Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates


Abstract
:1. Introduction
2. Experimental Section
2.1. Hemp Seed Products and Chemical Reagents
2.2. Preparation of Hemp Seed Protein Isolates (HPI)
2.3. Preparation of Enzymatic Hemp Seed Protein Hydrolysates (HPHs)
2.4. Determination of Degree of Hydrolysis
2.5. Amino Acid Composition Analysis
2.6. Analysis of Molecular Weight Distribution
2.7. Intrinsic Fluorescence
2.8. ACE Inhibition Assay
2.9. Renin Inhibition Assay
2.10. BP-lowering Effect of Peptides in Spontaneously Hypertensive Rats (SHRs)
2.11. Statistical Analysis
3. Results
3.1. Amino Acid Composition of HPI and HPHs
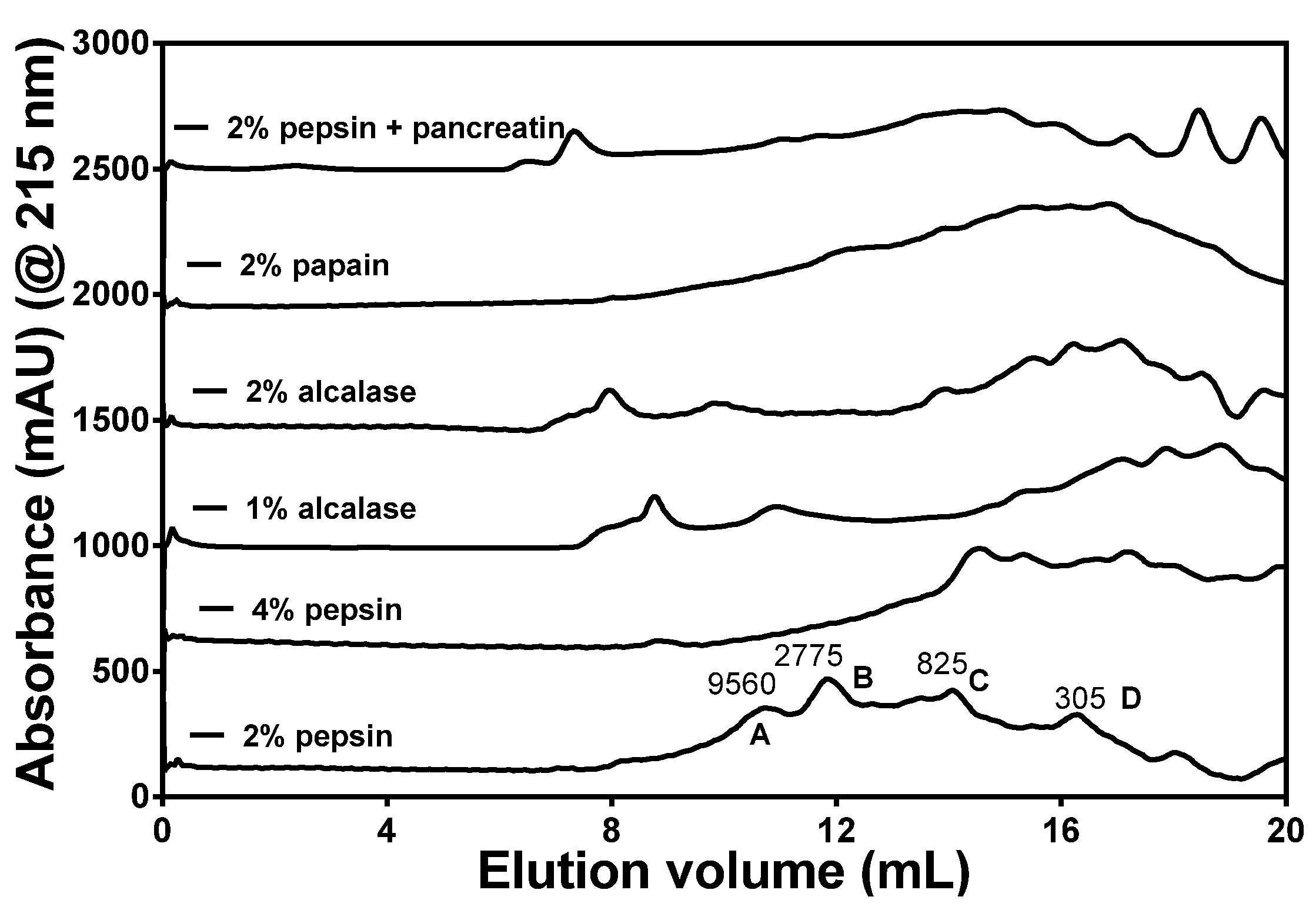
3.2. DH and Size Exclusion Chromatography Analysis of HPHs
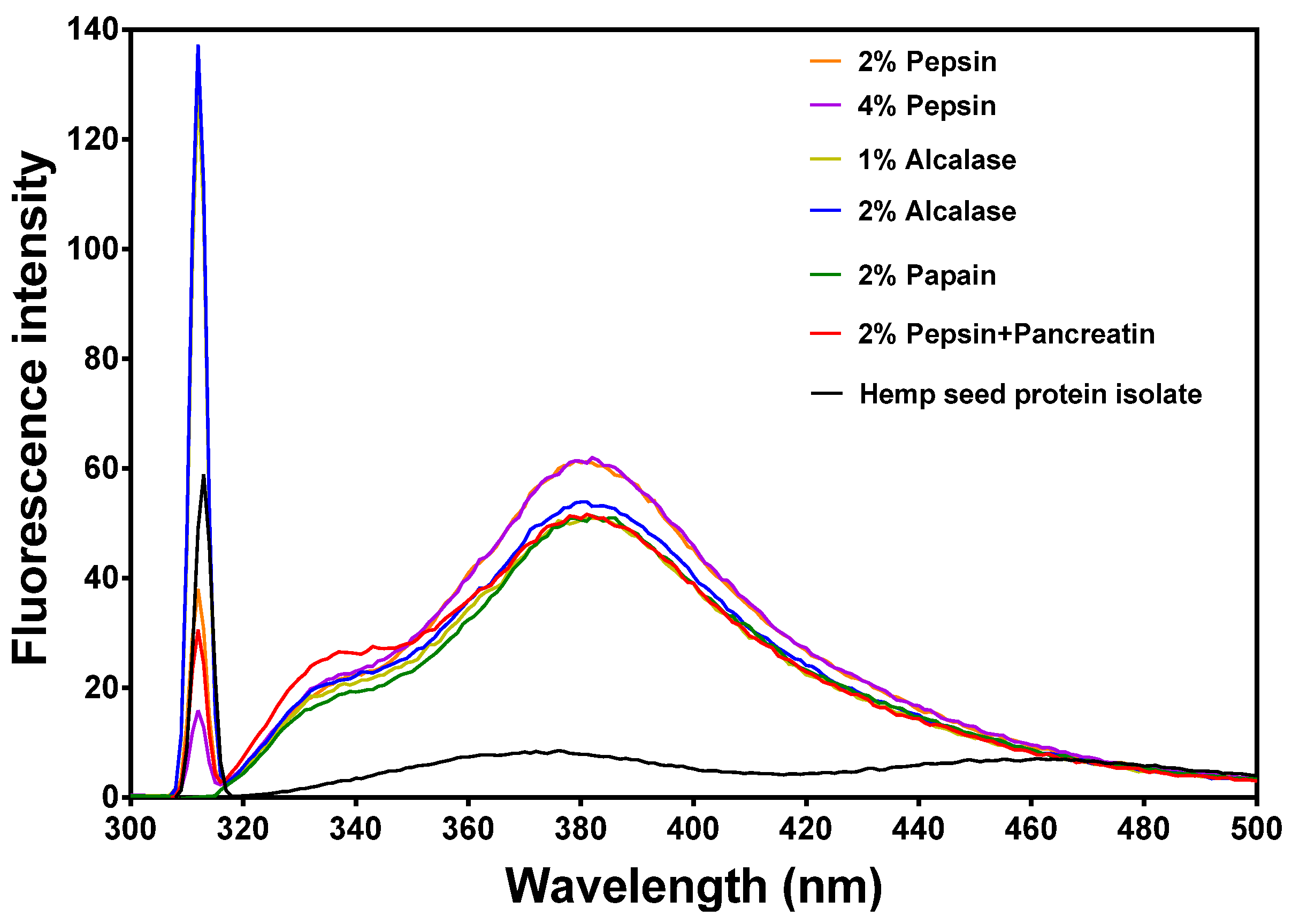
3.3. Intrinsic Fluorescence Properties

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid | HPI | 2% Pepsin | 4% Pepsin | 1% Alcalase | 2% Alcalase | 2% Papain | 2% Pepsin + Pancreatin | Mean ± SD | p-Value |
|---|---|---|---|---|---|---|---|---|---|
| Asx a | 11.31 | 11.36 | 11.37 | 11.51 | 11.55 | 11.18 | 10.74 | 11.29 ± 0.27 | 1.00 |
| Thr | 3.42 | 2.92 | 2.85 | 2.89 | 2.78 | 2.93 | 2.82 | 2.95 ± 0.22 | 0.14 |
| Ser | 5.58 | 5.98 | 5.95 | 6.10 | 6.09 | 6.00 | 5.84 | 5.94 ± 0.18 | 1.00 |
| Glx b | 19.10 | 19.36 | 19.28 | 19.63 | 19.64 | 19.56 | 19.06 | 19.38 ± 0.24 | 1.00 |
| Pro | 4.30 | 4.94 | 4.74 | 4.83 | 4.75 | 5.23 | 5.17 | 4.85 ± 0.31 | 1.00 |
| Gly | 4.31 | 3.82 | 3.84 | 3.73 | 3.70 | 3.63 | 3.56 | 3.80 ± 0.25 | 1.00 |
| Ala | 3.81 | 4.79 | 4.76 | 4.53 | 4.60 | 4.75 | 4.48 | 4.53 ± 0.34 | 0.32 |
| Cys | 1.25 | 0.12 | 1.34 | 1.20 | 1.22 | 1.31 | 1.43 | 1.13 ± 0.45 | 0.50 |
| Val | 4.66 | 4.60 | 4.33 | 4.25 | 4.20 | 4.36 | 4.71 | 4.45 ± 0.21 | 0.32 |
| Met | 1.92 | 2.26 | 2.18 | 2.28 | 2.24 | 2.34 | 2.21 | 2.20 ± 0.14 | 1.00 |
| Ile | 3.63 | 3.17 | 3.04 | 2.95 | 2.90 | 3.03 | 3.33 | 3.15 ± 0.26 | 0.14 |
| Leu | 6.46 | 6.64 | 6.48 | 6.38 | 6.39 | 6.35 | 6.60 | 6.48 ± 0.11 | 1.00 |
| Tyr | 3.61 | 3.49 | 3.39 | 3.39 | 3.38 | 3.28 | 3.62 | 3.45 ± 0.13 | 1.00 |
| Phe | 4.72 | 4.65 | 4.52 | 4.45 | 4.48 | 4.44 | 4.80 | 4.58 ± 0.14 | 0.32 |
| His | 3.15 | 3.09 | 3.13 | 3.12 | 3.13 | 3.02 | 3.00 | 3.10 ± 0.06 | 0.14 |
| Lys | 2.73 | 3.19 | 3.32 | 2.96 | 3.03 | 3.35 | 3.33 | 3.13 ± 0.23 | 0.14 |
| Arg | 15.06 | 14.63 | 14.47 | 14.77 | 14.80 | 14.24 | 14.21 | 14.60 ± 0.31 | 1.00 |
| Trp | 1.00 | 1.01 | 1.01 | 1.05 | 1.11 | 1.00 | 1.08 | 1.04 ± 0.04 | 0.50 |



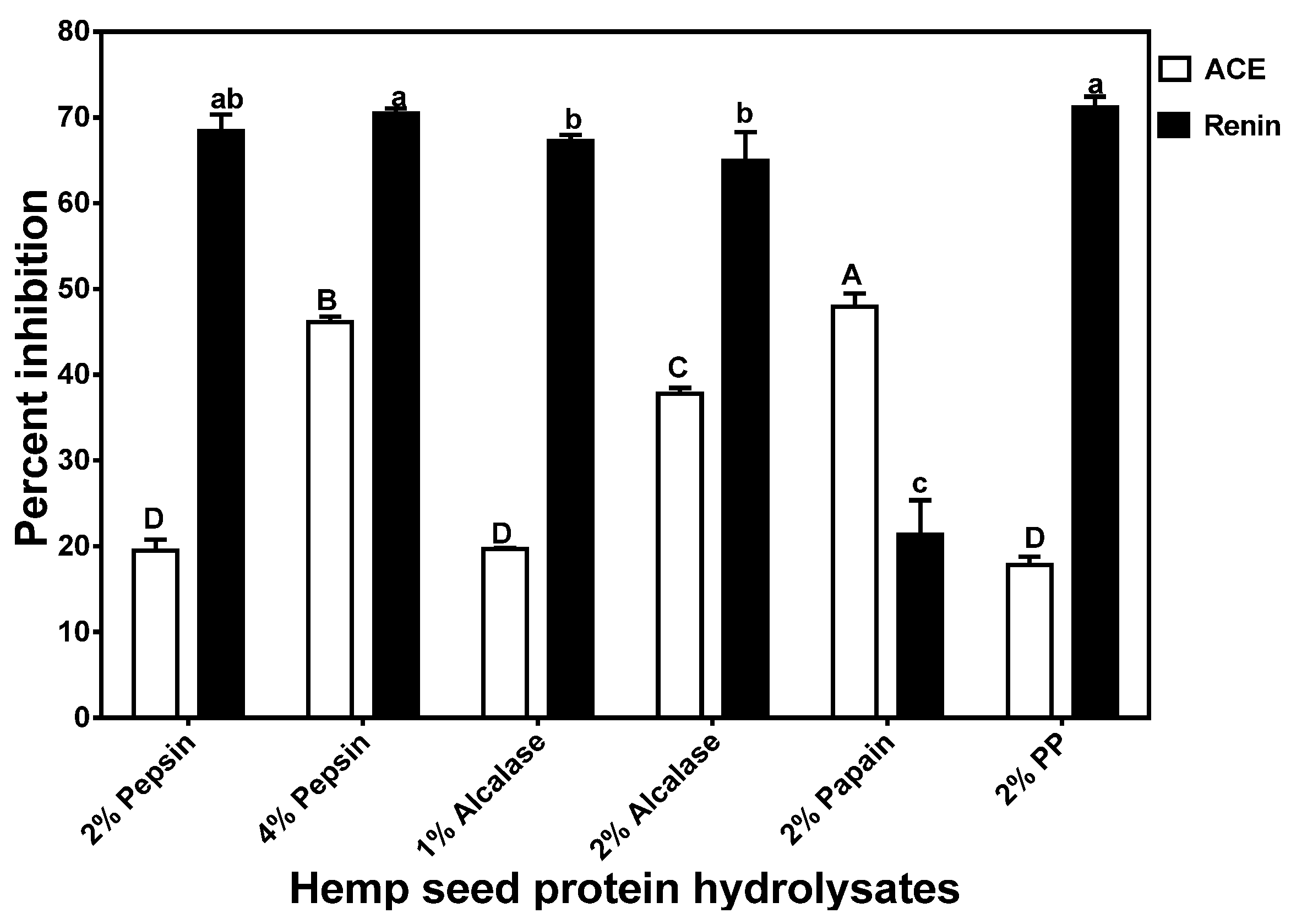
3.4. In Vitro Inhibition of ACE and Renin Activities of HPHs


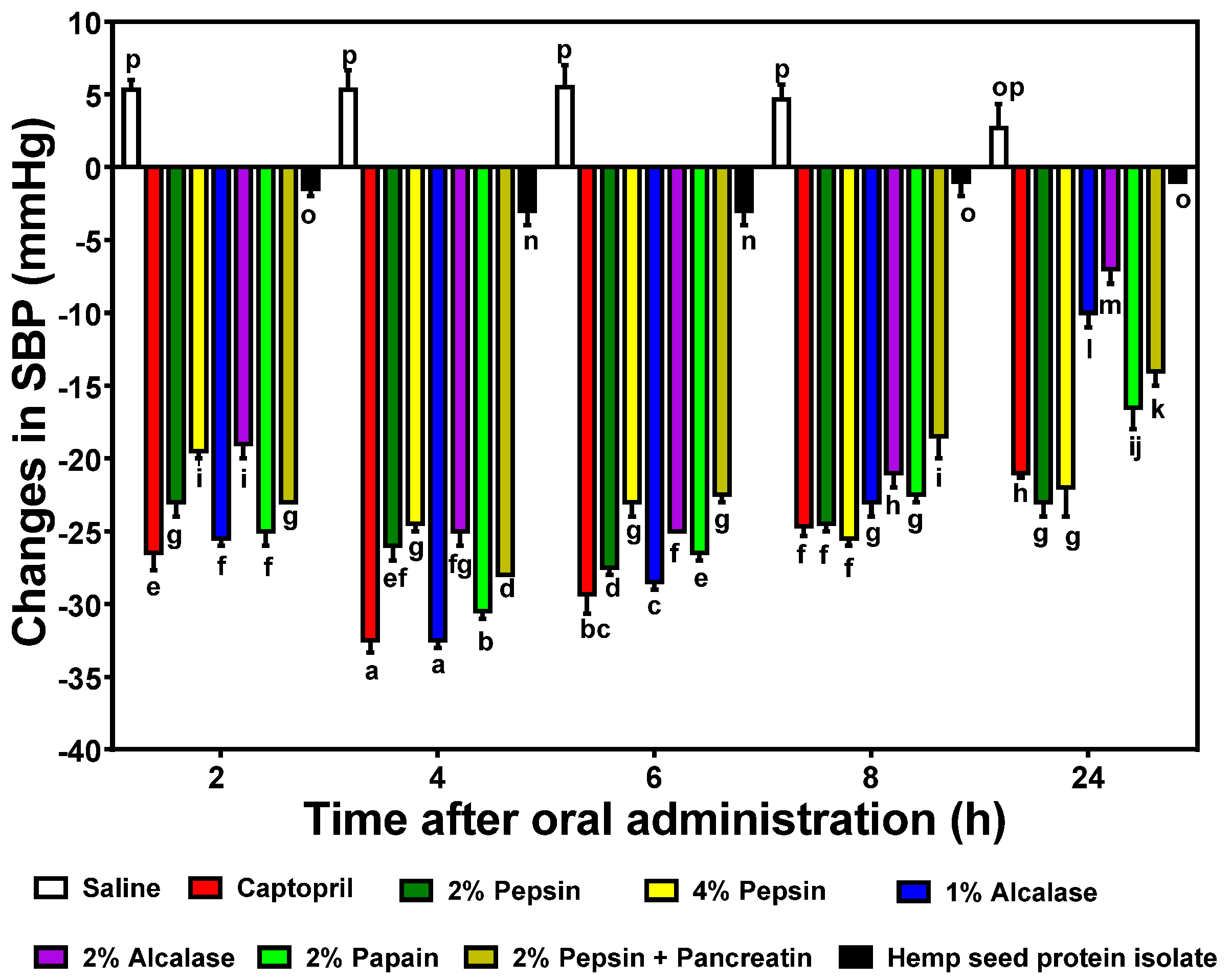
3.5. In vivo Reduction of Blood Pressure by HPHs

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- World Health Organization (WHO). A global brief on hypertension. In Silent Killer, Global Public Health Crisis; WHO: Geneva, USA, 2013; p. 40. Available online: http://www.who.int/cardiovascular_diseases/publications/global_brief_hypertension/en/ (accessed on 4 July 2015).
- Guessous, I.; Bochud, M.; Theler, J.M.; Gaspoz, J.M.; Pechère-Bertschi, A. 1999–2009 Trends in prevalence, unawareness, treatment and control of hypertension in Geneva, Switzerland. PLoS One 2012, 7, e39877. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; McAdam-Marx, C. The determinants of antihypertensive use and expenditure in patients with hypertension in the USA. J. Pharm. Health Serv. Res. 2014, 5, 11–18. [Google Scholar] [CrossRef]
- Anderson, I.; Ivatts, S.; Somanathan, A.; Rolfe, B. The challenges of health financing in five South Pacific countries. Asian-Pacific Econ. Lit. 2014, 28, 29–46. [Google Scholar] [CrossRef]
- Daien, V.; Duny, Y.; Ribstein, J.; Du, C.G.; Mimran, A.; Villain, M.; Daures, J.P.; Fesler, P. Treatment of hypertension with renin-angiotensin system inhibitors and renal dysfunction: A systematic review and meta-analysis. Am. J. Hypertens. 2012, 25, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Antihypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef] [PubMed]
- Pihlanto, A.; Mäkinen, S. Antihypertensive properties of plant protein derived peptides. In Bioactive Food Peptides in Health and Disease; Hernandez-Ledesma, B., Hsieh, C., Eds.; INTECH Publishers: Rijeka, Croatia, 2013; pp. 145–182. [Google Scholar]
- Norris, R.; FitzGerald, R. Antihypertensive peptides from food proteins. In Bioactive Food Peptides in Health and Disease; Hernandez-Ledesma, B., Hsieh, C., Eds.; INTECH Publishers: Rijeka, Croatia, 2013; pp. 45–72. [Google Scholar]
- He, R.; Alashi, A.; Malomo, S.A.; Girgih, A.T.; Chao, D.; Ju, X.; Aluko, R.E. Antihypertensive and free radical scavenging properties of enzymatic rapeseed protein hydrolysates. Food Chem. 2013, 141, 153–159. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Malomo, S.A.; Alashi, A.; Girgih, A.T.; Ju, X.; Aluko, R.E. Purification and hypotensive activity of rapeseed protein-derived renin and angiotensin converting enzyme inhibitory peptides. J. Funct. Foods 2013, 5, 781–789. [Google Scholar] [CrossRef]
- He, R.; Malomo, S.A.; Girgih, A.T.; Ju, X.; Aluko, R.E. Glycinyl-histidinyl-serine (GHS), a novel rapeseed protein-derived peptide has blood pressure-lowering effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2013, 61, 8396–8402. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; Udenigwe, C.C.; Li, H.; Adebiyi, A.P.; Aluko, R.E. Kinetics of enzyme inhibition and antihypertensive effects of hemp seed (Cannabis sativa L.) protein hydrolysate. J. Am. Oil Chem. Soc. 2011, 88, 1767–1774. [Google Scholar] [CrossRef]
- Girgih, A.T.; He, R.; Alashi, A.M.; Malomo, S.A.; Aluko, R.E. Preventive and treatment effects of hemp seed (Cannabis sativa L.) meal protein hydrolysate against blood pressure in spontaneously hypertensive rats. Eur. J. Nutr. 2014, 53, 1237–1246. [Google Scholar] [CrossRef] [PubMed]
- Girgih, A.T.; He, R.; Malomo, S.A.; Aluko, R.E. Structural and functional characterization of hemp seed (Cannabis sativa L.) protein-derived antioxidant and antihypertensive peptides. J. Funct. Foods 2014, 6, 384–394. [Google Scholar] [CrossRef]
- Mundi, S.; Aluko, R.E. Inhibitory properties of kidney bean protein hydrolysate and its membrane fractions against renin, angiotensin converting enzyme, and free radicals. Austin J. Nutr. Food Sci. 2014, 2, 1–11. [Google Scholar]
- Fitzgerald, C.; Aluko, R.E.; Hossain, M.; Rai, D.K.; Hayes, M. Potential of a renin inhibitory peptide from the red seaweed (Palmaria palmata) as a functional food ingredient following confirmation and characterization of a hypotensive effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2014, 62, 8352–8356. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Jia, J.; Fang, M.; Zhang, L.; Guo, M.; Xie, J.; Xia, Y.; Zhou, L.; Wei, D. In vitro and in vivo ACE inhibitory of pistachio hydrolysates and in silico mechanism of identified peptide binding with ACE. Process Biochem. 2014, 49, 898–904. [Google Scholar] [CrossRef]
- Zhang, C.; Cao, W.; Hong, P.; Ji, H.; Qin, X.; He, J. Angiotensin I-converting enzyme inhibitory activity of Acetes chinensis peptic hydrolysate and its antihypertensive effect in spontaneously hypertensive rats. Int. J. Food Sci. Technol. 2009, 44, 2042–2048. [Google Scholar] [CrossRef]
- Wang, C.; Tian, J.; Wang, Q. ACE inhibitory and antihypertensive properties of apricot almond meal hydrolysate. Eur. Food Res. Technol. 2011, 232, 549–556. [Google Scholar] [CrossRef]
- Huang, W.H.; Sun, J.; He, H.; Dong, H.W.; Li, J.T. Antihypertensive effect of corn peptides, produced by a continuous production in enzymatic membrane reactor, in spontaneously hypertensive rats. Food Chem. 2011, 128, 968–973. [Google Scholar] [CrossRef]
- Doyen, A.; Udenigwe, C.C.; Mitchell, P.L.; Marette, A.; Aluko, R.E. Anti-diabetic and antihypertensive activities of two flaxseed protein hydrolysate fractions revealed following their simultaneous separation by electrodialysis with ultrafiltration membranes. Food Chem. 2014, 145, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Alashi, A.M.; Blanchard, C.L.; Mailer, R.J.; Agboola, S.O.; Mawson, A.J.; He, R.; Malomo, S.A.; Girgih, A.T.; Aluko, R.E. Blood pressure lowering effects of Australian canola protein hydrolysates in spontaneously hypertensive rats. Food Res. Int. 2014, 55, 281–287. [Google Scholar] [CrossRef]
- Li, H.; Prairie, N.; Udenigwe, C.C.; Adebiyi, A.P.; Tappia, P.S.; Aukema, H.M.; Jones, P.J.H.; Aluko, R.E. Blood pressure lowering effect of a pea protein hydrolysate in hypertensive rats and humans. J. Agric. Food Chem. 2011, 59, 9854–9860. [Google Scholar] [CrossRef] [PubMed]
- Siow, H.; Gan, C. Extraction of antioxidative and antihypertensive bioactive peptides from Parkia speciosa seeds. Food Chem. 2013, 141, 3435–3442. [Google Scholar] [CrossRef] [PubMed]
- Marczak, E.; Usui, H.; Fujita, H.; Yang, Y.; Yokoo, M.; Lipkowski, A.; Yoshikawa, M. New antihypertensive peptides isolated from rapeseed. Peptides 2003, 24, 791–798. [Google Scholar] [CrossRef]
- Ambigaipalan, P.; Al-Khalifa, A.S.; Shahidi, F. Antioxidant and angiotensin I converting enzyme inhibitory activities of date seed protein hydrolysates prepared using Alcalase, Flavourzyme and Thermolysin. J. Funct. Foods 2015. [Google Scholar] [CrossRef]
- Markwell, M.A.K.; Haas, S.M.; Bieber, L.L.; Tolbert, N.E. Modification of Lowry procedure to simplify protein determination in membrane and lipoprotein samples. Anal. Biochem. 1978, 87, 206–210. [Google Scholar] [CrossRef]
- Adler-Nissen, J. Determination of the degree of hydrolysis of food protein hydrolysates by trinitrobenzenesulfonic acid. J. Agric. Food Chem. 1979, 27, 1256–1262. [Google Scholar] [CrossRef] [PubMed]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L. Rapid analysis of amino acids using pre-column derivatization. J. Chrom. B 1984, 336, 93–104. [Google Scholar] [CrossRef]
- Gehrke, C.W.; Wall, L.L.; Absheer, J.S.; Kaiser, F.E.; Zumwalt, R.W. Sample preparation for chromatography of amino-acids: Acid hydrolysis of proteins. J. Assoc. Off. Anal. Chem. 1985, 68, 811–821. [Google Scholar]
- Landry, J.; Delhaye, S. Simplified procedure for the determination of tryptophan of foods and feedstuffs from barytic hydrolysis. J. Agric. Food Chem. 1992, 40, 776–779. [Google Scholar] [CrossRef]
- Li, H.; Aluko, R.E. Structural modulation of calmodulin and calmodulin-dependent protein kinase II by pea protein hydrolysates. Int. J. Food Sci. Nutr. 2006, 57, 178–189. [Google Scholar] [CrossRef] [PubMed]
- Holmquist, B.; Bunning, P.; Riordan, J.F. A continuous spectrophotometric assay for angiotensin converting enzyme. Anal. Biochem. 1979, 95, 540–548. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2007, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Garcia, E.; Marina, M.L.; Garcia, M.C. Plum (Prunus domestica L.) by-product as a new and cheap source of bioactive peptides: Extraction method and peptides characterization. J. Funct. Foods 2014, 11, 428–437. [Google Scholar] [CrossRef]
- Ishiguro, K.; Sameshima, Y.; Kume, T.; Ikeda, K.; Matsumoto, J.; Yoshimoto, M. Hypotensive effect of a sweet potato protein digests in spontaneously hypertensive rats and purification of angiotensin I-converting enzyme inhibitory peptides. Food Chem. 2012, 131, 774–779. [Google Scholar] [CrossRef]
- Durak, A.; Baraniak, B.; Jakubczyk, A.; Swieca, M. Biologically active peptides obtained by enzymatic hydrolysis of Adzuki bean seeds. Food Chem. 2013, 141, 2177–2183. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Salas, P.A.; Montero-Moran, G.M.; Martínez-Cuevas, P.P.; Gonzalez, C.; de la Rosa, A.P.B. Characterization of antidiabetic and antihypertensive properties of canary seed (Phalaris canariensis L.) peptides. J. Agric. Food Chem. 2014, 62, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; FitzGerald, R.J. In vitro assessment of cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Fitzgerald, C.; Mora-Soler, L.; Gallagher, E.; O’Connor, P.; Prieto, J.; Soler-Vila, A.; Hayes, M. Isolation and characterization of bioactive pro-peptides with in vitro renin inhibitory activities from the macroalga Palmaria palmata. J. Agric. Food Chem. 2012, 60, 7421–7427. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Lin, Y.S.; Hou, W.C.; Aluko, R.E. Kinetics of the inhibition of renin and angiotensin I-converting enzyme by flaxseed protein hydrolysate fractions. J. Funct. Foods 2009, 1, 199–207. [Google Scholar] [CrossRef]
- Schmid, F.X. Spectra methods of characterizing protein conformation and conformational changes. In Protein Structure: A Practical Approach; Creighton, T.E., Ed.; Springer Publishing Co.: New York City, NY, USA, 1989; pp. 251–285. [Google Scholar]
- Nakamura, Y.; Yamamoto, N.; Sakai, K.; Takano, T. Antihypertensive effect of sour milk and peptides isolated from it that are inhibitors of angiotensin converting enzyme. J. Dairy Sci. 1995, 78, 1253–1257. [Google Scholar] [CrossRef]
- Li, H.; Aluko, R.E. Identification and inhibitory properties of multifunctional peptides from pea protein hydrolysate. J. Agric. Food Chem. 2010, 58, 11471–11476. [Google Scholar] [CrossRef] [PubMed]
- Aluko, R.E. Structure and function of plant protein-derived antihypertensive peptides. Curr. Op. Food Sci. 2015, 4, 44–50. [Google Scholar] [CrossRef]
- Onuh, J.O.; Girgih, A.T.; Malomo, S.A.; Aluko, R.E.; Aliani, M. Kinetics of in vitro renin and angiotensin converting enzyme inhibition by chicken skin protein hydrolysates and their blood pressure lowering effects in spontaneously hypertensive rats. J. Funct. Foods 2015, 14, 133–143. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malomo, S.A.; Onuh, J.O.; Girgih, A.T.; Aluko, R.E. Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates. Nutrients 2015, 7, 7616-7632. https://doi.org/10.3390/nu7095358
Malomo SA, Onuh JO, Girgih AT, Aluko RE. Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates. Nutrients. 2015; 7(9):7616-7632. https://doi.org/10.3390/nu7095358
Chicago/Turabian StyleMalomo, Sunday A., John O. Onuh, Abraham T. Girgih, and Rotimi E. Aluko. 2015. "Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates" Nutrients 7, no. 9: 7616-7632. https://doi.org/10.3390/nu7095358
APA StyleMalomo, S. A., Onuh, J. O., Girgih, A. T., & Aluko, R. E. (2015). Structural and Antihypertensive Properties of Enzymatic Hemp Seed Protein Hydrolysates. Nutrients, 7(9), 7616-7632. https://doi.org/10.3390/nu7095358