Restoration of Tear Secretion in a Murine Dry Eye Model by Oral Administration of Palmitoleic Acid



Abstract
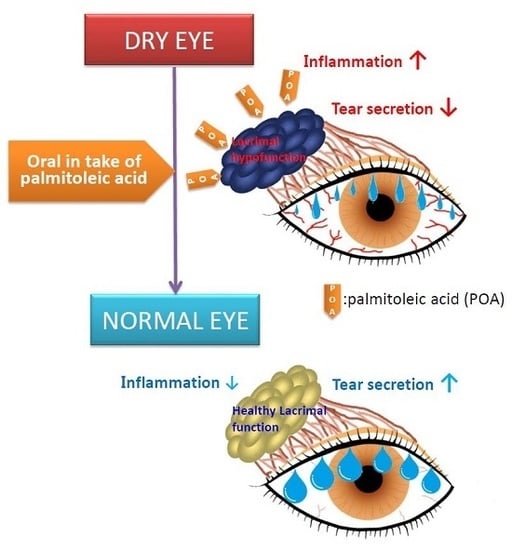
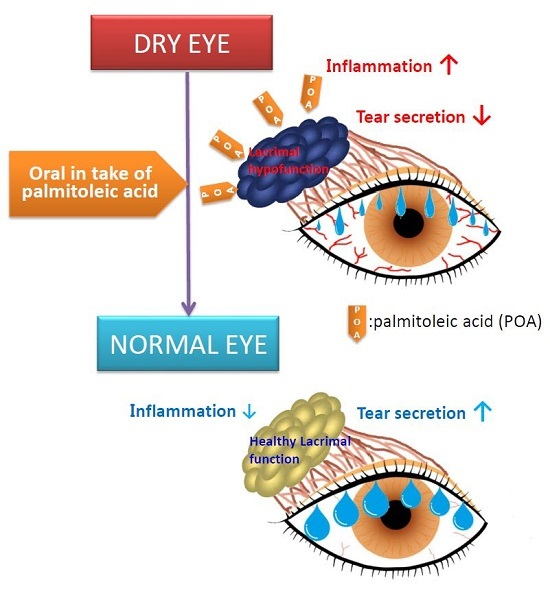
:
1. Introduction
2. Materials and Methods
2.1. Oil Products and Fatty Acids
2.2. Chemicals
2.3. Animals
2.4. Murine Dry Eye Models
2.5. Mouse Stress-Induced Dry Eye Model
2.6. Rat Blink-Suppressed Dry Eye Model
2.7. Measurement of Aqueous Tear Secretion
2.8. Measurement of Fatty Acids in Oil Products and Mouse Serum
2.9. Lacrimal Gland Cytokine Measurement
2.10. Statistical Analysis
3. Results
3.1. Fatty Acid Compositions of Sea Buckthorn Oil Products and Olive Oil
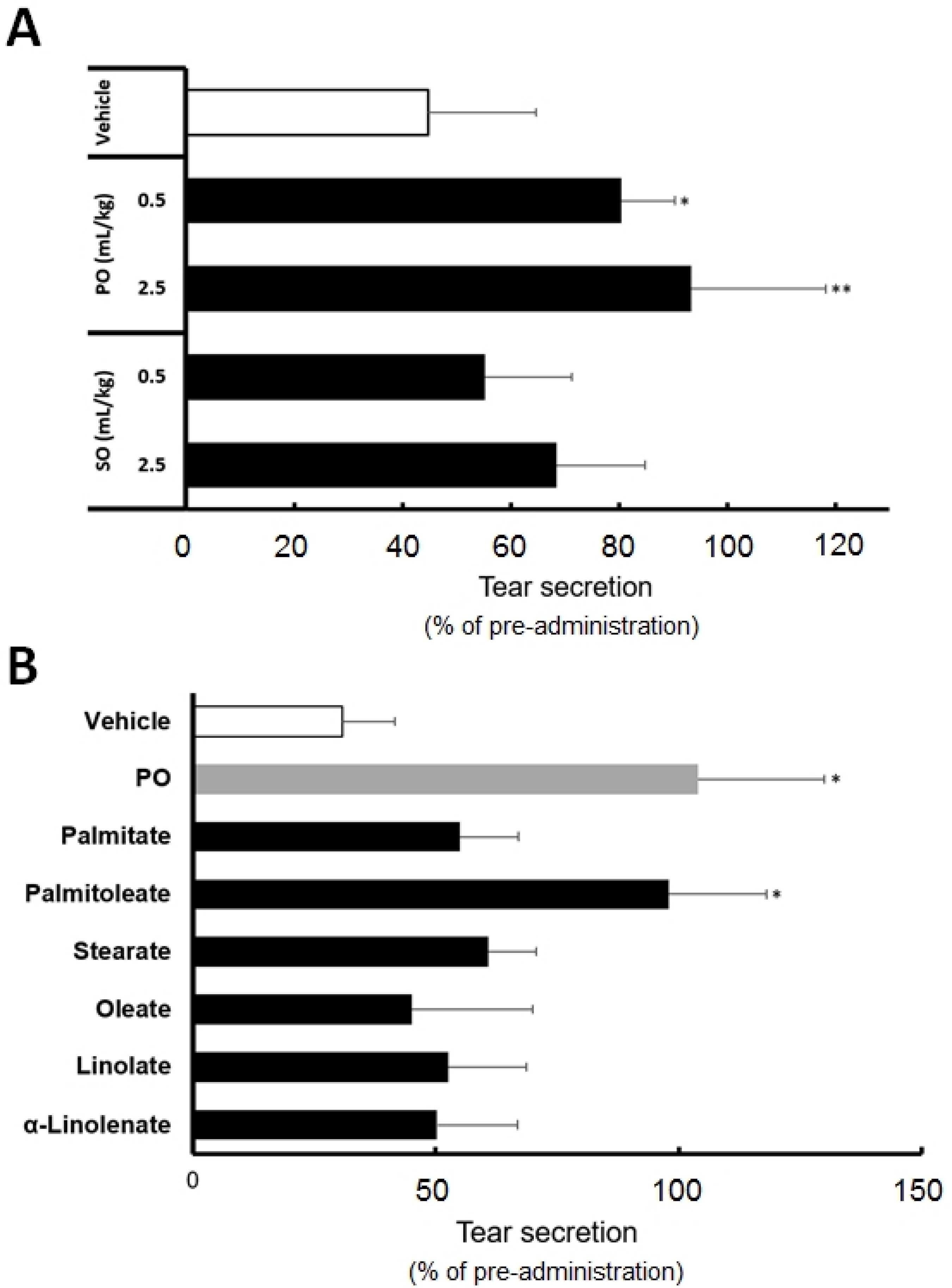
3.2. The Effects of Sea Buckthorn Oil Products on Tear Secretion
3.3. Palmitoleate Is an Active Component of Sea Buckthorn Pulp Oil
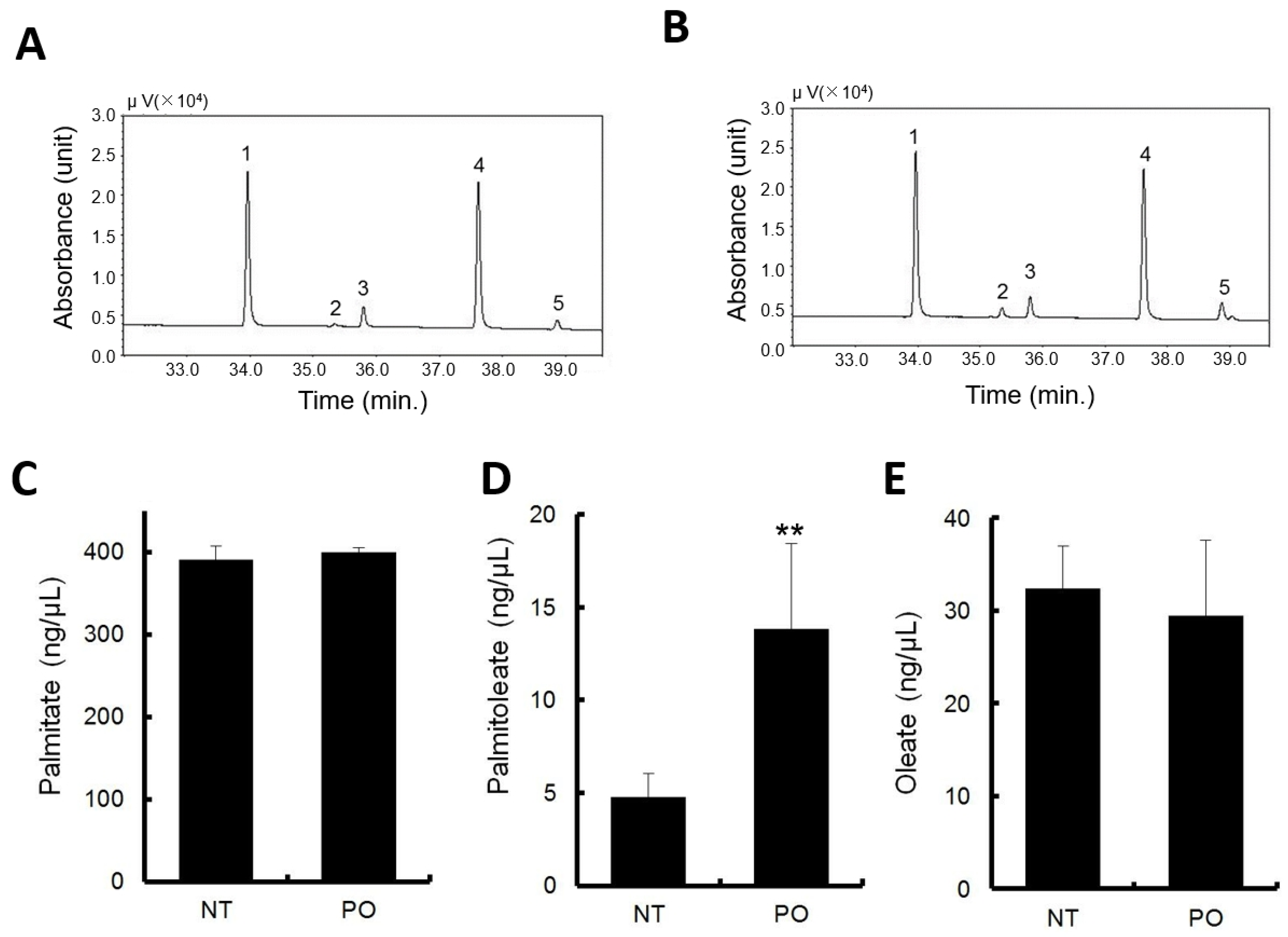
3.4. Orally Administrated Sea Buckthorn Pulp Oil Changed Serum Fatty Acid Composition
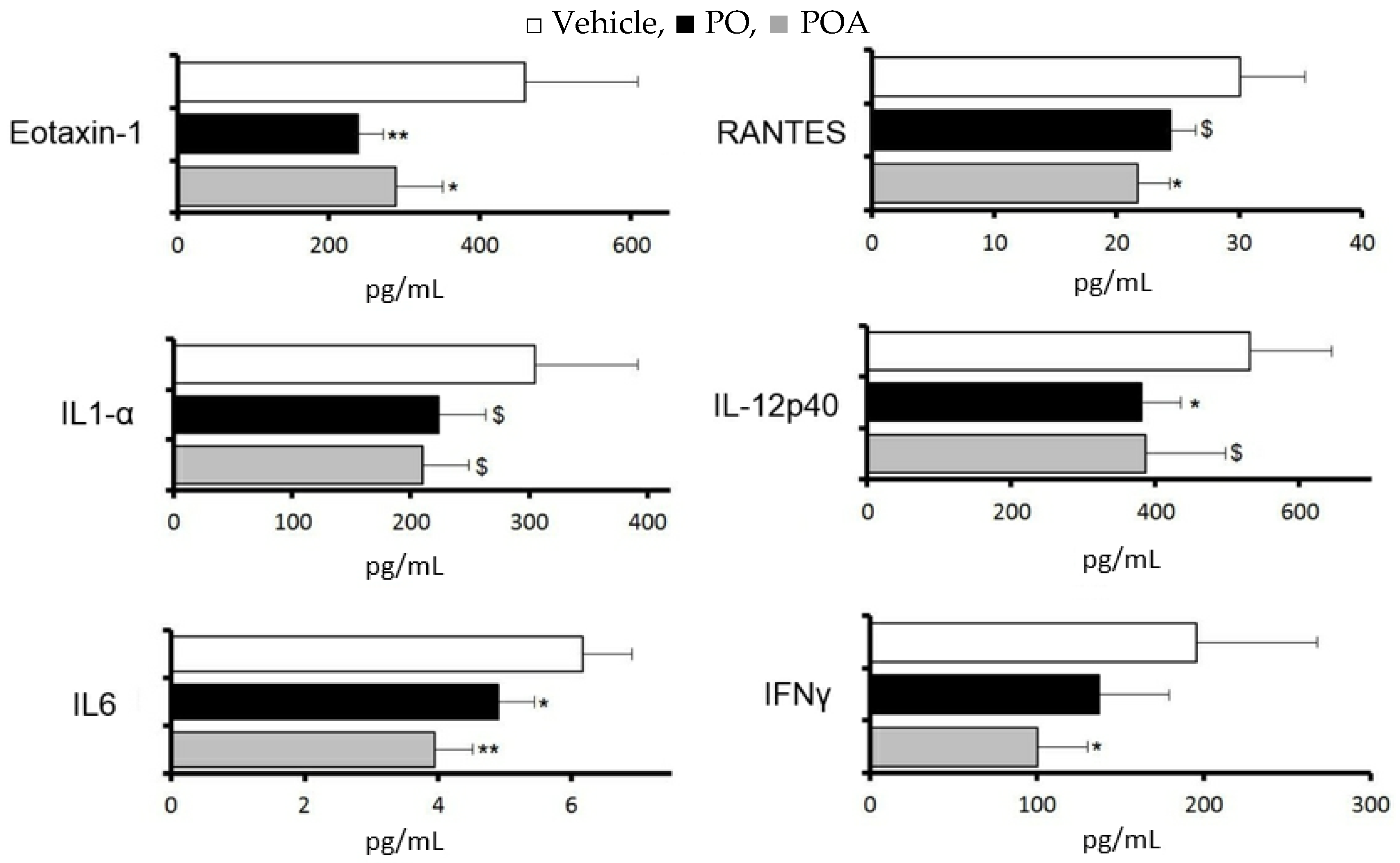
3.5. Changes of Cytokine and Chemokine Expression in the LG after Repeated Treatment with Sea Buckthorn Pulp Oil or Palmitoleate
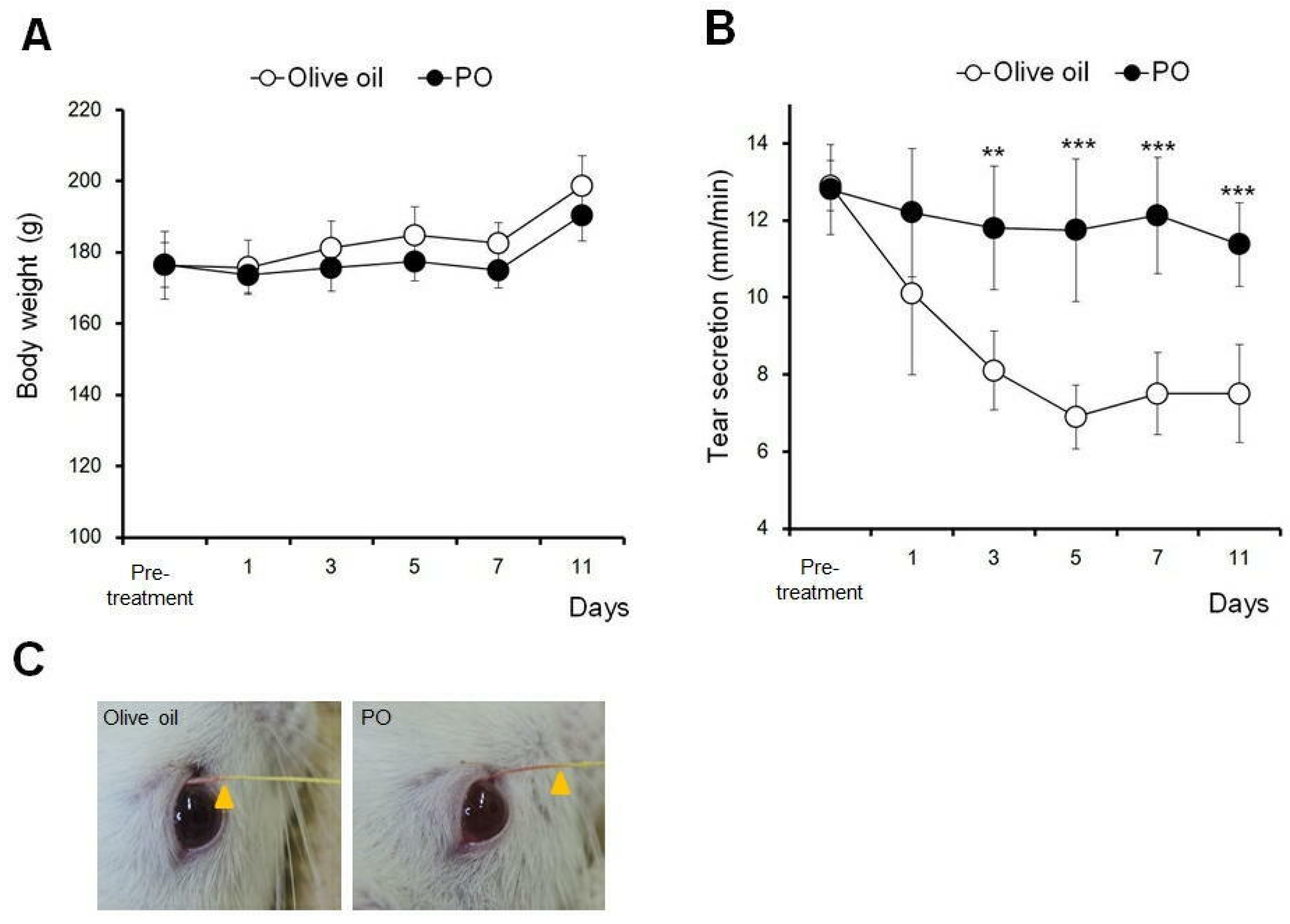
3.6. Sea Buckthorn Pulp Oil, Not Olive Oil, Restores Tear Secretion in a Rat Blink-Suppressed Dry Eye Model
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| PO | Sea buckthorn pulp oil |
| SO | Sea buckthorn seed oil |
| POA | Palmitoleate |
References
- Swenson, U.; Bartish, I.V. Taxonomic synopsis of Hippophae (Elaeagnaceae). Nord. J. Bot. 2002, 22, 369–374. [Google Scholar] [CrossRef]
- Suryakumar, G.; Gupta, A. Medicinal and therapeutic potential of Sea buckthorn (Hippophae rhamnoides L.). J. Ethnopharmacol. 2011, 138, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Larmo, P. The Health Effects of Sea Buckthorn Berries and Oil; University of Turku: Turku, Finland, 2011. [Google Scholar]
- Bal, L.M.; Meda, V.; Naik, S.N.; Satya, S. Sea buckthorn berries: A potential source of valuable nutrients for nutraceuticals and cosmoceuticals. Food Res. Int. 2011, 44, 1718–1727. [Google Scholar] [CrossRef]
- Yang, B.; Kallio, H. Composition and physiological effects of sea buckthorn (Hippophaë) lipids. Trends Food Sci. Technol. 2002, 13, 160–167. [Google Scholar] [CrossRef]
- Ito, H.; Asmussen, S.; Traber, D.L.; Cox, R.A.; Hawkins, H.K.; Connelly, R.; Traber, L.D.; Walker, T.W.; Malgerud, E.; Sakurai, H.; Enkhbaatar, P. Healing efficacy of sea buckthorn (Hippophae rhamnoides L.) seed oil in an ovine burn wound model. Burns 2014, 40, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.K.; Yogendra Kumar, M.S.; Gupta, A. Antioxidant, cytoprotective and antibacterial effects of Sea buckthorn (Hippophae rhamnoides L.) leaves. Food Chem. Toxicol. 2010, 48, 3443–3448. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, K. Tear dynamics and dry eye. Prog. Retin. Eye Res. 1998, 17, 565–596. [Google Scholar] [CrossRef]
- Kawashima, M.; Uchino, M.; Kawazoe, T.; Kamiyashiki, M.; Sano, K.; Tsubota, K. A field test of Web-based screening for dry eye disease to enhance awareness of eye problems among general Internet users: A latent strategy to promote health. J. Med. Internet Res. 2013, 15, e209. [Google Scholar] [CrossRef] [PubMed]
- Larmo, P.S.; Jä, R.L.; Setä, N.L.; Yang, B.; Viitanen, M.H.; Engblom, J.R.K.; Tahvonen, R.L.; Kallio, H.P. Oral Sea Buckthorn Oil Attenuates Tear Film Osmolarity and Symptoms in Individuals with Dry Eye 1–4. J. Nutr. 2010, 140, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Kallio, H.P. Fatty Acid Composition of Lipids in Sea Buckthorn (Hippophaë rhamnoides L.) Berries of Different Origins. J. Agric. Food Chem. 2001, 49, 1939–1947. [Google Scholar] [CrossRef] [PubMed]
- Beveridge, T.; Li, T.S.C.; Oomah, B.D.; Smith, A. Sea buckthorn products: Manufacture and composition. J. Agric. Food Chem. 1999, 47, 3840–3848. [Google Scholar] [CrossRef]
- Rustan, A.C.; Drevon, C.A. Fatty Acids: Structures and Properties. eLS 2005. [Google Scholar] [CrossRef]
- Nakamura, S.; Tanaka, J.; Imada, T.; Shimoda, H.; Tsubota, K. Delphinidin 3,5-O-diglucoside, a constituent of the maqui berry (Aristotelia chilensis) anthocyanin, restores tear secretion in a rat dry eye model. J. Funct. Foods 2014, 10, 346–354. [Google Scholar] [CrossRef]
- Nakamura, S.; Kinoshita, S.; Yokoi, N.; Ogawa, Y.; Shibuya, M.; Nakashima, H.; Hisamura, R.; Imada, T.; Imagawa, T.; Uehara, M.; et al. Lacrimal hypofunction as a new mechanism of dry eye in visual display terminal users. PLoS ONE 2010, 5, e11119. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B. The Mediterranean Diet and Mortality—Olive Oil and Beyond. N. Engl. J. Med. 2003, 348, 2595–2596. [Google Scholar] [CrossRef] [PubMed]
- Paillard, F.; Catheline, D.; Le Duff, F.; Bouriel, M.; Deugnier, Y.; Pouchard, M.; Daubert, J.C.; Legrand, P. Plasma palmitoleic acid, a product of stearoyl-coA desaturase activity, is an independent marker of triglyceridemia and abdominal adiposity. Nutr. Metab. Cardiovasc. Dis. 2008, 18, 436–440. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Gerhold, K.; Mayers, J.R.; Wiest, M.M.; Watkins, S.M.; Hotamisligil, G.S. Identification of a Lipokine, a Lipid Hormone Linking Adipose Tissue to Systemic Metabolism. Cell 2008, 134, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Çimen, I.; Kocatürk, B.; Koyuncu, S.; Tufanlı, Ö.; Onat, U.I.; Yıldırım, A.D.; Apaydın, O.; Demirsoy, Ş.; Aykut, Z.G.; Nguyen, U.T.; et al. Prevention of atherosclerosis by bioactive palmitoleate through suppression of organelle stress and inflammasome activation. Sci. Transl. Med. 2016, 8, 358ra126. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Li, H.; Xu, H.; Halim, V.; Zhang, W.; Wang, H.; Ong, K.T.; Woo, S.L.; Walzem, R.L.; Mashek, D.G.; et al. Palmitoleate induces hepatic steatosis but suppresses liver inflammatory response in mice. PLoS ONE 2012, 7, e39286. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.O.; Teixeira, A.A.S.; Lima, E.A.; Batatinha, H.A.P.; Gomes, L.M.; Carvalho-Silva, M.; Mota, I.T.; Streck, E.L.; Hirabara, S.M.; Rosa Neto, J.C. Palmitoleic acid (n-7) attenuates the immunometabolic disturbances caused by a high-fat diet independently of PPARα. Mediat. Inflamm. 2014, 2014, 582197. [Google Scholar] [CrossRef] [PubMed]
- Tsubota, K. Oxidative stress and inflammation: Hypothesis for the mechanism of aging. Nihon. Ganka Gakkai Zasshi 2007, 111, 193–205, discussion 206. [Google Scholar] [PubMed]
- Wakamatsu, T.H.; Dogru, M.; Matsumoto, Y.; Kojima, T.; Kaido, M.; Ibrahim, O.M.A.; Sato, E.A.; Igarashi, A.; Ichihashi, Y.; Satake, Y.; et al. Evaluation of lipid oxidative stress status in Sjögren syndrome patients. Invest. Ophthalmol. Vis. Sci. 2013, 54, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Calonge, M.; Enríquez-de-Salamanca, A.; Diebold, Y.; González-García, M.J.; Reinoso, R.; Herreras, J.M.; Corell, A. Dry eye disease as an inflammatory disorder. Ocul. Immunol. Inflamm. 2010, 18, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Jabs, D.A.; Lee, B.; Whittum-Hudson, J.A.; Prendergast, R.A. Th1 versus Th2 immune responses in autoimmune lacrimal gland disease in MRL/Mp mice. Investig. Ophthalmol. Vis. Sci. 2000, 41, 826–831. [Google Scholar]
- Teran, L.M.; Mochizuki, M.; Bartels, J.; Valencia, E.L.; Nakajima, T.; Hirai, K.; Schröder, J.M. Th1- and Th2-type cytokines regulate the expression and production of eotaxin and RANTES by human lung fibroblasts. Am. J. Respir. Cell Mol. Biol. 1999, 20, 777–786. [Google Scholar] [CrossRef] [PubMed]
- Selenica, M.-L.B.; Alvarez, J.A.; Nash, K.R.; Lee, D.C.; Cao, C.; Lin, X.; Reid, P.; Mouton, P.R.; Morgan, D.; Gordon, M.N. Diverse activation of microglia by chemokine (C-C motif) ligand 2 overexpression in brain. J. Neuroinflamm. 2013, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Gauldie, J.; Richards, C.; Harnish, D.; Lansdorp, P.; Baumann, H. Interferon beta 2/B-cell stimulatory factor type 2 shares identity with monocyte-derived hepatocyte-stimulating factor and regulates the major acute phase protein response in liver cells. Proc. Natl. Acad. Sci. USA 1987, 84, 7251–7255. [Google Scholar] [CrossRef] [PubMed]
- Lam, H.; Bleiden, L.; de Paiva, C.S.; Farley, W.; Stern, M.E.; Pflugfelder, S.C. Tear Cytokine Profiles in Dysfunctional Tear Syndrome. Am. J. Ophthalmol. 2009, 147, 198–205.e1. [Google Scholar] [CrossRef] [PubMed]
- Kimura-Shimmyo, A.; Kashiwamura, S.; Ueda, H.; Ikeda, T.; Kanno, S.; Akira, S.; Nakanishi, K.; Mimura, O.; Okamura, H. Cytokine-induced injury of the lacrimal and salivary glands. J. Immunother. 2002, 25 (Suppl. S1), S42–S51. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Vadlamudi, V.; Toshida, H.; Beuerman, R.W. Loss of parasympathetic innervation leads to sustained expression of pro-inflammatory genes in the rat lacrimal gland. Auton. Neurosci. Basic Clin. 2006, 124, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Törnwall, J.; Lane, T.E.; Fox, R.I.; Fox, H.S. T cell attractant chemokine expression initiates lacrimal gland destruction in nonobese diabetic mice. Lab. Investig. 1999, 79, 1719–1726. [Google Scholar] [PubMed]


 palmitoleate (POA). All data represent the mean ± SD of four to five mice. $ p < 0.1, * p < 0.05, ** p < 0.01 versus the vehicle.
palmitoleate (POA). All data represent the mean ± SD of four to five mice. $ p < 0.1, * p < 0.05, ** p < 0.01 versus the vehicle.
palmitoleate (POA). All data represent the mean ± SD of four to five mice. $ p < 0.1, * p < 0.05, ** p < 0.01 versus the vehicle.
palmitoleate (POA). All data represent the mean ± SD of four to five mice. $ p < 0.1, * p < 0.05, ** p < 0.01 versus the vehicle.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Sea-Buckthorn Oil | Olive Oil | |
|---|---|---|---|
| PO | SO | ||
| Palmitate C16:0 | 30.0 | 11.3 | 9.7 |
| Palmitoleate C16:1 | 30.1 | 7.5 | 0.6 |
| Stearate C18 | 0.6 | 2.0 | 3.1 |
| Oleate C18:1 | 28.6 | 18.5 | 78.4 |
| Linolate C18:2 | 2.8 | 22.8 | 6.9 |
| α-Linoleate C18:3 | 1.0 | 21.8 | 0.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, S.; Kimura, Y.; Mori, D.; Imada, T.; Izuta, Y.; Shibuya, M.; Sakaguchi, H.; Oonishi, E.; Okada, N.; Matsumoto, K.; et al. Restoration of Tear Secretion in a Murine Dry Eye Model by Oral Administration of Palmitoleic Acid. Nutrients 2017, 9, 364. https://doi.org/10.3390/nu9040364
Nakamura S, Kimura Y, Mori D, Imada T, Izuta Y, Shibuya M, Sakaguchi H, Oonishi E, Okada N, Matsumoto K, et al. Restoration of Tear Secretion in a Murine Dry Eye Model by Oral Administration of Palmitoleic Acid. Nutrients. 2017; 9(4):364. https://doi.org/10.3390/nu9040364
Chicago/Turabian StyleNakamura, Shigeru, Yuki Kimura, Daisuke Mori, Toshihiro Imada, Yusuke Izuta, Michiko Shibuya, Hisayo Sakaguchi, Erina Oonishi, Naoko Okada, Kenji Matsumoto, and et al. 2017. "Restoration of Tear Secretion in a Murine Dry Eye Model by Oral Administration of Palmitoleic Acid" Nutrients 9, no. 4: 364. https://doi.org/10.3390/nu9040364
APA StyleNakamura, S., Kimura, Y., Mori, D., Imada, T., Izuta, Y., Shibuya, M., Sakaguchi, H., Oonishi, E., Okada, N., Matsumoto, K., & Tsubota, K. (2017). Restoration of Tear Secretion in a Murine Dry Eye Model by Oral Administration of Palmitoleic Acid. Nutrients, 9(4), 364. https://doi.org/10.3390/nu9040364