubMed]
- Chen, X.; J Rozance, P.J.; Hay, W.W., Jr.; Limesand, S.W. Insulin-like growth factor and fibroblast growth factor expression profiles in growth-restricted fetal sheep pancreas. Exp. Biol. Med. 2012, 237, 524–529. [Google Scholar] [CrossRef]
- Rozance, P.J.; Anderson, M.; Martinez, M.; Fahy, A.; Macko, A.R.; Kailey, J.; Seedorf, G.J.; Abman, S.H.; Hay, W.W., Jr.; Limesand, S.W. Placental insufficiency decreases pancreatic vascularity and disrupts hepatocyte growth factor signaling in the pancreatic islet endothelial cell in fetal sheep. Diabetes 2014. [Google Scholar] [CrossRef]
- Limesand, S.W.; Rozance, P.J.; Zerbe, G.O.; Hutton, J.C.; Hay, W.W., Jr. Attenuated insulin release and storage in fetal sheep pancreatic islets with intrauterine growth restriction. Endocrinology 2006, 147, 1488–1497. [Google Scholar] [CrossRef] [PubMed]
- Sperling, M.A.; Christensen, R.A.; Ganguli, S.; Anand, R. Adrenergic modulation of pancreatic hormone secretion in utero: Studies in fetal sheep. Pediatr. Res. 1980, 14, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Vuguin, P.M.; Kedees, M.H.; Cui, L.; Guz, Y.; Gelling, R.W.; Nejathaim, M.; Charron, M.J.; Teitelman, G. Ablation of the glucagon receptor gene increases fetal lethality and produces alterations in islet development and maturation. Endocrinology 2006, 147, 3995–4006. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; MacDougall, M.L.; McDowell, M.T.; Xi, L.; Wei, R.; Zavadoski, W.J.; Molloy, M.P.; Baker, J.D.; Kuhn, M.; Cabrera, O.; et al. Polyomic profiling reveals significant hepatic metabolic alterations in glucagon-receptor (GCGR) knockout mice: Implications on anti-glucagon therapies for diabetes. BMC Genomics 2011, 12, 281. [Google Scholar] [CrossRef] [PubMed]
- Gelling, R.W.; Du, X.Q.; Dichmann, D.S.; Romer, J.; Huang, H.; Cui, L.; Obici, S.; Tang, B.; Holst, J.J.; Fledelius, C.; et al. Lower blood glucose, hyperglucagonemia, and pancreatic alpha cell hyperplasia in glucagon receptor knockout mice. Proc. Natl. Acad. Sci. USA 2003, 100, 1438–1443. [Google Scholar] [CrossRef] [PubMed]
- Ouhilal, S.; Vuguin, P.; Lingguang, C.; Xiu-Quan, D.; Gelling, R.W.; Reznik, S.E.; Russell, R.; Parlow, A.F.; Karpovsky, C.; Santoro, N.; et al. Hypoglycemia, hyperglucagonemia, and fetoplacental defects in glucagon receptor knockout mice: A role for glucagon action in pregnancy maintenance. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E522–E531. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, D.; Christou, H. Current concepts in intrauterine growth restriction. J. Intensive Care Med. 2004, 19, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Oliver, E.; Malin, G.; Khan, K.S.; Meads, C. Effectiveness of interventions for the prevention of small-for-gestational age fetuses and perinatal mortality: A review of systematic reviews. AOGS 2013, 92, 143–151. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Davis, M.A.; Macko, A.R.; Steyn, L.V.; Anderson, M.J.; Limesand, S.W. Fetal Adrenal Demedullation Lowers Circulating Norepinephrine and Attenuates Growth Restriction but not Reduction of Endocrine Cell Mass in an Ovine Model of Intrauterine Growth Restriction. Nutrients 2015, 7, 500-516. https://doi.org/10.3390/nu7010500
Davis MA, Macko AR, Steyn LV, Anderson MJ, Limesand SW. Fetal Adrenal Demedullation Lowers Circulating Norepinephrine and Attenuates Growth Restriction but not Reduction of Endocrine Cell Mass in an Ovine Model of Intrauterine Growth Restriction. Nutrients. 2015; 7(1):500-516. https://doi.org/10.3390/nu7010500
Chicago/Turabian StyleDavis, Melissa A., Antoni R. Macko, Leah V. Steyn, Miranda J. Anderson, and Sean W. Limesand. 2015. "Fetal Adrenal Demedullation Lowers Circulating Norepinephrine and Attenuates Growth Restriction but not Reduction of Endocrine Cell Mass in an Ovine Model of Intrauterine Growth Restriction" Nutrients 7, no. 1: 500-516. https://doi.org/10.3390/nu7010500
APA StyleDavis, M. A., Macko, A. R., Steyn, L. V., Anderson, M. J., & Limesand, S. W. (2015). Fetal Adrenal Demedullation Lowers Circulating Norepinephrine and Attenuates Growth Restriction but not Reduction of Endocrine Cell Mass in an Ovine Model of Intrauterine Growth Restriction. Nutrients, 7(1), 500-516. https://doi.org/10.3390/nu7010500