DHA Supplementation Alone or in Combination with Other Nutrients Does not Modulate Cerebral Hemodynamics or Cognitive Function in Healthy Older Adults

 ,
,
Abstract
:1. Introduction
2. Experimental Section
2.1. Participants
2.2. Study Design

2.2.1. Near Infrared Spectroscopy (NIRS)
2.2.2. Cognitive Tasks
2.2.3. Blood Sample Collection and Preparation
2.2.4. Whole Blood Fatty Acid Analysis
2.3. Procedure
2.4. Statistical Analysis
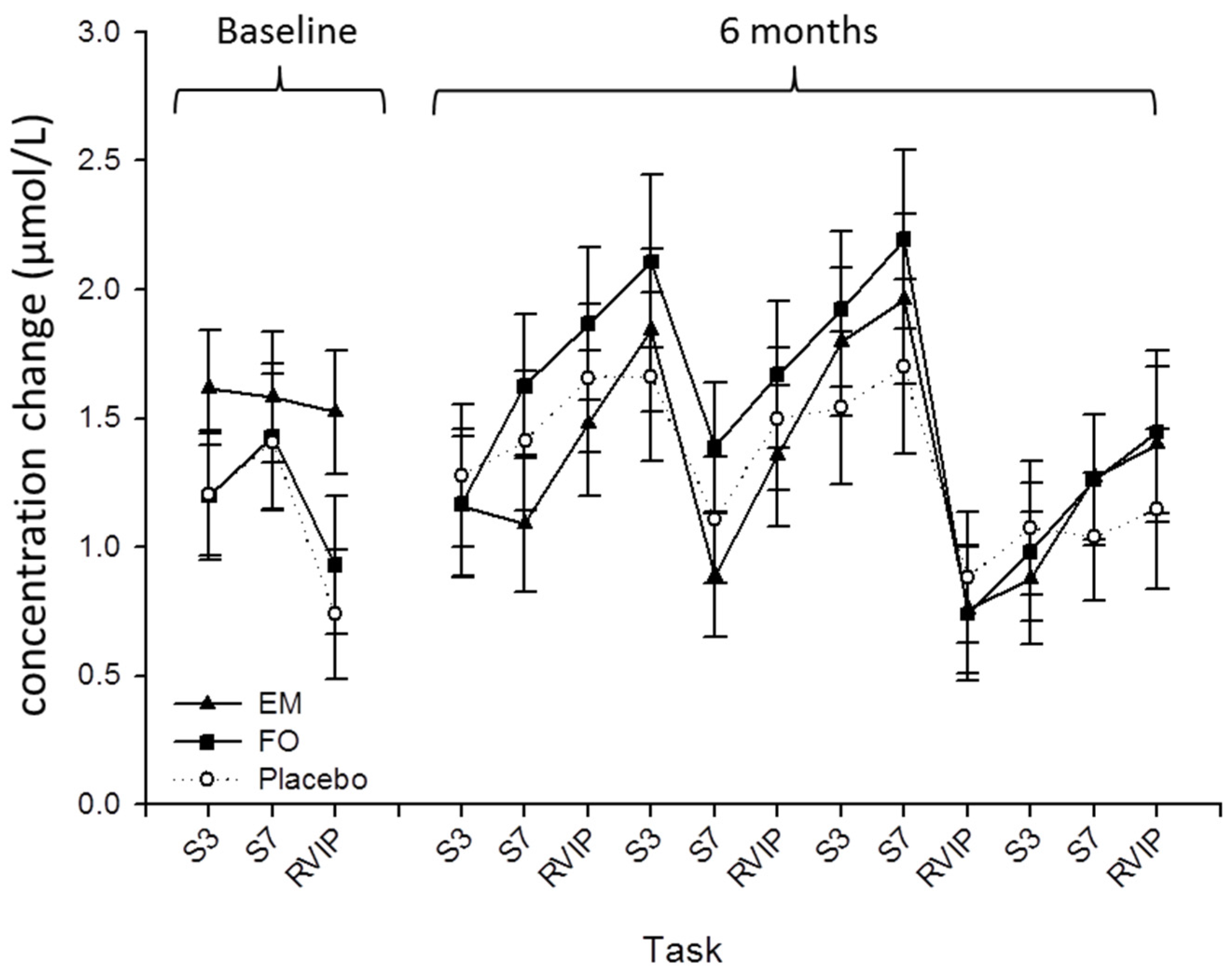
3. Results
3.1. Baseline Differences

{kind=link}
{kind=link}
| EM (n = 30) | FO (n = 26) | Placebo (n = 28) | p 1 | |
|---|---|---|---|---|
| Male/Female | 12/18 | 10/16 | 10/18 | 0.503 |
| Age (years) | 59.87 ± 4.70 2 | 60.31 ± 4.89 | 59.64 ± 5.28 | 0.883 |
| BMI (kg/m2) | 26.17 ± 3.44 | 25.65 ± 3.29 | 25.76 ± 4.25 | 0.857 |
| Blood pressure (systolic mmHg) | 120 ± 11 | 123 ± 14 | 124 ± 15 | 0.437 |
| Blood pressure (diastolic mmHg) | 81 ± 8 | 83 ± 8 | 84 ± 7 | 0.293 |
| Handedness (left/right) | 3/27 | 2/24 | 2/26 | 0.471 |
| Years in education | 13.30 ± 3.16 | 13.54 ± 3.39 | 14.86 ± 3.95 | 0.205 |
| Baseline | Rep 1 | Rep 2 | Rep 3 | Rep 4 | F | p | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |||||
| Serial 3 s total | EM | 26.67 | 1.81 | 28.51 | 0.75 | 32.53 | 0.75 | 32.35 | 0.92 | 32.03 | 0.86 | Treat | 0.04 | 0.96 |
| FO | 29.19 | 2.73 | 29.69 | 0.84 | 31.60 | 0.84 | 30.93 | 1.03 | 32.43 | 0.97 | Treat × rep | 1.21 | 0.31 | |
| Placebo | 28.54 | 2.24 | 28.53 | 0.79 | 31.70 | 0.80 | 31.60 | 0.98 | 32.47 | 0.91 | ||||
| Serial 3 s error | EM | 2.27 | 0.42 | 2.19 | 0.45 | 1.93 | 0.33 | 2.21 | 0.38 | 2.46 | 0.35 | Treat | 0.79 | 0.46 |
| FO | 2.73 | 0.59 | 2.14 | 0.52 | 2.31 | 0.37 | 1.88 | 0.43 | 2.24 | 0.39 | Treat × rep | 0.41 | 0.87 | |
| Placebo | 1.86 | 0.26 | 2.88 | 0.48 | 2.36 | 0.35 | 2.65 | 0.41 | 2.42 | 0.37 | ||||
| Serial 7 s total | EM | 19.20 | 1.41 | 21.61 | 0.66 | 22.12 | 0.68 | 21.87 | 0.72 | 22.35 | 0.77 | Treat | 0.49 | 0.61 |
| FO | 22.23 | 2.35 | 21.91 | 0.74 | 22.84 | 0.76 | 22.94 | 0.81 | 23.66 | 0.87 | Treat × rep | 0.58 | 0.727 1 | |
| Placebo | 21.21 | 1.83 | 21.22 | 0.70 | 21.80 | 0.72 | 22.80 | 0.76 | 22.62 | 0.82 | ||||
| Serial 7 s error | EM | 2.80 | 0.67 | 3.36 | 0.56 | 2.76 | 0.39 | 2.37 | 0.33 | 2.73 | 0.41 | Treat | 0.19 | 0.83 |
| FO | 3.12 | 0.47 | 2.73 | 0.62 | 2.92 | 0.44 | 2.53 | 0.37 | 3.10 | 0.46 | Treat × rep | 0.52 | 0.758 1 | |
| Placebo | 2.93 | 0.35 | 2.65 | 0.59 | 3.05 | 0.41 | 2.15 | 0.35 | 2.47 | 0.44 | ||||
| RVIP accuracy | EM | 51.58 | 4.50 | 50.41 | 3.90 | 50.21 | 4.36 | 48.83 | 4.71 | 50.39 | 4.74 | Treat | 0.44 | 0.65 |
| FO | 51.44 | 5.27 | 48.83 | 4.36 | 44.43 | 4.88 | 44.48 | 5.27 | 42.61 | 5.30 | Treat × rep | 0.54 | 0.760 1 | |
| Placebo | 50.71 | 5.30 | 48.62 | 4.13 | 48.34 | 4.62 | 45.73 | 4.99 | 40.98 | 5.03 | ||||
| RVIP RT | EM | 497.62 | 10.07 | 492.84 | 20.09 | 504.63 | 28.75 | 472.02 | 31.67 | 495.97 | 32.43 | Treat | 1.43 | 0.25 |
| FO | 473.45 | 21.44 | 462.49 | 22.42 | 431.84 | 32.10 | 437.02 | 35.35 | 404.81 | 36.20 | Treat × rep | 0.74 | 0.597 1 | |
| Placebo | 477.06 | 19.60 | 498.49 | 21.33 | 465.37 | 30.53 | 436.43 | 33.62 | 434.72 | 34.43 | ||||
| Mental fatigue | EM | 54.03 | 2.99 | 54.57 | 3.20 | 59.44 | 3.60 | 65.94 | 3.71 | 71.35 | 3.52 | Treat | 0.60 | 0.55 |
| FO | 58.15 | 4.24 | 57.06 | 3.59 | 66.98 | 4.03 | 72.21 | 4.16 | 76.32 | 3.95 | Treat × rep | 0.79 | 0.530 1 | |
| Placebo | 51.89 | 4.48 | 53.39 | 3.40 | 63.71 | 3.82 | 70.33 | 3.94 | 74.89 | 3.74 | ||||

3.2. Compliance
| EM (n = 30) | FO (n = 26) | Placebo (n = 28) | F | p | |||||
|---|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | ||||
| AA | Baseline | 9.41 | 1.21 | 9.47 | 1.28 | 10.08 | 1.23 | 14.99 | <0.001 |
| 6 months | 8.56 1 | 1.08 | 8.56 1 | 1.11 | 10.00 2,3 | 1.16 | |||
| Total n-6 PUFA | Baseline | 32.77 | 2.36 | 32.86 | 2.24 | 33.62 | 2.27 | 16.23 | <0.001 |
| 6 months | 30.79 1 | 2.34 | 31.32 1 | 2.24 | 33.73 2,3 | 2.07 | |||
| EPA | Baseline | 1.08 | 0.45 | 0.93 | 0.28 | 1.02 | 0.36 | 19.81 | <0.001 |
| 6 months | 1.69 1 | 0.72 | 1.42 1 | 0.49 | 0.98 2,3 | 0.32 | |||
| DHA | Baseline | 2.85 | 0.74 | 2.70 | 0.71 | 2.83 | 0.70 | 108.45 | <0.001 |
| 6 months | 5.41 1 | 1.11 | 5.56 1 | 0.85 | 2.80 2,3 | 0.63 | |||
| Total n-3 PUFA | Baseline | 5.91 | 1.12 | 5.58 | 0.89 | 5.75 | 1.03 | 74.25 | <0.001 |
| 6 months | 8.81 1 | 1.77 | 8.56 1 | 1.25 | 5.70 2,3 | 1.00 | |||
| AA:EPA | Baseline | 9.91 | 3.82 | 11.09 | 3.69 | 10.83 | 3.40 | 34.48 | <0.001 |
| 6 months | 6.05 1 | 3.07 | 6.60 1 | 2.08 | 11.12 2,3 | 3.54 | |||
| AA:DHA | Baseline | 3.55 | 1.16 | 3.78 | 1.20 | 3.74 | 0.86 | 122.25 | <0.001 |
| 6 months | 1.70 1 | 0.68 | 1.59 1 | 0.45 | 3.72 2,3 | 0.80 | |||
3.3. Near Infrared Spectroscopy
3.4. Cognitive Performance
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Colcombe, S.J.; Erickson, K.I.; Raz, N.; Webb, A.G.; Cohen, N.J.; McAuley, E.; Kramer, A.F. Aerobic fitness reduces brain tissue loss in aging humans. J. Gerontol. A Biol. Sci. Med. Sci. 2003, 58, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Barberger-Gateau, P.; Raffaitin, C.; Letenneur, L.; Berr, C.; Tzourio, C.; Dartigues, J.F.; Alperovitch, A. Dietary patterns and risk of dementia: The three-city cohort study. Neurology 2007, 69, 1921–1930. [Google Scholar] [CrossRef] [PubMed]
- Del Parigi, A.; Panza, F.; Capurso, C.; Solfrizzi, V. Nutritional factors, cognitive decline, and dementia. Brain Res. Bull. 2006, 69, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Kalmijn, S.; Launer, L.J.; Ott, A.; Witteman, J.C.; Hofman, A.; Breteler, M.M. Dietary fat intake and the risk of incident dementia in the rotterdam study. Ann. Neurol. 1997, 42, 776–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heude, B.; Ducimetiere, P.; Berr, C. Cognitive decline and fatty acid composition of erythrocyte membranes—The eva study. Am. J. Clin. Nutr. 2003, 77, 803–808. [Google Scholar] [PubMed]
- Dullemeijer, C.; Durga, J.; Brouwer, I.A.; van de Rest, O.V.; Kok, F.J.; Brummer, R.J.M.; van Boxtel, M.P.J.; Verhoef, P. N-3 fatty acid proportions in plasma and cognitive performance in older adults. Am. J. Clin. Nutr. 2007, 86, 1479–1485. [Google Scholar] [PubMed]
- Kalmijn, S.; van Boxtel, M.P.J.; Ocke, M.; Verschuren, W.M.M.; Kromhout, D.; Launer, L.J. Dietary intake of fatty acids and fish in relation to cognitive performance at middle age. Neurology 2004, 62, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Tangney, C.C.; Bienias, J.L.; Wilson, R.S. Fish consumption and cognitive decline with age in a large community study. Arch. Neurol. 2005, 62, 1849–1853. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.; Li, Q.; Chu, J.; Zeng, W.; Yang, M.; Zhu, S. Effect of n-3 pufa supplementation on cognitive function throughout the life span from infancy to old age: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2014, 100, 1422–1436. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.K.; Shahar, S.; Chin, A.V.; Yusoff, N.A. Docosahexaenoic acid-concentrated fish oil supplementation in subjects with mild cognitive impairment (MCI): A 12-month randomised, double-blind, placebo-controlled trial. Psychopharmacology 2013, 225, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Sinn, N.; Milte, C.M.; Street, S.J.; Buckley, J.D.; Coates, A.M.; Petkov, J.; Howe, P.R. Effects of n-3 fatty acids, EPA v. DHA, on depressive symptoms, quality of life, memory and executive function in older adults with mild cognitive impairment: A 6-month randomised controlled trial. Br. J. Nutr. 2012, 107, 1682–1693. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.V.; Kerti, L.; Hermannstadter, H.M.; Fiebach, J.B.; Schreiber, S.J.; Schuchardt, J.P.; Hahn, A.; Floel, A. Long-chain omega-3 fatty acids improve brain function and structure in older adults. Cereb. Cortex. 2014, 24, 3059–3068. [Google Scholar] [CrossRef] [PubMed]
- Yurko-Mauro, K.; McCarthy, D.; Rom, D.; Nelson, E.B.; Ryan, A.S.; Blackwell, A.; Salem, N., Jr.; Stedman, M. Beneficial effects of docosahexaenoic acid on cognition in age-related cognitive decline. Alzheimers Dement. 2010, 6, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Strike, S.C.; Carlisle, A.; Gibson, E.L.; Dyall, S.C. A high omega-3 fatty acid multinutrient supplement benefits cognition and mobility in older women: A randomized, double-blind, placebo-controlled pilot study. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 71, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Kevala, K.; Kim, J.; Moon, H.S.; Jun, S.B.; Lovinger, D.; Kim, H.Y. Docosahexaenoic acid promotes hippocampal neuronal development and synaptic function. J. Neurochem. 2009, 111, 510–521. [Google Scholar] [CrossRef] [PubMed]
- Denis, I.; Potier, B.; Vancassel, S.; Heberden, C.; Lavialle, M. Omega-3 fatty acids and brain resistance to ageing and stress: Body of evidence and possible mechanisms. Ageing Res. Rev. 2013, 12, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Pongrac, J.L.; Slack, P.J.; Innis, S.M. Dietary polyunsaturated fat that is low in (n-3) and high in (n-6) fatty acids alters the snare protein complex and nitrosylation in rat hippocampus. J. Nutr. 2007, 137, 1852–1856. [Google Scholar] [PubMed]
- Su, H.M. Mechanisms of n-3 fatty acid-mediated development and maintenance of learning memory performance. J. Nutr. Biochem. 2010, 21, 364–373. [Google Scholar] [CrossRef] [PubMed]
- Cutuli, D.; De Bartolo, P.; Caporali, P.; Laricchiuta, D.; Foti, F.; Ronci, M.; Rossi, C.; Neri, C.; Spalletta, G.; Caltagirone, C.; et al. N-3 polyunsaturated fatty acids supplementation enhances hippocampal functionality in aged mice. Front. Aging Neurosci. 2014, 6, 17. [Google Scholar] [CrossRef] [PubMed]
- Cysneiros, R.M.; Ferrari, D.; Arida, R.M.; Terra, V.C.; de Almeida, A.C.G.; Cavalheiro, E.A.; Scorza, F.A. Qualitative analysis of hippocampal plastic changes in rats with epilepsy supplemented with oral omega-3 fatty acids. Epilepsy Behav. 2010, 17, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Denis, I.; Potier, B.; Heberden, C.; Vancassel, S. Omega-3 polyunsaturated fatty acids and brain aging. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Salvati, S.; Natali, F.; Attorri, L.; Di Benedetto, R.; Leonardi, F.; Di Biase, A.; Ferri, F.; Fortuna, S.; Lorenzini, P.; Sanchez, M.; et al. Eicosapentaenonic add stimulates the expression of myelin proteins in rat brain. J. Neurosci. Res. 2008, 86, 776–784. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, Y.; Hashimoto, M.; Sugioka, K.; Maruyama, M.; Fujii, Y.; Hagiwara, R.; Hara, T.; Hossain, S.M.; Shido, O. Improvement of spatial cognition with dietary docosahexaenoic acid is associated with an increase in Fos expression in rat CA1 hippocampus. Clin. Exp. Pharmacol. Physiol. 2004, 31, 700–703. [Google Scholar] [CrossRef] [PubMed]
- Cansev, M.; Wurtman, R.J. Chronic administration of docosahexaenoic acid or eicosapentaenoic acid, but not arachidonic acid, alone or in combination with uridine, increases brain phosphatide and synaptic protein levels in gerbils. Neuroscience 2007, 148, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Aid, S.; Vancassel, S.; Poumes-Ballihaut, C.; Chalon, S.; Guesnet, P.; Lavialle, M. Effect of a diet-induced n-3 PUFA depletion on cholinergic parameters in the rat hippocampus. J. Lipid Res. 2003, 44, 1545–1551. [Google Scholar] [CrossRef] [PubMed]
- Gold, P.E. Acetylcholine modulation of neural systems involved in learning and memory. Neurobiol. Learn. Mem. 2003, 80, 194–210. [Google Scholar] [CrossRef] [PubMed]
- Griffith, W.H.; DuBois, D.W.; Fincher, A.; Peebles, K.A.; Bizon, J.L.; Murchison, D. Characterization of age-related changes in synaptic transmission onto f344 rat basal forebrain cholinergic neurons using a reduced synaptic preparation. J. Neurophysiol. 2014, 111, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Bartus, R.T. On neurodegenerative diseases, models, and treatment strategies: Lessons learned and lessons forgotten a generation following the cholinergic hypothesis. Exp. Neurol. 2000, 163, 495–529. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, H.; Sato, K.; Kakiuchi, T.; Nishiyama, S. Age-related impairment of coupling mechanism between neuronal activation and functional cerebral blood flow response was restored by cholinesterase inhibition: Pet study with microdialysis in the awake monkey brain. Brain Res. 2000, 857, 158–164. [Google Scholar] [CrossRef]
- Tsukada, H.; Kakiuchi, T.; Fukumoto, D.; Nishiyama, S.; Koga, K. Docosahexaenoic acid (DHA) improves the age-related impairment of the coupling mechanism between neuronal activation and functional cerebral blood flow response: A pet study in conscious monkeys. Brain Res. 2000, 862, 180–186. [Google Scholar] [CrossRef]
- Jackson, P.A.; Reay, J.L.; Scholey, A.B.; Kennedy, D.O. DHA-rich fish oil modulates the cerebral hemodynamic response to cognitive tasks in healthy young adults. Biol. Psychol. 2012, 89, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Pardini, M.; Serrati, C.; Guida, S.; Mattei, C.; Abate, L.; Massucco, D.; Sassos, D.; Amore, M.; Krueger, F.; Cocito, L.; et al. Souvenaid reduces behavioral deficits and improves social cognition skills in frontotemporal dementia: A proof-of-concept study. Neurodegener. Dis. 2015, 15, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; Twisk, J.W.; Blesa, R.; Scarpini, E.; von Arnim, C.A.; Bongers, A.; Harrison, J.; Swinkels, S.H.; Stam, C.J.; de Waal, H.; et al. Efficacy of souvenaid in mild alzheimer's disease: Results from a randomized, controlled trial. J. Alzheimers Dis. 2012, 31, 225–236. [Google Scholar] [PubMed]
- Kennedy, D.O.; Wightman, E.L.; Reay, J.L.; Lietz, G.; Okello, E.J.; Wilde, A.; Haskell, C.F. Effects of resveratrol on cerebral blood flow variables and cognitive performance in humans: A double-blind, placebo-controlled, crossover investigation. Am. J. Clin. Nutr. 2010, 91, 1590–1597. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Haskell-Ramsay, C.F.; Thompson, K.G.; Blackwell, J.R.; Winyard, P.G.; Forster, J.; Jones, A.M.; Kennedy, D.O. Dietary nitrate modulates cerebral blood flow parameters and cognitive performance in humans: A double-blind, placebo-controlled, crossover investigation. Physiol. Behav. 2015, 149, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Reay, J.L.; Haskell, C.F.; Williamson, G.; Dew, T.P.; Kennedy, D.O. Effects of resveratrol alone or in combination with piperine on cerebral blood flow parameters and cognitive performance in human subjects: A randomised, double-blind, placebo-controlled, cross-over investigation. Br. J. Nutr. 2014, 112, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Wightman, E.L.; Haskell, C.F.; Forster, J.S.; Veasey, R.C.; Kennedy, D.O. Epigallocatechin gallate, cerebral blood flow parameters, cognitive performance and mood in healthy humans: A double-blind, placebo-controlled, crossover investigation. Hum. Psychopharmacol. Clin. Exp. 2012, 27, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Haskell, C.F. Cerebral blood flow and behavioural effects of caffeine in habitual and non-habitual consumers of caffeine: A near infrared spectroscopy study. Biological. Psychol. 2011, 86, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Konagai, C.; Yanagimoto, K.; Hayamizu, K.; Han, L.; Tsuji, T.; Koga, Y. Effects of krill oil containing n-3 polyunsaturated fatty acids in phospholipid form on human brain function: A randomized controlled trial in healthy elderly volunteers. Clin. Interv. Aging 2013, 8, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Obrig, H.; Villringer, A. Beyond the visible—Imaging the human brain with light. J. Cereb. Blood Flow Metabo. 2003, 23, 1–18. [Google Scholar] [CrossRef]
- Bell, J.G.; Mackinlay, E.E.; Dick, J.R.; Younger, I.; Lands, B.; Gilhooly, T. Using a fingertip whole blood sample for rapid fatty acid measurement: Method validation and correlation with erythrocyte polar lipid compositions in UK subjects. Br. J. Nutr. 2011, 106, 1408–1415. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.A.; Reay, J.L.; Scholey, A.B.; Kennedy, D.O. Docosahexaenoic acid-rich fish oil modulates the cerebral hemodynamic response to cognitive tasks in healthy young adults. Biol. Psychol. 2012, 89, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Hossain, M.S.; Yamasaki, H.; Yazawa, K.; Masumura, S. Effects of eicosapentaenoic acid and docosahexaenoic acid on plasma membrane fluidity of aortic endothelial cells. Lipids 1999, 34, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Kim, S.W.; Li, X.L.; Datta, S.; Pond, W.G.; Wu, G.Y. Dietary supplementation with cholesterol and docosahexaenoic acid increases the activity of the arginine-nitric oxide pathway in tissues of young pigs. Nitric. Oxide. 2008, 19, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Kitaura, H.; Uozumi, N.; Tohmi, M.; Yamazaki, M.; Sakimura, K.; Kudoh, M.; Shimizu, T.; Shibuki, K. Roles of nitric oxide as a vasodilator in neurovascular coupling of mouse somatosensory cortex. Neurosci. Res. 2007, 59, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Bauer, I.; Crewther, D.P.; Pipingas, A.; Rowsell, R.; Cockerell, R.; Crewther, S.G. Omega-3 fatty acids modify human cortical visual processing-a double-blind, crossover study. PLoS ONE 2011, 6, e28214. [Google Scholar] [CrossRef] [PubMed]
- Bauer, I.; Hughes, M.; Rowsell, R.; Cockerell, R.; Pipingas, A.; Crewther, S.; Crewther, D. Omega-3 supplementation improves cognition and modifies brain activation in young adults. Hum. Psychopharmaco. 2014, 29, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Jacka, F.N.; Pasco, J.A.; Henry, M.J.; Kotowicz, M.A.; Nicholson, G.C.; Berk, M. Dietary omega-3 fatty acids and depression in a community sample. Nutr. Neurosci. 2004, 7, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.A.; Deary, M.; Reay, J.L.; Scholey, A.B.; Kennedy, D.O. Twelve weeks' dietary supplementation with 1 g fish oil does not improve cognitive performance or mood in healthy young adults. Br. J. Nutr. 2012, 107, 1232–1243. [Google Scholar] [CrossRef] [PubMed]
- Jackson, P.A.; Reay, J.L.; Scholey, A.B.; Kennedy, D.O. DHA-rich oil modulates the cerebral haemodynamic response to cognitive tasks in healthy young adults: A near IR spectroscopy pilot study. Br. J. Nutr. 2012, 107, 1093–1098. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Giltay, E.J.; Kromhout, D. Effects of n-3 fatty acids on cognitive decline: A randomized, double-blind, placebo-controlled trial in stable myocardial infarction patients. Alzheimers. Dement. 2012, 8, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Dangour, A.D.; Allen, E.; Elbourne, D.; Fasey, N.; Fletcher, A.E.; Hardy, P.; Holder, G.E.; Knight, R.; Letley, L.; Richards, M.; et al. Effect of 2-y n-3 long-chain polyunsaturated fatty acid supplementation on cognitive function in older people: A randomized, double-blind, controlled trial. Am. J. Clin. Nutr. 2010, 91, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- van de Rest, O.; Geleijnse, J.M.; Kok, F.J.; van Staveren, W.A.; Dullemeijer, C.; Olderikkert, M.G.; Beekman, A.T.; de Groot, C.P. Effect of fish oil on cognitive performance in older subjects: A randomized, controlled trial. Neurology 2008, 71, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Yurko-Mauro, K.; Alexander, D.D.; Van Elswyk, M.E. Docosahexaenoic acid and adult memory: A systematic review and meta-analysis. PLoS ONE 2015, 10, 18. [Google Scholar]
- Kennedy, D.O.; Scholey, A.B.; Wesnes, K.A. Modulation of cognition and mood following administration of single doses of ginkgo biloba, ginseng, and a ginkgo/ginseng combination to healthy young adults. Physiol. Behav. 2002, 75, 739–751. [Google Scholar] [CrossRef]
- Mix, J.A.; Crews, W.D. A double-blind, placebo-controlled, randomized trial of ginkgo biloba extract EGb 761 in a sample of cognitively intact older adults: Neuropsychological findings. Hum. Psychopharmacol. 2002, 17, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Burns, N.R.; Bryan, J.; Nettelbeck, T. Ginkgo biloba: No robust effect on cognitive abilities or mood in healthy young or older adults. Hum. Psychopharmacol. 2006, 21, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Snitz, B.E.; O'Meara, E.S.; Carlson, M.C.; Arnold, A.M.; Ives, D.G.; Rapp, S.R.; Saxton, J.; Lopez, O.L.; Dunn, L.O.; Sink, K.M.; et al. Ginkgo biloba for preventing cognitive decline in older adults a randomized trial. JAMA 2009, 302, 2663–2670. [Google Scholar] [CrossRef] [PubMed]
- Solomon, P.R.; Adams, F.; Silver, A.; Zimmer, J.; DeVeaux, R. Ginkgo for memory enhancement—A randomized controlled trial. JAMA 2002, 288, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Van de Rest, O.; van Hooijdonk, L.W.A.; Doets, E.; Schiepers, O.J.G.; Eilander, A.; de Groot, L. B vitamins and n-3 fatty acids for brain development and function: Review of human studies. Ann. Nutr. Metab. 2012, 60, 272–292. [Google Scholar] [CrossRef] [PubMed]
- Van der Zwaluw, N.L.; Dhonukshe-Rutten, R.A.M.; van Wijngaarden, J.P.; Brouwer-Brolsma, E.M.; van de Rest, O.; In’t Veld, P.H.; Enneman, A.W.; van Dijk, S.C.; Ham, A.C.; Swart, K.M.A.; et al. Results of 2-year vitamin B treatment on cognitive performance secondary data from an RCT. Neurology 2014, 83, 2158–2166. [Google Scholar] [CrossRef] [PubMed]
- Barberger-Gateau, P. Nutrition and brain aging: How can we move ahead? Eur. J. Clin. Nutr. 2014, 68, 1245–1249. [Google Scholar] [CrossRef] [PubMed]
- Vakhapova, V.; Cohen, T.; Richter, Y.; Herzog, Y.; Kam, Y.; Korczyn, A.D. Phosphatidylserine containing omega-3 fatty acids may improve memory abilities in nondemented elderly individuals with memory complaints: Results from an open-label extension study. Dement. Geriatri. Cogn. Disord. 2014, 38, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Vakhapova, V.; Cohen, T.; Richter, Y.; Herzog, Y.; Korczyn, A.D. Phosphatidylserine containing ω-3 fatty acids may improve memory abilities in non-demented elderly with memory complaints: A double-blind placebo-controlled trial. Dement. Geriatri. Cogn. Disord. 2010, 29, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Marangoni, F.; Colombo, C.; Galli, C. A method for the direct evaluation of the fatty acid status in a drop of blood from a fingertip in humans: Applicability to nutritional and epidemiological studies. Anal. Biochem. 2004, 326, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Bailey-Hall, E.; Nelson, E.B.; Ryan, A.S. Validation of a rapid measure of blood PUFA levels in humans. Lipids 2008, 43, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzi, V.; Frisardi, V.; Capurso, C.; D’Introno, A.; Colacicco, A.M.; Vendemiale, G.; Capurso, A.; Panza, F. Dietary fatty acids in dementia and predementia syndromes: Epidemiological evidence and possible underlying mechanisms. Ageing Res. Rev. 2010, 9, 184–199. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.E.; Tye, C.; Kuntsi, J.; Vassos, E.; Asherson, P. Omega-3 polyunsaturated fatty acid supplementation and cognition: A systematic review and meta-analysis. J. Psychopharmacol. 2015, 29, 753–763. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackson, P.A.; Forster, J.S.; Bell, J.G.; Dick, J.R.; Younger, I.; Kennedy, D.O. DHA Supplementation Alone or in Combination with Other Nutrients Does not Modulate Cerebral Hemodynamics or Cognitive Function in Healthy Older Adults. Nutrients 2016, 8, 86. https://doi.org/10.3390/nu8020086
Jackson PA, Forster JS, Bell JG, Dick JR, Younger I, Kennedy DO. DHA Supplementation Alone or in Combination with Other Nutrients Does not Modulate Cerebral Hemodynamics or Cognitive Function in Healthy Older Adults. Nutrients. 2016; 8(2):86. https://doi.org/10.3390/nu8020086
Chicago/Turabian StyleJackson, Philippa A., Joanne S. Forster, J. Gordon Bell, James R. Dick, Irene Younger, and David O. Kennedy. 2016. "DHA Supplementation Alone or in Combination with Other Nutrients Does not Modulate Cerebral Hemodynamics or Cognitive Function in Healthy Older Adults" Nutrients 8, no. 2: 86. https://doi.org/10.3390/nu8020086
APA StyleJackson, P. A., Forster, J. S., Bell, J. G., Dick, J. R., Younger, I., & Kennedy, D. O. (2016). DHA Supplementation Alone or in Combination with Other Nutrients Does not Modulate Cerebral Hemodynamics or Cognitive Function in Healthy Older Adults. Nutrients, 8(2), 86. https://doi.org/10.3390/nu8020086