Therapeutic Versatility of Resveratrol Derivatives


Abstract
1. Introduction
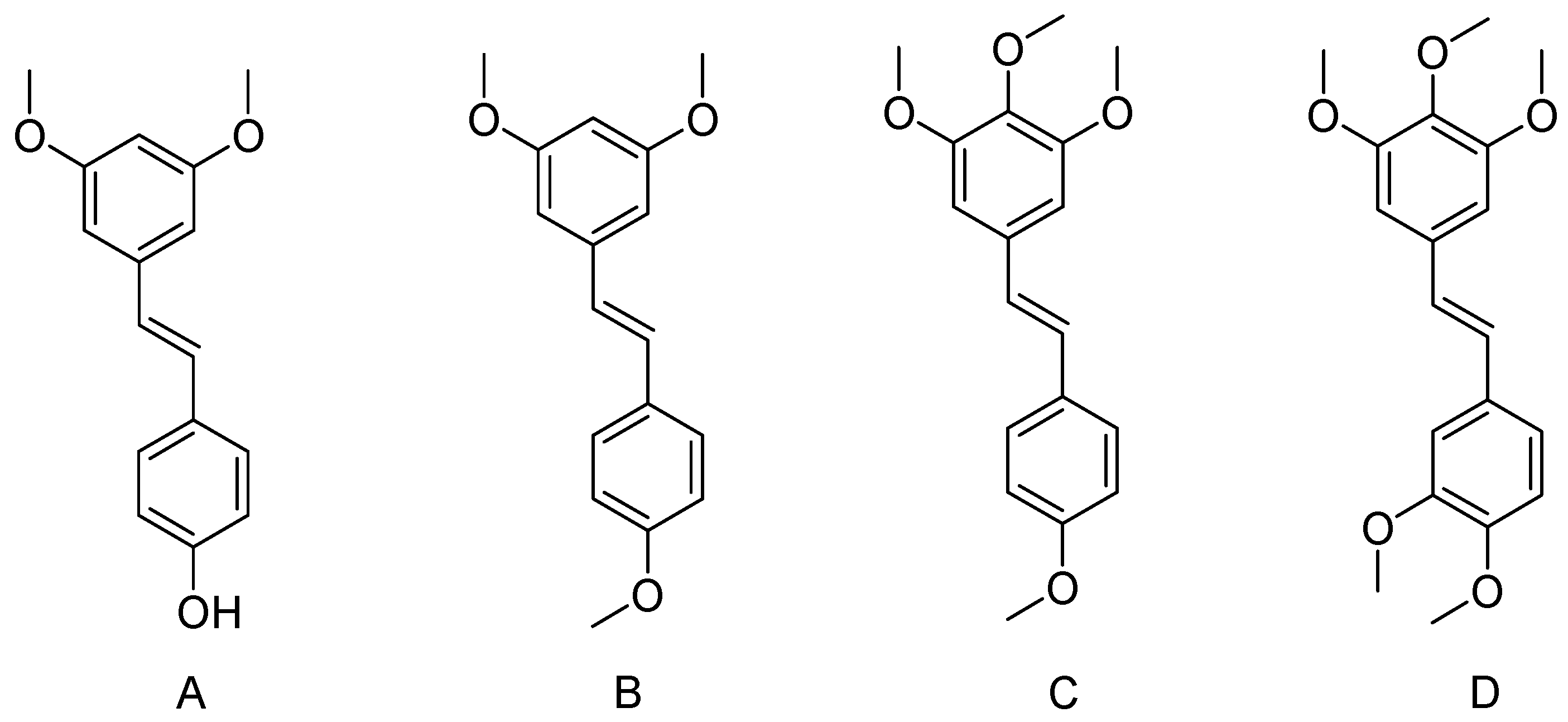
2. Methoxylated Derivatives of Resveratrol
2.1. Pterostilbene (3,5-Dimethoxy-4′ Hydroxystilbene)
2.1.1. Antioxidant Activity
2.1.2. Chemotherapeutic Activity
2.1.3. Cardioprotective Activity
2.1.4. Neuroprotective Activity
2.1.5. Antidiabetics
2.2. Trimethoxystilbene
2.2.1. Chemotherapeutic Activity
2.2.2. Cardioprotective Activity
2.2.3. Anti-HCV Activity
2.3. Tetramethoxystilbene
2.3.1. Chemotherapeutic Activity
2.3.2. Effect on Activity of Cytochromes P450
2.3.3. Cardioprotective Activity
2.4. Pentamethoxystilbene
2.4.1. Chemotherapeutic Activity
2.4.2. Cytochrome P-450 Inhibitor
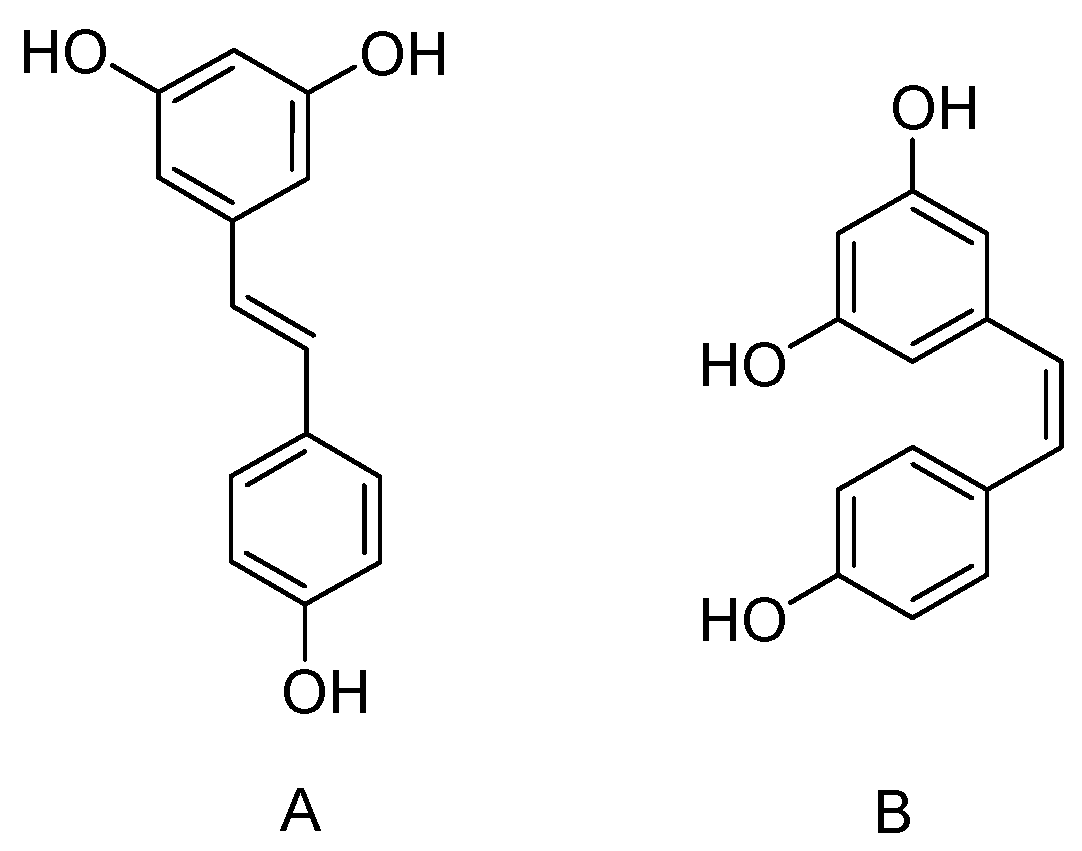
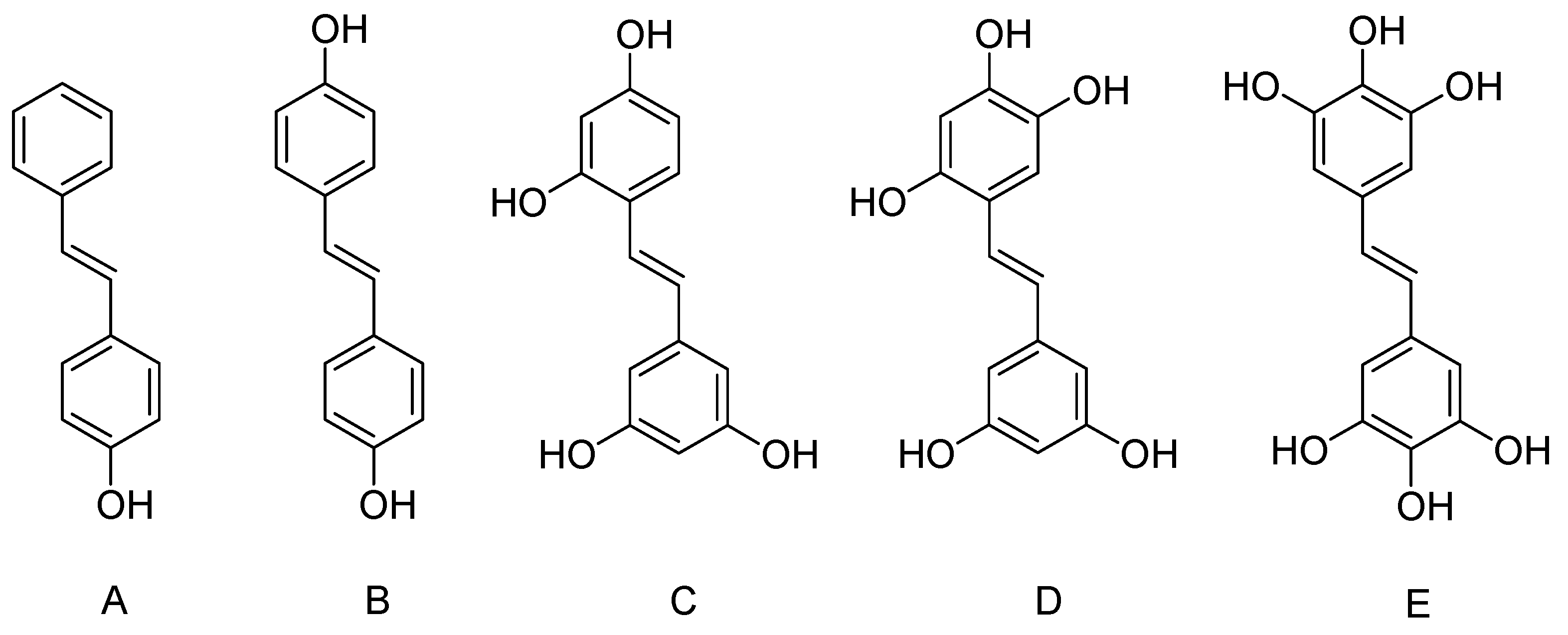
3. Hydroxylated Resveratrol Derivatives
3.1. Dihydroxystilbene
3.1.1. Chemotherapeutic Activity
3.1.2. Cardioprotective Activity
3.2. Tetrahydroxystilbene
3.2.1. Cardioprotective Activity
3.2.2. Chemotherapeutic Activity
3.2.3. Antimicrobial Activity
3.2.4. Neuroprotective Activity
3.3. Hexahydroxystilbene
3.3.1. Chemotherapeutic Activity
3.3.2. Anti-HIV
3.3.3. Selective COX-2 Inhibitor
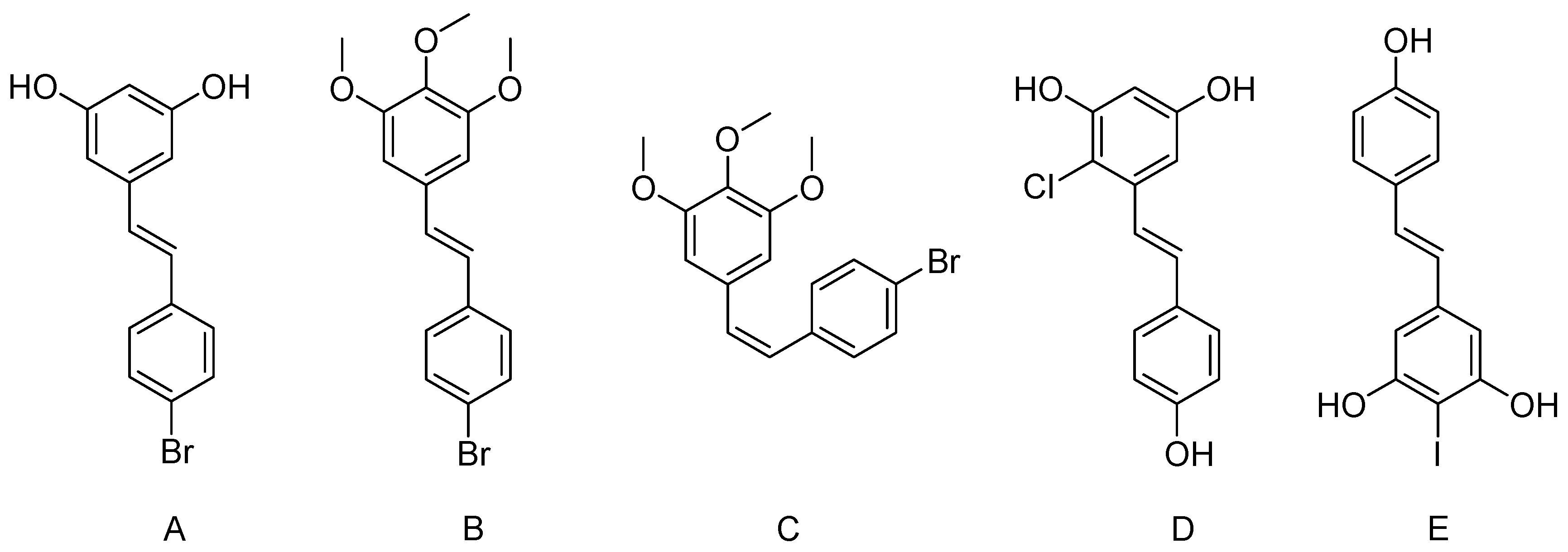
4. Halogenated Derivatives of Resveratrol
4.1. Chemotherapeutic Activity
4.2. Cardioprotective Activity
4.3. Free Radical Scavenging Activities
4.4. Antimicrobial Activity
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Biasutto, L.; Mattarei, A.; Azzolini, M.; La Spina, M.; Sassi, N.; Romio, M.; Paradisi, C.; Zoratti, M. Resveratrol derivatives as a pharmacological tool. Ann. N. Y. Acad. Sci. 2017, 1403, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Kursvietiene, L.; Staneviciene, I.; Mongirdiene, A.; Bernatoniene, J. Multiplicity of effects and health benefits of resveratrol. Medicina 2016, 52, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.; Crozier, A. Plant foods and herbal sources of resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef] [PubMed]
- Weiskirchen, S.; Weiskirchen, R. Resveratrol: How much wine do you have to drink to stay healthy? Adv. Nutr. 2016, 7, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Francioso, A.; Mastromarino, P.; Masci, A.; d’Erme, M.; Mosca, L. Chemistry, stability and bioavailability of resveratrol. Med. Chem. 2014, 10, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, S.S.; Canto, C. The molecular targets of resveratrol. Biochim. Biophys. Acta 2015, 1852, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Smoliga, J.M.; Baur, J.A.; Hausenblas, H.A. Resveratrol and health—A comprehensive review of human clinical trials. Mol. Nutr. Food Res. 2011, 55, 1129–1141. [Google Scholar] [CrossRef] [PubMed]
- Park, E.J.; Pezzuto, J.M. The pharmacology of resveratrol in animals and humans. Biochim. Biophys. Acta 2015, 1852, 1071–1113. [Google Scholar] [CrossRef] [PubMed]
- Wicklow, B.; Wittmeier, K.; GW, T.J.; McGavock, J.; Robert, M.; Duhamel, T.; Dolinsky, V.W. Proposed trial: Safety and efficacy of resveratrol for the treatment of non-alcoholic fatty liver disease (NAFLD) and associated insulin resistance in adolescents who are overweight or obese adolescents—Rationale and protocol. Biochem. Cell Boil. 2015, 93, 522–530. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.R.; Scott, E.; Brown, V.A.; Gescher, A.J.; Steward, W.P.; Brown, K. Clinical trials of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Walle, T. Bioavailability of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, C.; Li, H.; Wu, M.; Ren, C.; Zhen, Y.; Ma, X.; Diao, Y.; Ma, X.; Deng, S.; et al. Differential sensitivities of bladder cancer cell lines to resveratol are unrelated to its metabolic profile. Oncotarget. 2017, 8, 40289–40304. [Google Scholar] [CrossRef] [PubMed]
- Bitterman, J.L.; Chung, J.H. Metabolic effects of resveratrol: Addressing the controversies. Cell. Mol. Life Sci. 2015, 72, 1473–1488. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Wang, L.; Zhu, M.; Zhang, L.; Yan, L. Properties and molecular mechanisms of resveratrol: A review. Die Pharm. 2015, 70, 501–506. [Google Scholar]
- Szekeres, T.; Fritzer-Szekeres, M.; Saiko, P.; Jager, W. Resveratrol and resveratrol analogues—Structure-activity relationship. Pharm. Res. 2010, 27, 1042–1048. [Google Scholar] [CrossRef] [PubMed]
- Delmas, D.; Aires, V.; Limagne, E.; Dutartre, P.; Mazue, F.; Ghiringhelli, F.; Latruffe, N. Transport, stability, and biological activity of resveratrol. Ann. N. Y. Acad. Sci. 2011, 1215, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Mikula-Pietrasik, J.; Kuczmarska, A.; Ksiazek, K. Biological multifunctionality of resveratrol and its derivatives. Postepy Biochem. 2015, 61, 336–343. [Google Scholar] [PubMed]
- Pan, M.H.; Gao, J.H.; Lai, C.S.; Wang, Y.J.; Chen, W.M.; Lo, C.Y.; Wang, M.; Dushenkov, S.; Ho, C.T. Antitumor activity of 3,5,4′-trimethoxystilbene in colo 205 cells and xenografts in SCID mice. Mol. Carcinog. 2008, 47, 184–196. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, G.A.; Savio, M.; Forti, L.; Shevelev, I.; Ramadan, K.; Stivala, L.A.; Vannini, V.; Hubscher, U.; Spadari, S.; Maga, G. Inhibition of mammalian DNA polymerases by resveratrol: Mechanism and structural determinants. Biochem. J. 2005, 389, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Das, J.; Pany, S.; Majhi, A. Chemical modifications of resveratrol for improved protein kinase c alpha activity. Bioorg. Med. Chem. 2011, 19, 5321–5333. [Google Scholar] [CrossRef] [PubMed]
- Estrela, J.M.; Ortega, A.; Mena, S.; Rodriguez, M.L.; Asensi, M. Pterostilbene: Biomedical applications. Crit. Rev. Clin. Lab. Sci. 2013, 50, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Murias, M.; Jager, W.; Handler, N.; Erker, T.; Horvath, Z.; Szekeres, T.; Nohl, H.; Gille, L. Antioxidant, prooxidant and cytotoxic activity of hydroxylated resveratrol analogues: Structure-activity relationship. Biochem. Pharm. 2005, 69, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Savio, M.; Coppa, T.; Bianchi, L.; Vannini, V.; Maga, G.; Forti, L.; Cazzalini, O.; Lazze, M.C.; Perucca, P.; Prosperi, E.; et al. The resveratrol analogue 4,4′-dihydroxy-trans-stilbene inhibits cell proliferation with higher efficiency but different mechanism from resveratrol. Int. J. Biochem. Cell Boil. 2009, 41, 2493–2502. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Wei, X.; Zhang, C.J.; Jin, X.L.; Tang, J.J.; Fan, G.J.; Zhou, B. Hypohalous acid-mediated halogenation of resveratrol and its role in antioxidant and antimicrobial activities. Food Chem. 2012, 135, 1239–1244. [Google Scholar] [CrossRef] [PubMed]
- Kosuru, R.; Rai, U.; Prakash, S.; Singh, A.; Singh, S. Promising therapeutic potential of pterostilbene and its mechanistic insight based on preclinical evidence. Eur. J. Pharm. 2016, 789, 229–243. [Google Scholar] [CrossRef] [PubMed]
- Remsberg, C.M.; Yanez, J.A.; Ohgami, Y.; Vega-Villa, K.R.; Rimando, A.M.; Davies, N.M. Pharmacometrics of pterostilbene: Preclinical pharmacokinetics and metabolism, anticancer, antiinflammatory, antioxidant and analgesic activity. Phytother. Res. PTR 2008, 22, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Yue, B.D.; Ho, P.C. Determination of pterostilbene in rat plasma by a simple HPLC-UV method and its application in pre-clinical pharmacokinetic study. Biomed. Chromatogr. 2009, 23, 1308–1315. [Google Scholar] [CrossRef] [PubMed]
- Rimando, A.M.; Cuendet, M.; Desmarchelier, C.; Mehta, R.G.; Pezzuto, J.M.; Duke, S.O. Cancer chemopreventive and antioxidant activities of pterostilbene, a naturally occurring analogue of resveratrol. J. Agric. Food Chem. 2002, 50, 3453–3457. [Google Scholar] [CrossRef] [PubMed]
- Hasiah, A.H.; Ghazali, A.R.; Weber, J.F.; Velu, S.; Thomas, N.F.; Inayat Hussain, S.H. Cytotoxic and antioxidant effects of methoxylated stilbene analogues on HepG2 hepatoma and chang liver cells: Implications for structure activity relationship. Hum. Exp. Toxicol. 2011, 30, 138–144. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.; McFadden, D. A review of pterostilbene antioxidant activity and disease modification. Oxidative Med. Cell. Longev. 2013, 2013, 575482. [Google Scholar] [CrossRef] [PubMed]
- McCormack, D.; McFadden, D. Pterostilbene and cancer: Current review. J. Surg. Res. 2012, 173, e53–e61. [Google Scholar] [CrossRef] [PubMed]
- Wawszczyk, J.; Kapral, M.; Hollek, A.; Weglarz, L. In vitro evaluation of antiproliferative and cytotoxic properties of pterostilbene against human colon cancer cells. Acta Pol. Pharm. 2014, 71, 1051–1055. [Google Scholar] [PubMed]
- Pan, M.H.; Chiou, Y.S.; Chen, W.J.; Wang, J.M.; Badmaev, V.; Ho, C.T. Pterostilbene inhibited tumor invasion via suppressing multiple signal transduction pathways in human hepatocellular carcinoma cells. Carcinogenesis 2009, 30, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.S.; Ho, C.T.; Tu, S.H.; Pan, M.H.; Chuang, C.H.; Chang, H.W.; Chang, C.H.; Wu, C.H.; Ho, Y.S. Long-term ethanol exposure-induced hepatocellular carcinoma cell migration and invasion through lysyl oxidase activation are attenuated by combined treatment with pterostilbene and curcumin analogues. J. Agric. Food Chem. 2013, 61, 4326–4335. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Chang, Y.H.; Badmaev, V.; Nagabhushanam, K.; Ho, C.T. Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. J. Agric. Food Chem. 2007, 55, 7777–7785. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Hong, B.H.; Ho, C.T.; Yen, G.C. Targeting cancer stem cells in breast cancer: Potential anticancer properties of 6-shogaol and pterostilbene. J. Agric. Food Chem. 2015, 63, 2432–2441. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, P.; Asensi, M.; Segarra, R.; Ortega, A.; Benlloch, M.; Obrador, E.; Varea, M.T.; Asensio, G.; Jorda, L.; Estrela, J.M. Association between pterostilbene and quercetin inhibits metastatic activity of B16 melanoma. Neoplasia 2005, 7, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Rimando, A.M.; Levenson, A.S. Resveratrol and pterostilbene as a microrna-mediated chemopreventive and therapeutic strategy in prostate cancer. Ann. N. Y. Acad. Sci. 2017, 1403, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Acharya, J.D.; Ghaskadbi, S.S. Protective effect of pterostilbene against free radical mediated oxidative damage. BMC Complement. Altern. Med. 2013, 13, 238. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhou, G.; Song, W.; Tan, X.; Guo, Y.; Zhou, B.; Jing, H.; Zhao, S.; Chen, L. Pterostilbene protects vascular endothelial cells against oxidized low-density lipoprotein-induced apoptosis in vitro and in vivo. Apoptosis Int. J. Program. Cell Death 2012, 17, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Zhang, L.; Li, F.; Hu, C.P.; Zhang, Z. Restoration of sirt1 function by pterostilbene attenuates hypoxia-reoxygenation injury in cardiomyocytes. Eur. J. Pharmacol. 2016, 776, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Magyar, K.; Halmosi, R.; Palfi, A.; Feher, G.; Czopf, L.; Fulop, A.; Battyany, I.; Sumegi, B.; Toth, K.; Szabados, E. Cardioprotection by resveratrol: A human clinical trial in patients with stable coronary artery disease. Clin. Hemorheol. Microcirc. 2012, 50, 179–187. [Google Scholar] [PubMed]
- Kasiotis, K.M.; Pratsinis, H.; Kletsas, D.; Haroutounian, S.A. Resveratrol and related stilbenes: Their anti-aging and anti-angiogenic properties. Food Chem. Toxicol. 2013, 61, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Rimando, A.; Pallas, M.; Camins, A.; Porquet, D.; Reeves, J.; Shukitt-Hale, B.; Smith, M.A.; Joseph, J.A.; Casadesus, G. Low-dose pterostilbene, but not resveratrol, is a potent neuromodulator in aging and alzheimer’s disease. Neurobiol. Aging 2012, 33, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Saw, C.L.; Guo, Y.; Yang, A.Y.; Paredes-Gonzalez, X.; Ramirez, C.; Pung, D.; Kong, A.N. The berry constituents quercetin, kaempferol, and pterostilbene synergistically attenuate reactive oxygen species: Involvement of the Nrf2-ARE signaling pathway. Food Chem. Toxicol. 2014, 72, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Thangthaeng, N.; Miller, M.G.; Shukitt-Hale, B. Effects of pterostilbene and resveratrol on brain and behavior. Neurochem. Int. 2015, 89, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.A.; Fisher, D.R.; Cheng, V.; Rimando, A.M.; Shukitt-Hale, B. Cellular and behavioral effects of stilbene resveratrol analogues: Implications for reducing the deleterious effects of aging. J. Agric. Food Chem. 2008, 56, 10544–10551. [Google Scholar] [CrossRef] [PubMed]
- Amarnath Satheesh, M.; Pari, L. The antioxidant role of pterostilbene in streptozotocin-nicotinamide-induced type 2 diabetes mellitus in wistar rats. J. Pharm. Pharmacol. 2006, 58, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Rimando, A.M.; Nagmani, R.; Feller, D.R.; Yokoyama, W. Pterostilbene, a new agonist for the peroxisome proliferator-activated receptor alpha-isoform, lowers plasma lipoproteins and cholesterol in hypercholesterolemic hamsters. J. Agric. Food Chem. 2005, 53, 3403–3407. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Zhang, W.; Go, M.L.; Choo, Q.Y.; Ho, P.C. Determination of Z-3,5,4′-trimethoxystilbene in rat plasma by a simple HPLC method: Application in a pre-clinical pharmacokinetic study. J. Pharm. Biomed. Anal. 2010, 53, 693–697. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Ho, P.C. A rapid HPLC method for the quantification of 3,5,4′-trimethoxy-trans-stilbene (TMS) in rat plasma and its application in pharmacokinetic study. J. Pharm. Biomed. Anal. 2009, 49, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Hsu, L.S.; Lin, C.L.; Hong, H.M.; Pan, M.H.; Way, T.D.; Chen, W.J. 3,5,4′-trimethoxystilbene, a natural methoxylated analog of resveratrol, inhibits breast cancer cell invasiveness by downregulation of PI3K/Akt and Wnt/beta-catenin signaling cascades and reversal of epithelial-mesenchymal transition. Toxicol. Appl. Pharmacol. 2013, 272, 746–756. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.J.; Yang, Y.T.; Ho, C.T.; Yen, G.C. Mechanisms of apoptotic effects induced by resveratrol, dibenzoylmethane, and their analogues on human lung carcinoma cells. J. Agric. Food Chem. 2009, 57, 5235–5243. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Weng, C.J.; Ho, C.T.; Yen, G.C. Resveratrol analog-3,5,4′-trimethoxy-trans-stilbene inhibits invasion of human lung adenocarcinoma cells by suppressing the MAPK pathway and decreasing matrix metalloproteinase-2 expression. Mol. Nutr. Food Res. 2009, 53, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Guo, X.; Chen, H.; Lin, T.; Xu, Y.; Chen, Q.; Liu, J.; Zeng, J.; Zhang, X.K.; Yao, X. A resveratrol analog, phoyunbene b, induces G2/M cell cycle arrest and apoptosis in HepG2 liver cancer cells. Bioorg. Med. Chem. Lett. 2012, 22, 2114–2118. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Luo, X.J.; Yang, Z.B.; Zhang, J.J.; Li, T.B.; Zhang, X.J.; Ma, Q.L.; Zhang, G.G.; Hu, C.P.; Peng, J. Inhibition of NOX/VPO1 pathway and inflammatory reaction by trimethoxystilbene in prevention of cardiovascular remodeling in hypoxia-induced pulmonary hypertensive rats. J. Cardiovasc. Pharmacol. 2014, 63, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Wang, X.; Qin, X.; Jiang, X.; Xiang, D.; Xie, L.; Hu, J.; Gao, J. Effects of trimethoxystilbene on proliferation and apoptosis of pulmonary artery smooth muscle cells. Cell Biochem. Biophys. 2012, 64, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dai, F.; Cui, L.; Jing, H.; Fan, P.; Tan, X.; Guo, Y.; Zhou, G. Novel role for TRPC4 in regulation of macroautophagy by a small molecule in vascular endothelial cells. Biochim. Biophys. Acta 2015, 1853, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Alex, D.; Leong, E.C.; Zhang, Z.J.; Yan, G.T.; Cheng, S.H.; Leong, C.W.; Li, Z.H.; Lam, K.H.; Chan, S.W.; Lee, S.M. Resveratrol derivative, trans-3,5,4′-trimethoxystilbene, exerts antiangiogenic and vascular-disrupting effects in zebrafish through the downregulation of VEGFR2 and cell-cycle modulation. J. Cell. Biochem. 2010, 109, 339–346. [Google Scholar] [PubMed]
- Belleri, M.; Ribatti, D.; Nicoli, S.; Cotelli, F.; Forti, L.; Vannini, V.; Stivala, L.A.; Presta, M. Antiangiogenic and vascular-targeting activity of the microtubule-destabilizing trans-resveratrol derivative 3,5,4′-trimethoxystilbene. Mol. Pharmacol. 2005, 67, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.B.; Kotturi, H.; Waris, G.; Mohammed, A.; Chandrakesan, P.; May, R.; Sureban, S.; Weygant, N.; Qu, D.; Rao, C.V.; et al. (Z)-3,5,4′-trimethoxystilbene limits hepatitis c and cancer pathophysiology by blocking microtubule dynamics and cell-cycle progression. Cancer Res. 2016, 76, 4887–4896. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Molavi, O.; Haddadi, A.; Lai, R.; Gossage, R.A.; Lavasanifar, A. Resveratrol analog trans 3,4,5,4'-tetramethoxystilbene (DMU-212) mediates anti-tumor effects via mechanism different from that of resveratrol. Cancer Chemother. Pharmacol. 2008, 63, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Zhang, W.; Go, M.L.; Tringali, C.; Spatafora, C.; Ho, P.C. Quantification of trans-3,4,5,4′-tetramethoxystilbene in rat plasma by HPLC: Application to pharmacokinetic study. J. Agric. Food Chem. 2011, 59, 1072–1077. [Google Scholar] [CrossRef] [PubMed]
- Sale, S.; Verschoyle, R.D.; Boocock, D.; Jones, D.J.L.; Wilsher, N.; Ruparelia, K.C.; Potter, G.A.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Pharmacokinetics in mice and growth-inhibitory properties of the putative cancer chemopreventive agent resveratrol and the synthetic analogue trans 3,4,5,4′-tetramethoxystilbene. Br. J. Cancer 2004, 90, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Androutsopoulos, V.P.; Fragiadaki, I.; Spandidos, D.A.; Tosca, A. The resveratrol analogue, 3,4,5,4′-trans-tetramethoxystilbene, inhibits the growth of a375 melanoma cells through multiple anticancer modes of action. Int. J. Oncol. 2016, 49, 1305–1314. [Google Scholar] [CrossRef] [PubMed]
- Androutsopoulos, V.P.; Fragiadaki, I.; Tosca, A. Activation of ERK1/2 is required for the antimitotic activity of the resveratrol analogue 3,4,5,4′-tetramethoxystilbene (DMU-212) in human melanoma cells. Exp. Dermatol. 2015, 24, 632–634. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.K.; Qiang, P.F.; Xu, Q.P.; Zhao, Y.H.; Dai, F.; Zhang, L. Trans-3,4,5,4′-tetramethoxystilbene, a resveratrol analog, potently inhibits angiogenesis in vitro and in vivo. Acta Pharmacol. Sin. 2013, 34, 1174–1182. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, H.; Myszkowski, K.; Abraszek, J.; Kwiatkowska-Borowczyk, E.; Amarowicz, R.; Murias, M.; Wierzchowski, M.; Jodynis-Liebert, J. Dmu-212 inhibits tumor growth in xenograft model of human ovarian cancer. Biomed. Pharmacother. 2014, 68, 397–400. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Park, N.; Chun, Y.J. Role of annexin a5 on mitochondria-dependent apoptosis induced by tetramethoxystilbene in human breast cancer cells. Biomol. Ther. 2014, 22, 519–524. [Google Scholar] [CrossRef] [PubMed]
- Cichocki, M.; Baer-Dubowska, W.; Wierzchowski, M.; Murias, M.; Jodynis-Liebert, J. 3,4,5,4′-trans-tetramethoxystilbene (DMU-212) modulates the activation of NF-κB, AP-1, and STAT3 transcription factors in rat liver carcinogenesis induced by initiation-promotion regimen. Mol. Cell. Biochem. 2014, 391, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Gosslau, A.; Pabbaraja, S.; Knapp, S.; Chen, K.Y. Trans- and cis-stilbene polyphenols induced rapid perinuclear mitochondrial clustering and p53-independent apoptosis in cancer cells but not normal cells. Eur. J. Pharmacol. 2008, 587, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Chun, Y.J.; Kim, S.; Kim, D.; Lee, S.K.; Guengerich, F.P. A new selective and potent inhibitor of human cytochrome P450 1B1 and its application to antimutagenesis. Cancer Res. 2001, 61, 8164–8170. [Google Scholar] [PubMed]
- Chun, Y.J.; Oh, Y.K.; Kim, B.J.; Kim, D.; Kim, S.S.; Choi, H.K.; Kim, M.Y. Potent inhibition of human cytochrome P450 1B1 by tetramethoxystilbene. Toxicol. Lett. 2009, 189, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Zhu, W.; Gonzalez, F.J. Potential role of CYP1B1 in the development and treatment of metabolic diseases. Pharmacol. Ther. 2017, 178, 18–30. [Google Scholar] [CrossRef] [PubMed]
- Peter Guengerich, F.; Chun, Y.J.; Kim, D.; Gillam, E.M.; Shimada, T. Cytochrome P450 1B1: A target for inhibition in anticarcinogenesis strategies. Mutat. Res. 2003, 523–524, 173–182. [Google Scholar] [CrossRef]
- Crofton, J.T.; Share, L.; Shade, R.E.; Lee-Kwon, W.J.; Manning, M.; Sawyer, W.H. The importance of vasopressin in the development and maintenance of doc-salt hypertension in the rat. Hypertension 1979, 1, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Malik, K.U.; Jennings, B.L.; Yaghini, F.A.; Sahan-Firat, S.; Song, C.Y.; Estes, A.M.; Fang, X.R. Contribution of cytochrome P450 1B1 to hypertension and associated pathophysiology: A novel target for antihypertensive agents. Prostaglandins Other Lip. Mediat. 2012, 98, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Sahan-Firat, S.; Jennings, B.L.; Yaghini, F.A.; Song, C.Y.; Estes, A.M.; Fang, X.R.; Farjana, N.; Khan, A.I.; Malik, K.U. 2,3′,4,5′-tetramethoxystilbene prevents deoxycorticosterone-salt-induced hypertension: Contribution of cytochrome P450 1B1. Am. J. physiol. 2010, 299, H1891–H1901. [Google Scholar] [CrossRef] [PubMed]
- Jennings, B.L.; Anderson, L.J.; Estes, A.M.; Fang, X.R.; Song, C.Y.; Campbell, W.B.; Malik, K.U. Involvement of cytochrome P-450 1B1 in renal dysfunction, injury, and inflammation associated with angiotensin ii-induced hypertension in rats. Am. J. physiol. 2012, 302, F408–E420. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.S.; Spatafora, C.; Tringali, C.; Ho, P.C. Determination of trans-2,4,3′,4′,5′-pentamethoxystilbene in rat plasma and its application to a pharmacokinetic study. J. Pharm. Biomed. Anal. 2012, 57, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.H.; Lin, C.L.; Tsai, J.H.; Ho, C.T.; Chen, W.J. 3,5,3′,4′,5′-pentamethoxystilbene (MR-5), a synthetically methoxylated analogue of resveratrol, inhibits growth and induces G1 cell cycle arrest of human breast carcinoma MCF-7 cells. J. Agric. Food Chem. 2010, 58, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, W.K.; Zheng, Z.; Che, C.T.; Yu, L.; Li, Z.J.; Wu, Y.C.; Cheng, K.W.; Yu, J.; Cho, C.H.; et al. 2,3′,4,4′,5′-pentamethoxy-trans-stilbene, a resveratrol derivative, is a potent inducer of apoptosis in colon cancer cells via targeting microtubules. Biochem. Pharmacol. 2009, 78, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Wu, W.K.; Li, Z.J.; Chan, K.M.; Wong, C.C.; Ye, C.G.; Yu, L.; Sung, J.J.; Cho, C.H.; Wang, M. 2,3′,4,4′,5′-pentamethoxy-trans-stilbene, a resveratrol derivative, inhibits colitis-associated colorectal carcinogenesis in mice. Br. J. Pharmacol. 2010, 160, 1352–1361. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.K.; Kim, Y.; Kim, M.Y.; Chun, Y.J.; Kim, S. Potent inhibition of recombinant human cytochrome P-450 1A1 by pentamethoxystilbene. J. Toxicol. Environ. Health Part A 2004, 67, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Shi, J.S.; Zhou, H.; Wilson, B.; Hong, J.S.; Gao, H.M. Resveratrol protects dopamine neurons against lipopolysaccharide-induced neurotoxicity through its anti-inflammatory actions. Mol. Pharmacol. 2010, 78, 466–477. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Yeo, S.C.; Elhennawy, M.G.; Xiang, X.; Lin, H.S. Determination of naturally occurring resveratrol analog trans-4,4′-dihydroxystilbene in rat plasma by liquid chromatography-tandem mass spectrometry: Application to a pharmacokinetic study. Anal. Bioanal. Chem. 2015, 407, 5793–5801. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Sumiyoshi, M.; Baba, K. Antitumor and antimetastatic activity of synthetic hydroxystilbenes through inhibition of lymphangiogenesis and M2 macrophage differentiation of tumor-associated macrophages. Anticancer Res. 2016, 36, 137–148. [Google Scholar] [PubMed]
- Saha, B.; Patro, B.S.; Koli, M.; Pai, G.; Ray, J.; Bandyopadhyay, S.K.; Chattopadhyay, S. Trans-4,4′-dihydroxystilbene (DHS) inhibits human neuroblastoma tumor growth and induces mitochondrial and lysosomal damages in neuroblastoma cell lines. Oncotarget 2017. [Google Scholar] [CrossRef]
- Kimura, Y.; Sumiyoshi, M.; Baba, K. Antitumor activities of synthetic and natural stilbenes through antiangiogenic action. Cancer Sci. 2008, 99, 2083–2096. [Google Scholar] [CrossRef] [PubMed]
- Maccario, C.; Savio, M.; Ferraro, D.; Bianchi, L.; Pizzala, R.; Pretali, L.; Forti, L.; Stivala, L.A. The resveratrol analog 4,4′-dihydroxy-trans-stilbene suppresses transformation in normal mouse fibroblasts and inhibits proliferation and invasion of human breast cancer cells. Carcinogenesis 2012, 33, 2172–2180. [Google Scholar] [CrossRef] [PubMed]
- Lappano, R.; Rosano, C.; Madeo, A.; Albanito, L.; Plastina, P.; Gabriele, B.; Forti, L.; Stivala, L.A.; Iacopetta, D.; Dolce, V.; et al. Structure-activity relationships of resveratrol and derivatives in breast cancer cells. Mol. Nutr. Food Res. 2009, 53, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Ruotolo, R.; Calani, L.; Fietta, E.; Brighenti, F.; Crozier, A.; Meda, C.; Maggi, A.; Ottonello, S.; Del Rio, D. Anti-estrogenic activity of a human resveratrol metabolite. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 1086–1092. [Google Scholar] [CrossRef] [PubMed]
- Balan, K.V.; Wang, Y.; Chen, S.W.; Chen, J.C.; Zheng, L.F.; Yang, L.; Liu, Z.L.; Pantazis, P.; Wyche, J.H.; Han, Z. Proteasome-independent down-regulation of estrogen receptor-alpha (ERα) in breast cancer cells treated with 4,4′-dihydroxy-trans-stilbene. Biochem. Pharmacol. 2006, 72, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Fan, G.J.; Liu, X.D.; Qian, Y.P.; Shang, Y.J.; Li, X.Z.; Dai, F.; Fang, J.G.; Jin, X.L.; Zhou, B. 4,4′-dihydroxy-trans-stilbene, a resveratrol analogue, exhibited enhanced antioxidant activity and cytotoxicity. Bioorg. Med. Chem. 2009, 17, 2360–2365. [Google Scholar] [CrossRef] [PubMed]
- Coppa, T.; Lazze, M.C.; Cazzalini, O.; Perucca, P.; Pizzala, R.; Bianchi, L.; Stivala, L.A.; Forti, L.; Maccario, C.; Vannini, V.; et al. Structure-activity relationship of resveratrol and its analogue, 4,4′-dihydroxy-trans-stilbene, toward the endothelin axis in human endothelial cells. J. Med. Food 2011, 14, 1173–1180. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Tsukahara, Y.; Kojima, T.; Murata, S.; Murakami, A.; Takada, K.; Takaoka, M.; Morimoto, S. Effects of phosphoramidon on endothelin-1 and big endothelin-1 production in human aortic endothelial cells. Biol. Pharm. Bull. 1995, 18, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.C.; Fang, J.G.; Chen, W.F.; Zhou, B.; Yang, L.; Liu, Z.L. Structure-activity relationship studies of resveratrol and its analogues by the reaction kinetics of low density lipoprotein peroxidation. Bioorg. Chem. 2006, 34, 142–157. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.G.; Lu, M.; Chen, Z.H.; Zhu, H.H.; Li, Y.; Yang, L.; Wu, L.M.; Liu, Z.L. Antioxidant effects of resveratrol and its analogues against the free-radical-induced peroxidation of linoleic acid in micelles. Chemistry 2002, 8, 4191–4198. [Google Scholar] [CrossRef]
- Setoguchi, Y.; Oritani, Y.; Ito, R.; Inagaki, H.; Maruki-Uchida, H.; Ichiyanagi, T.; Ito, T. Absorption and metabolism of piceatannol in rats. J. Agric. Food Chem. 2014, 62, 2541–2548. [Google Scholar] [CrossRef] [PubMed]
- Roupe, K.A.; Yanez, J.A.; Teng, X.W.; Davies, N.M. Pharmacokinetics of selected stilbenes: Rhapontigenin, piceatannol and pinosylvin in rats. J. Pharm. Pharmaco. 2006, 58, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.L.; Chan, S.W. A review of the pharmacological effects of piceatannol on cardiovascular diseases. Phytother. Res. 2014, 28, 1581–1588. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.J.; Lin, C.Y.; Hsieh, T.C.; Olson, S.C.; Wu, J.M. Control of eotaxin-1 expression and release by resveratrol and its metabolites in culture human pulmonary artery endothelial cells. Am. J. Cardiovasc. Dis. 2011, 1, 16–30. [Google Scholar] [PubMed]
- Choi, K.H.; Kim, J.E.; Song, N.R.; Son, J.E.; Hwang, M.K.; Byun, S.; Kim, J.H.; Lee, K.W.; Lee, H.J. Phosphoinositide 3-kinase is a novel target of piceatannol for inhibiting PDGF-BB-induced proliferation and migration in human aortic smooth muscle cells. Cardiovasc. Res. 2010, 85, 836–844. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, Y.; Kawakami, S.; Yanae, K.; Sano, S.; Uchida, H.; Inagaki, H.; Ito, T. Effect of long-term piceatannol treatment on eNOS levels in cultured endothelial cells. Biochem. Biophys. Res. Commun. 2013, 430, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Ordemann, J.; Muller, J.M.; Dubiel, W. The COP9 signalosome, cullin 3 and Keap1 supercomplex regulates CHOP stability and adipogenesis. Biol. Open 2012, 1, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Khurana, R.; Simons, M.; Martin, J.F.; Zachary, I.C. Role of angiogenesis in cardiovascular disease: A critical appraisal. Circulation 2005, 112, 1813–1824. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.Y.; Wang, F.; Li, Y.L.; Liu, K.; Xu, H. Antioxidant activities of stilbenoids from rheum emodi wall. Evid. Based Complement. Altern. Med. 2012, 2012, 603678. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Park, S.A.; Almazari, I.; Kim, E.H.; Na, H.K.; Surh, Y.J. Piceatannol induces heme oxygenase-1 expression in human mammary epithelial cells through activation of are-driven NRF2 signaling. Arch. Biochem. Biophys. 2010, 501, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, K.W.; Kim, M.S.; Lee, H.J. Piceatannol attenuates hydrogen-peroxide- and peroxynitrite-induced apoptosis of PC12 cells by blocking down-regulation of Bcl-XL and activation of JNK. J. Nutr. Biochem. 2008, 19, 459–466. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Tomas-Barberan, F.A.; Espin, J.C. The grape and wine polyphenol piceatannol is a potent inducer of apoptosis in human sk-mel-28 melanoma cells. Eur. J. Nutr. 2004, 43, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Tomas-Barberan, F.A.; Espin, J.C. Grape polyphenol resveratrol and the related molecule 4-hydroxystilbene induce growth inhibition, apoptosis, s-phase arrest, and upregulation of cyclins A, E, and B1 in human Sk-Mel-28 melanoma cells. J. Agric. Food Chem. 2003, 51, 4576–4584. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, Y.; Lee, Y.; Chung, J.H. Ceramide accelerates ultraviolet-induced MMP-1 expression through jak1/stat-1 pathway in cultured human dermal fibroblasts. J. Lip. Res. 2008, 49, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.E.; Karras, J.G.; Murphy, T.F.; Barton, A.; Huang, H.F. Signal transducer and activator of transcription 3 (STAT3) activation in prostate cancer: Direct STAT3 inhibition induces apoptosis in prostate cancer lines. Mol. Cancer Ther. 2004, 3, 11–20. [Google Scholar] [PubMed]
- Lee, Y.M.; Lim, D.Y.; Cho, H.J.; Seon, M.R.; Kim, J.K.; Lee, B.Y.; Park, J.H. Piceatannol, a natural stilbene from grapes, induces G1 cell cycle arrest in androgen-insensitive DU145 human prostate cancer cells via the inhibition of CDK activity. Cancer Lett. 2009, 285, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Wolter, F.; Clausnitzer, A.; Akoglu, B.; Stein, J. Piceatannol, a natural analog of resveratrol, inhibits progression through the s phase of the cell cycle in colorectal cancer cell lines. J. Nutr. 2002, 132, 298–302. [Google Scholar] [PubMed]
- Kuo, P.L.; Hsu, Y.L. The grape and wine constituent piceatannol inhibits proliferation of human bladder cancer cells via blocking cell cycle progression and inducing FAS/membrane bound FAS ligand-mediated apoptotic pathway. Mol. Nutr. Food Res. 2008, 52, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Lancon, A.; Hanet, N.; Jannin, B.; Delmas, D.; Heydel, J.M.; Lizard, G.; Chagnon, M.C.; Artur, Y.; Latruffe, N. Resveratrol in human hepatoma HepG2 cells: Metabolism and inducibility of detoxifying enzymes. Drug Metab. Dispos. Boil. Fate Chem. 2007, 35, 699–703. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Park, C.; Lee, J.O.; Kim, G.Y.; Lee, W.H.; Choi, Y.H.; Ryu, C.H. Induction of apoptosis by piceatannol in human leukemic U937 cells through down-regulation of BCL-2 and activation of caspases. Oncol. Rep. 2008, 19, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.H.; Chang, L.S. Piceatannol induces Fas and Fasl up-regulation in human leukemia U937 cells via Ca2+/p38α MAPK-mediated activation of c-JUN and ATF-2 pathways. Int. J. Biochem. Cell Biol. 2010, 42, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Fullbeck, M.; Huang, X.; Dumdey, R.; Frommel, C.; Dubiel, W.; Preissner, R. Novel curcumin- and emodin-related compounds identified by in silico 2D/3D conformer screening induce apoptosis in tumor cells. BMC Cancer 2005, 5, 97. [Google Scholar] [CrossRef] [PubMed]
- Duarte, N.; Kayser, O.; Abreu, P.; Ferreira, M.J. Antileishmanial activity of piceatannol isolated from euphorbia lagascae seeds. Phytother. Res. 2008, 22, 455–457. [Google Scholar] [CrossRef] [PubMed]
- Kedzierski, L.; Curtis, J.M.; Kaminska, M.; Jodynis-Liebert, J.; Murias, M. In vitro antileishmanial activity of resveratrol and its hydroxylated analogues against leishmania major promastigotes and amastigotes. Parasitol. Res. 2007, 102, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.C.; Sharma, M.; Sharma, A. Inhibitory effect of piceatannol, a protein tyrosine kinase inhibitor, on asexual maturation of plasmodium falciparum. Indian J. Exp. Boil. 1999, 37, 418–420. [Google Scholar]
- Sharma, A.; Mishra, N.C. Inhibition of a protein tyrosine kinase activity in plasmodium falciparum by chloroquine. Indian J. Biochem. Biophys. 1999, 36, 299–304. [Google Scholar] [PubMed]
- Docherty, J.J.; McEwen, H.A.; Sweet, T.J.; Bailey, E.; Booth, T.D. Resveratrol inhibition of propionibacterium acnes. J. Antimicrob. Chemother. 2007, 59, 1182–1184. [Google Scholar] [CrossRef] [PubMed]
- Angeles, D.C.; Ho, P.; Dymock, B.W.; Lim, K.L.; Zhou, Z.D.; Tan, E.K. Antioxidants inhibit neuronal toxicity in parkinson’s disease-linked LRRK2. Ann. Clin. Transl. Neurol. 2016, 3, 288–294. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, K.W.; Lee, H.J. Protective effects of piceatannol against beta-amyloid-induced neuronal cell death. Ann. N. Y. Acad. Sci. 2007, 1095, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Pavanetto, M.; Zarpellon, A.; Borgo, C.; Donella-Deana, A.; Deana, R. Regulation of serotonin transport in human platelets by tyrosine kinase syk. Cell. Physiol. Biochem. 2011, 27, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Bastianetto, S.; Dumont, Y.; Han, Y.; Quirion, R. Comparative neuroprotective properties of stilbene and catechin analogs: Action via a plasma membrane receptor site? CNS Neurosci. Ther. 2009, 15, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Mikula-Pietrasik, J.; Sosinska, P.; Wierzchowski, M.; Piwocka, K.; Ksiazek, K. Synthetic resveratrol analogue, 3,3′,4,4′,5,5′-hexahydroxy-trans-stilbene, accelerates senescence in peritoneal mesothelium and promotes senescence-dependent growth of gastrointestinal cancers. Int. J. Mol. Sci. 2013, 14, 22483–22498. [Google Scholar] [CrossRef] [PubMed]
- Murias, M.; Handler, N.; Erker, T.; Pleban, K.; Ecker, G.; Saiko, P.; Szekeres, T.; Jager, W. Resveratrol analogues as selective cyclooxygenase-2 inhibitors: Synthesis and structure-activity relationship. Bioorg. Med. Chem. 2004, 12, 5571–5578. [Google Scholar] [CrossRef] [PubMed]
- Murias, M.; Luczak, M.W.; Niepsuj, A.; Krajka-Kuzniak, V.; Zielinska-Przyjemska, M.; Jagodzinski, P.P.; Jager, W.; Szekeres, T.; Jodynis-Liebert, J. Cytotoxic activity of 3,3′,4,4′,5,5′-hexahydroxystilbene against breast cancer cells is mediated by induction of p53 and downregulation of mitochondrial superoxide dismutase. Toxicol. In Vitro 2008, 22, 1361–1370. [Google Scholar] [CrossRef] [PubMed]
- Saiko, P.; Pemberger, M.; Horvath, Z.; Savinc, I.; Grusch, M.; Handler, N.; Erker, T.; Jaeger, W.; Fritzer-Szekeres, M.; Szekeres, T. Novel resveratrol analogs induce apoptosis and cause cell cycle arrest in HT29 human colon cancer cells: Inhibition of ribonucleotide reductase activity. Oncol. Rep. 2008, 19, 1621–1626. [Google Scholar] [PubMed]
- Kucinska, M.; Piotrowska, H.; Luczak, M.W.; Mikula-Pietrasik, J.; Ksiazek, K.; Wozniak, M.; Wierzchowski, M.; Dudka, J.; Jager, W.; Murias, M. Effects of hydroxylated resveratrol analogs on oxidative stress and cancer cells death in human acute T cell leukemia cell line: Prooxidative potential of hydroxylated resveratrol analogs. Chem. Boil. Interact. 2014, 209, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Paulitschke, V.; Schicher, N.; Szekeres, T.; Jager, W.; Elbling, L.; Riemer, A.B.; Scheiner, O.; Trimurtulu, G.; Venkateswarlu, S.; Mikula, M.; et al. 3,3′,4,4′,5,5′-hexahydroxystilbene impairs melanoma progression in a metastatic mouse model. J. Invest. Dermatol. 2010, 130, 1668–1679. [Google Scholar] [CrossRef] [PubMed]
- Horvath, Z.; Murias, M.; Saiko, P.; Erker, T.; Handler, N.; Madlener, S.; Jaeger, W.; Grusch, M.; Fritzer-Szekeres, M.; Krupitza, G.; et al. Cytotoxic and biochemical effects of 3,3′,4,4′,5,5′-hexahydroxystilbene, a novel resveratrol analog in HL-60 human promyelocytic leukemia cells. Exp. Hematol. 2006, 34, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Pai, R.S. Recent advances of resveratrol in nanostructured based delivery systems and in the management of HIV/aids. J. Control. Release 2014, 194, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Clouser, C.L.; Chauhan, J.; Bess, M.A.; van Oploo, J.L.; Zhou, D.; Dimick-Gray, S.; Mansky, L.M.; Patterson, S.E. Anti-HIV-1 activity of resveratrol derivatives and synergistic inhibition of HIV-1 by the combination of resveratrol and decitabine. Bioorg. Med. Chem. Lett. 2012, 22, 6642–6646. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.S.; Quashie, P.K.; Mesplede, T.; Xu, H.; Quan, Y.; Jaeger, W.; Szekeres, T.; Wainberg, M.A. A resveratrol analog termed 3,3′,4,4′,5,5′-hexahydroxy-trans-stilbene is a potent HIV-1 inhibitor. J. Med. Virol. 2015, 87, 2054–2060. [Google Scholar] [CrossRef] [PubMed]
- Moran, B.W.; Anderson, F.P.; Devery, A.; Cloonan, S.; Butler, W.E.; Varughese, S.; Draper, S.M.; Kenny, P.T. Synthesis, structural characterisation and biological evaluation of fluorinated analogues of resveratrol. Bioorg. Med. Chem. 2009, 17, 4510–4522. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.J.; Min, H.Y.; Joo Park, H.; Chung, H.J.; Kim, S.; Nam Han, Y.; Lee, S.K. G2/m cell cycle arrest and induction of apoptosis by a stilbenoid, 3,4,5-trimethoxy-4′-bromo-cis-stilbene, in human lung cancer cells. Life Sci. 2004, 75, 2829–2839. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, G.T.; Gertz, M.; Steegborn, C. Crystal structures of Sirt3 complexes with 4′-bromo-resveratrol reveal binding sites and inhibition mechanism. Chem. Boil. 2013, 20, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Bourgault, S.; Choi, S.; Buxbaum, J.N.; Kelly, J.W.; Price, J.L.; Reixach, N. Mechanisms of transthyretin cardiomyocyte toxicity inhibition by resveratrol analogs. Biochem. Biophys. Res. Commun. 2011, 410, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Nagaradja, E.; Bentabed-Ababsa, G.; Scalabrini, M.; Chevallier, F.; Philippot, S.; Fontanay, S.; Duval, R.E.; Halauko, Y.S.; Ivashkevich, O.A.; Matulis, V.E.; et al. Deprotometalation-iodolysis and computed CH acidity of 1,2,3- and 1,2,4-triazoles. Application to the synthesis of resveratrol analogues. Bioorg. Med. Chem. 2015, 23, 6355–6363. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methoxylated Resveratrol Derivatives | Mechanism | Therapeutic Benefits | Comparison with RSV | Effect on Biomarker | References |
|---|---|---|---|---|---|
| Pterostilbene | Suppressing various signal transduction pathways. Inhibiting ethanol-induced oxidative DNA damage. | Antioxidant. Chemotherapeutic activity in pancreatic, melanoma, leukemia, breast, lung and gastric cancer. apoptosis, Anti-invasive, antimetastatic, anti-inflammatory. Manage hypertension and vessel diameter. Antidiabetic. | Increased lipophilicity over resveratrol. | COX-2 ↓, iNOS ↓, NF-ĸB ↓, AP-1 ↓, MMP-9 ↓, Akt ↓, p38 MAPK ↓, TNF-α ↓, IL-1b ↓, IL-6 ↓, COX-1 ↓, COX-2 ↓, LDL ↓, HDL ↑, | [30,33,36,40,42,44,49,85] |
| Trimethoxystilbene | Down-regulating phosphatidylinositol 3-kinase (PI3K)/AKT signaling. Blockade of neovascularization by antiangiogenic compounds. Immature vessel disruption by vascular-targeting agents. Bundling of microtubules and speckle-like structures of DCLK1—microtubule complexes. | Chemotherapeutic role against breast, lung and liver cancer. Vascular-targeting agent. Anti-HCV | 30 to 100 time more potent than the resveratrol in inhibiting endothelial cell proliferation and morphogenesis. | NF-ĸB ↓, AP-1 ↓, MMP-2 ↓, MMP-9 ↓, VEGFR2 mRNA expression ↓, β-catenin ↓. | [54,55,56,57,61] |
| Tetramethoxy stilbene | Perinuclear mitochondrial clustering by membrane permeability transition, release of cytochrome c into the cytosol and DNA fragmentation. Anti-angiogenic activity by Inhibiting the phosphorylation of multiple downstream signaling components of VEGFR2. | Inhibiting the growth of various cancers, including colon, prostate, ovarian and liver. Inhibits Cytochromes P450. Manage hypertension and cardiac fibrosis. | High potency and bioavailability than resveratrol. | Bax ↑, Bcl-2 ↓. Akt ↓, FAK ↓, c-Src ↓, mTOR ↓, p70S6K ↓ and Erk1/2 ↓. | [65,68,69,70,73,75,76,77] |
| Pentamethoxystilbene | Induces G1 cell-cycle arrest and G1 cell-cycle regulatory proteins. | Inhibiting the growth of breast and colon cancer. Inhibits Cytochromes P450. | Potent inhibition of cell growth than resveratrol | cyclin D1 ↓, D3 ↓ and E ↓, CDK2 ↓, 4 ↓ and 6 ↓, ERK1 ↓, p38 MAPK ↓. | [81,83,84] |
| Hydroxylated Resveratrol Derivatives | Mechanism | Therapeutic Benefits | Comparison with Resveratrol | Effect on Biomarkers | Reference |
|---|---|---|---|---|---|
| Dihydroxystilbenes | Inhibits cancer progression by metastasis and tumor growth via G1-phase arrest. Reduced total endothelin-1 secretion and endothelin-1 messenger RNA (mRNA) levels in human endothelial cells. Active in protecting against hemin-induced lipid peroxidation and ROS production. | Chemotherapeutic against lung and breast cancer. Manage vascular abnormalities. Antioxidant. | More active than resveratrol. | p21↑, p53↑, VEGF↓, LDL↓ | [87,90,92,93,97,98] |
| Tetrahydroxystilbene | Inhibits oxidation of LDL-c in plasma, platelet aggregation, and inflammation. Antiproliferative, cytotoxic, hormesis and proapototic aginist cancer cells. Suppress the mutation of leucine-rich repeat kinase-2, inhibits the Serotonin uptake. | Management of atherosclerosis, hypertension, myocardial ischemia. Chemotherapeutic activity against liver, leukemia and cervix cancer. Antioxidant. Antimicrobial. Effective in Parkinson and Alzheimer. | Stronger antioxidant and tumor suppressing activity than resveratrol. | Bcl-2↓, COX 2↓, LDL↓, AMPK↑, NF-κB↓, Cyclin D1↓, PTK↓. | [101,111,112,121,123,126,127,129] |
| Hexahydroxystilbene | Cause a dysbalance of intra-cellular deoxyribonucleoside triphosphates. Inhibition of viral attachment and reverse transcription to host cells before replication. Inhibits prostaglandin-endoperoxide synthase. | Inhibit growth in numerous malignancies, including breast and colon cancers ,leukemia, melanoma, and glioma cells. Anti HIV-1. selective COX-2 inhibitor, Antioxidant. | 6600-fold higher antiradical activity than resveratrol, most effective free radical scavenger of all resveratrol analogues, higher anti-HIV-1 activity than resveratrol. | NF-κB, p53↑, COX-1↓ and COX-2↓, SOD↑, SA-β-gal↓, SIRT1 expression↑. | [130,131,133,135,137] |
| Halogenated Resveratrol Derivatives | Mechanism | Therapeutic Effects | Comparison with Resveratrol | Reference |
|---|---|---|---|---|
| (E)-2,6-dibromo-4-(3,5-dibromostyryl)phenol | stabilize the native tetramer of amyloid transthyretin and modify the quaternary structure of monomeric transthyretin in solution | Cardiprotective effect | Higher bioavailability than resveratrol | [143] |
| (E)-3,5-di-fluoro-4′-acetoxystilbene | Inhibiting upregulation of cellular transporter proteins belonging to the ABC superfamily | antiproliferative | greater anticancer activity than resveratrol | [140] |
| 3,4,5-trimethoxy-4′-brom-cis-stilbene | Suppressing the growth of cancer cell through G2/M phase cell cycle arrest | inhibitor of the growth of lung cancer cells | more effective than in suppressing tumor growth than resveratrol | [141] |
| 4′-Bromo-Resveratrol | potently inhibited Sirt1 and Sirt3 by overlapping through extending its bromo-phenyl group at the active site | Therapeutic effects in aging, transcription, apoptosis, inflammation related diseases | inhibited Sirt3 with much higher potency than resveratrol | [142] |
| 2-bromo-resveratrol | unknown | Antimicrobial, Antiproliferative | 3 fold lower MIC values against C. albicans than resveratrol | [144] |
| 2-chloro-resveratrol | unknown | Antimicrobial, Antiproliferative | 30 fold lower MIC values against C. albicans than resveratrol | [144] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nawaz, W.; Zhou, Z.; Deng, S.; Ma, X.; Ma, X.; Li, C.; Shu, X. Therapeutic Versatility of Resveratrol Derivatives. Nutrients 2017, 9, 1188. https://doi.org/10.3390/nu9111188
Nawaz W, Zhou Z, Deng S, Ma X, Ma X, Li C, Shu X. Therapeutic Versatility of Resveratrol Derivatives. Nutrients. 2017; 9(11):1188. https://doi.org/10.3390/nu9111188
Chicago/Turabian StyleNawaz, Waqas, Zhongqin Zhou, Sa Deng, Xiaodong Ma, Xiaochi Ma, Chuangang Li, and Xiaohong Shu. 2017. "Therapeutic Versatility of Resveratrol Derivatives" Nutrients 9, no. 11: 1188. https://doi.org/10.3390/nu9111188
APA StyleNawaz, W., Zhou, Z., Deng, S., Ma, X., Ma, X., Li, C., & Shu, X. (2017). Therapeutic Versatility of Resveratrol Derivatives. Nutrients, 9(11), 1188. https://doi.org/10.3390/nu9111188