Genistein Ameliorates Ischemia/Reperfusion-Induced Renal Injury in a SIRT1-Dependent Manner

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals, I/R, Genistein Treatment, and SIRT1 Inhibition
2.2. Biochemical Determinations
2.3. Kidney Histology
2.4. Immunohistochemistry
2.5. TUNEL Assay
2.6. Western Blotting
2.7. Statistical Analysis
3. Results
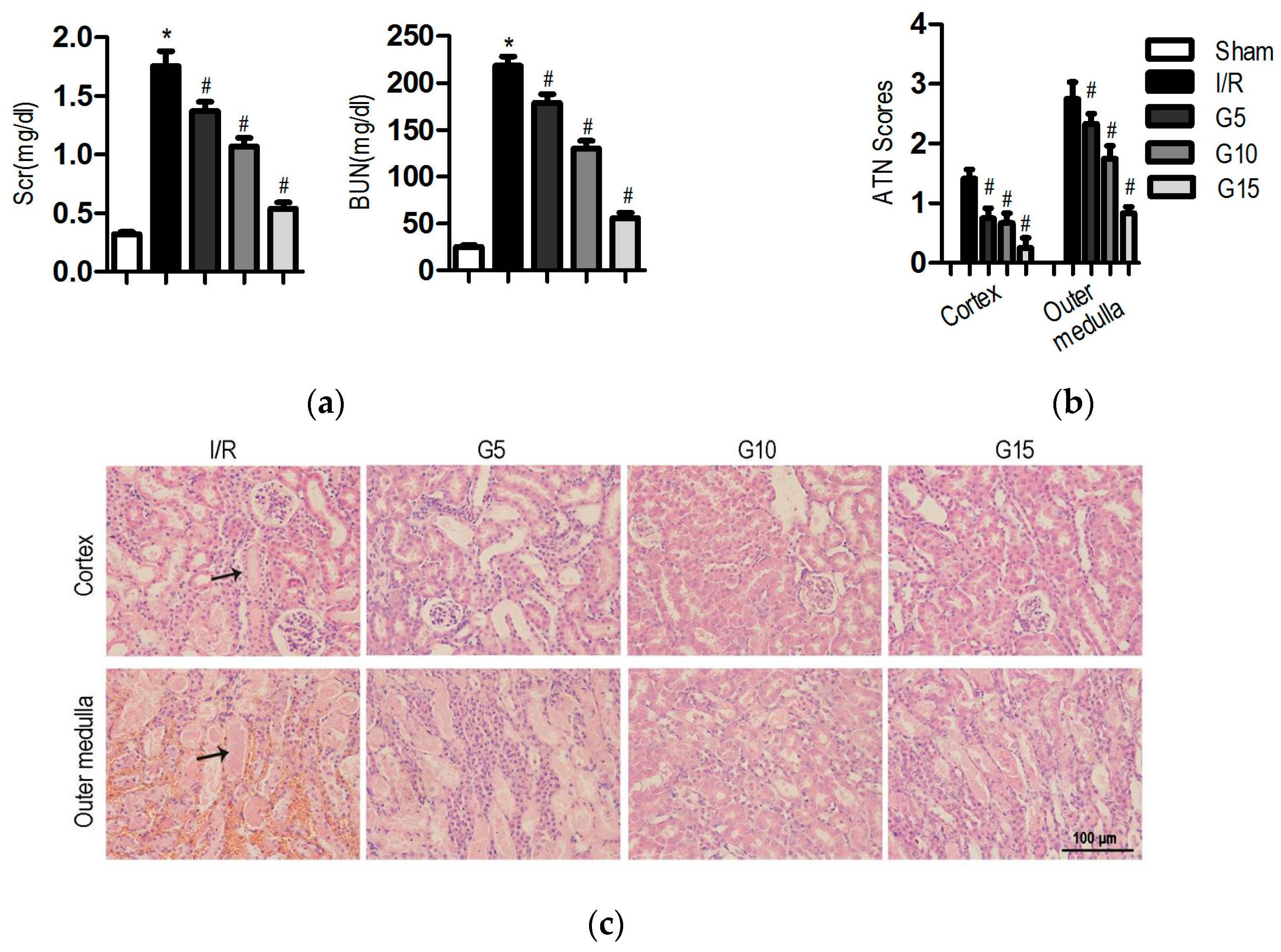
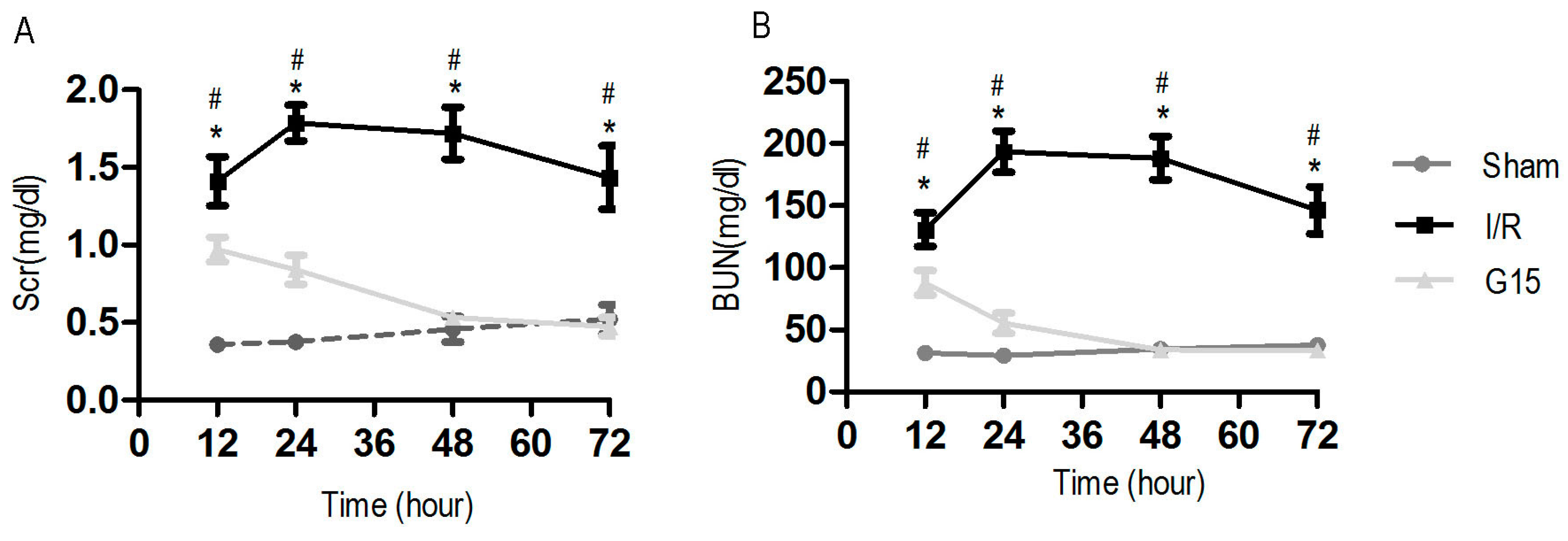
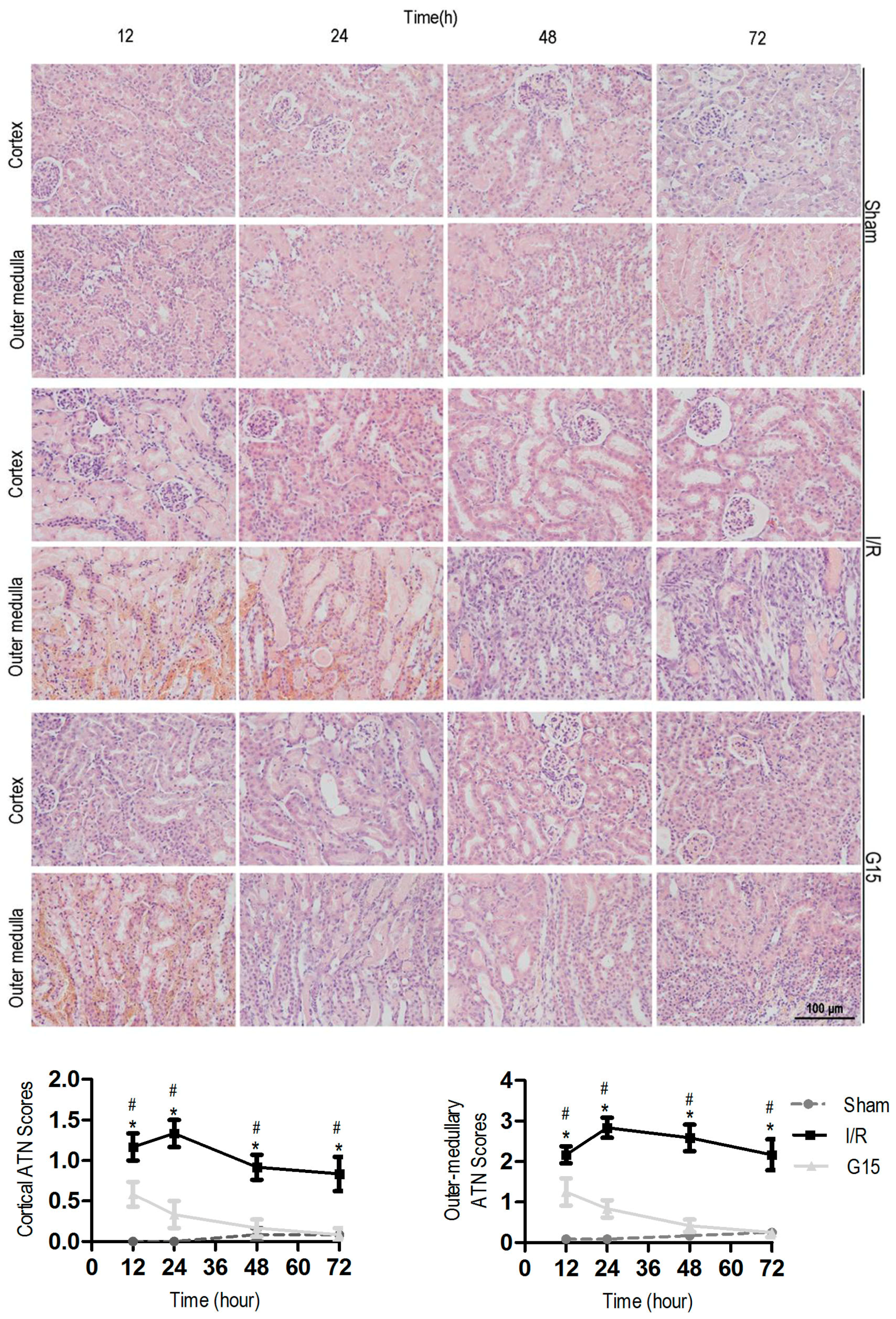
3.1. Genistein Protects the Kidney Against I/R Injury
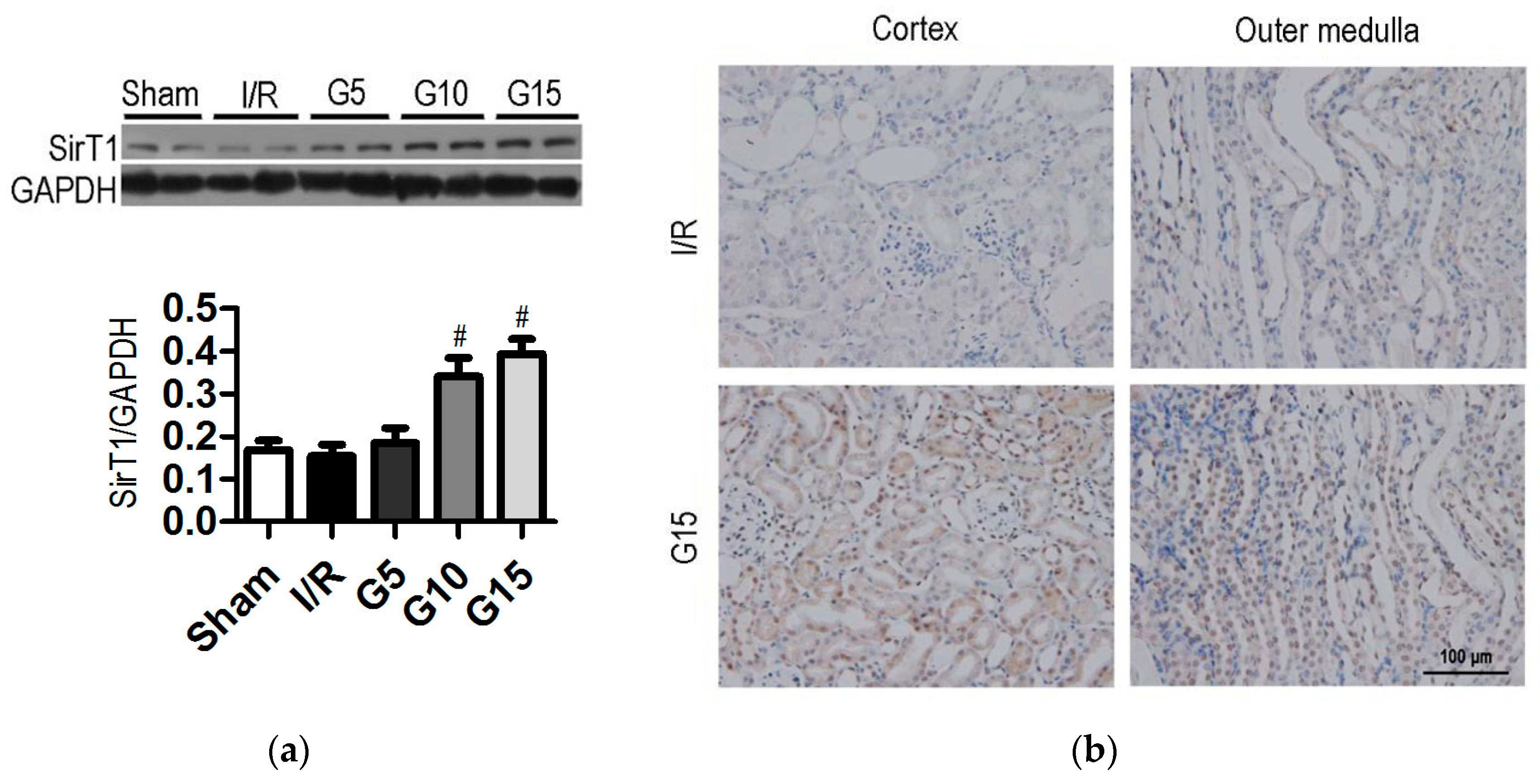
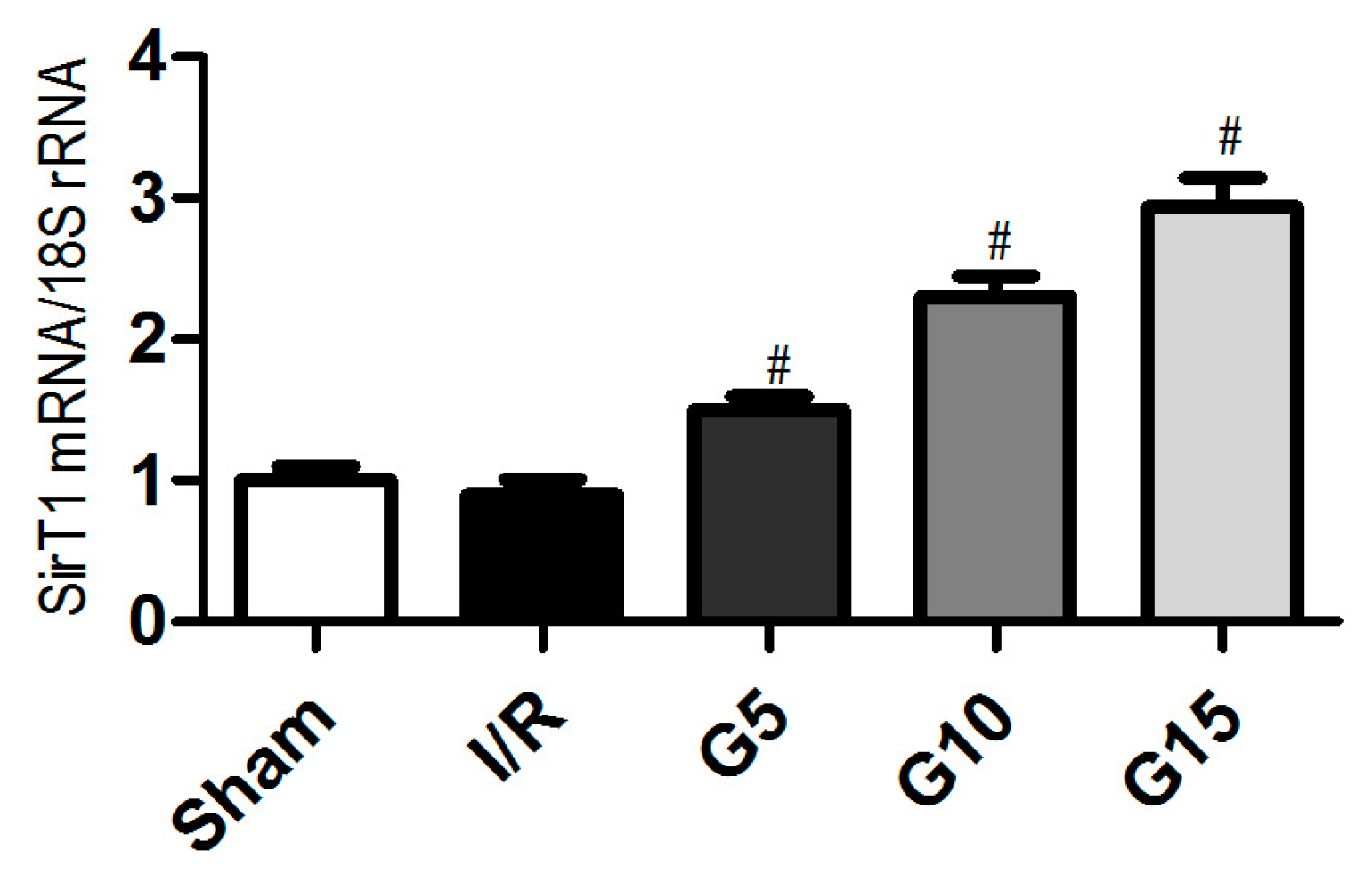
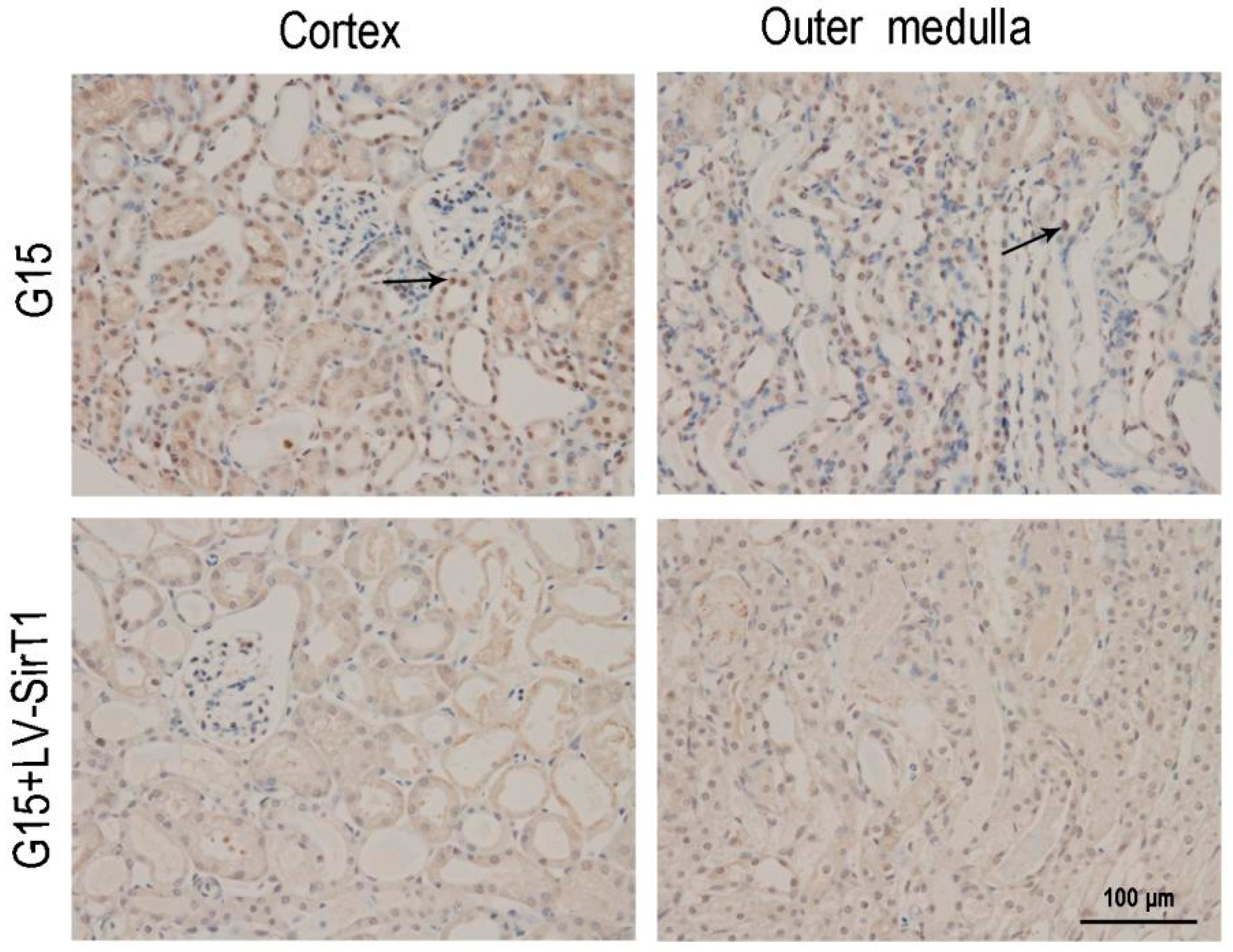
3.2. Genistein Increases SIRT1 Expression in Renal Cells after Renal I/R-Induced Injury
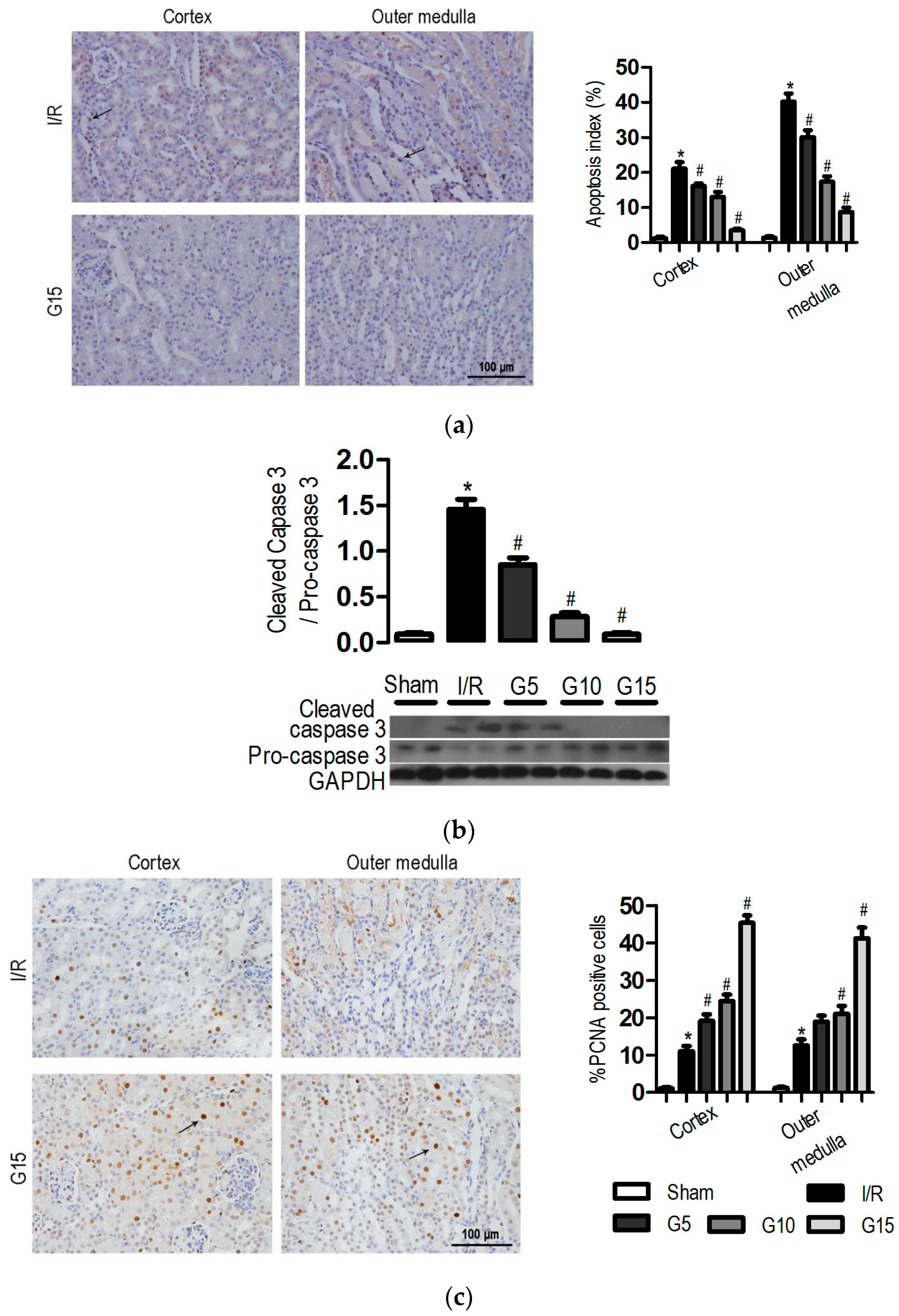
3.3. Genistein Inhibits Apoptosis and Increases Proliferation after Renal I/R-Induced Injury
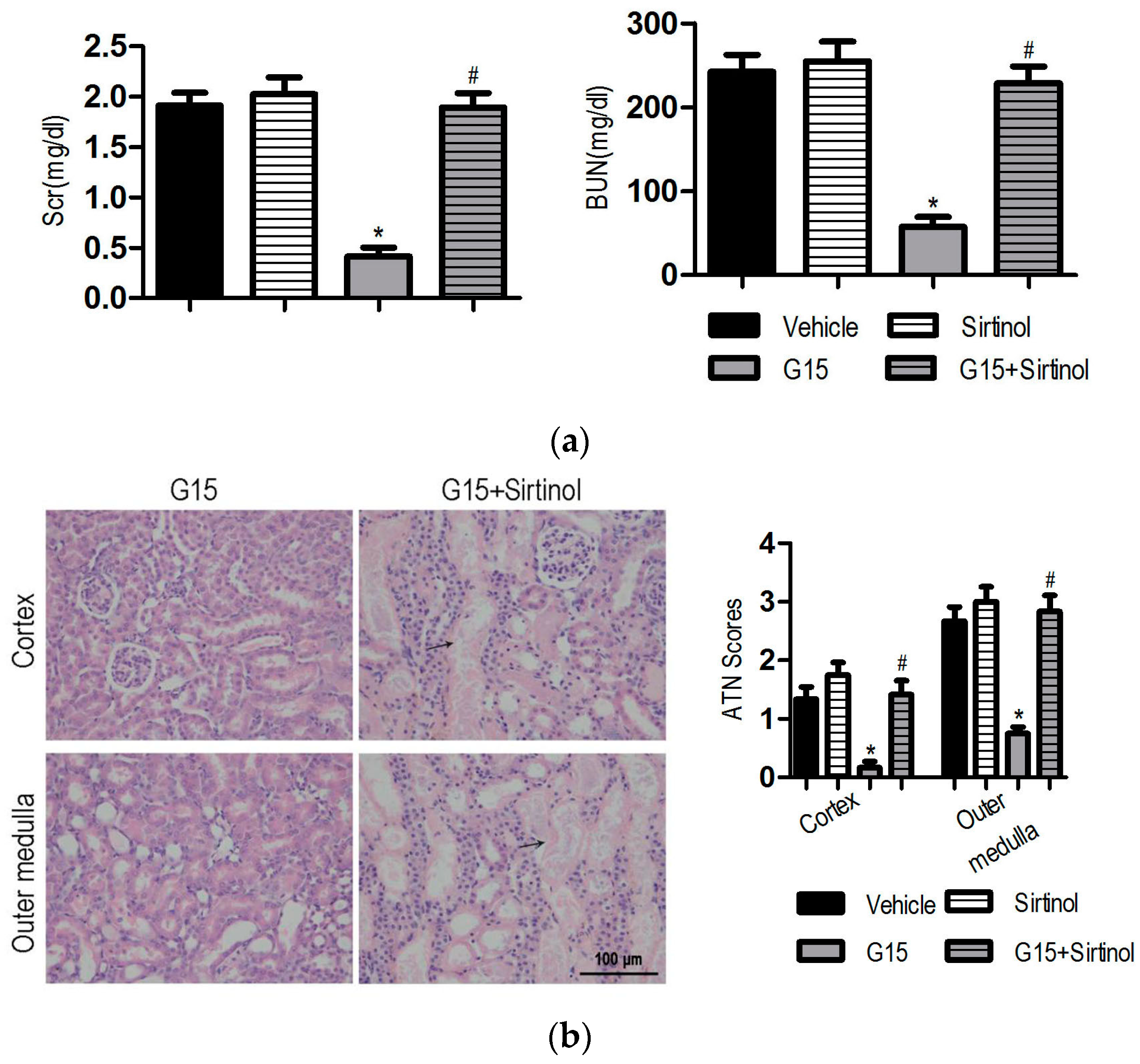
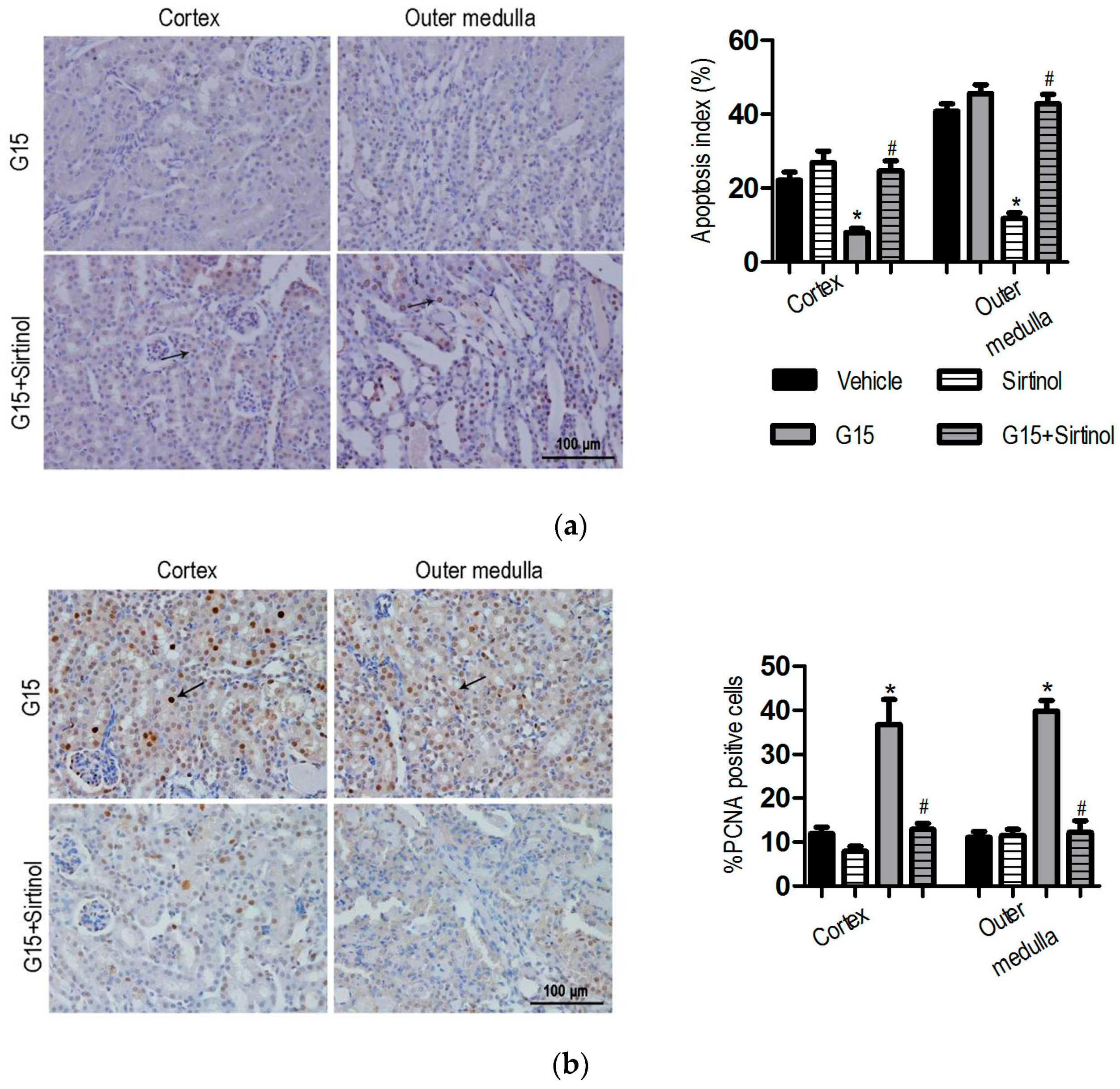
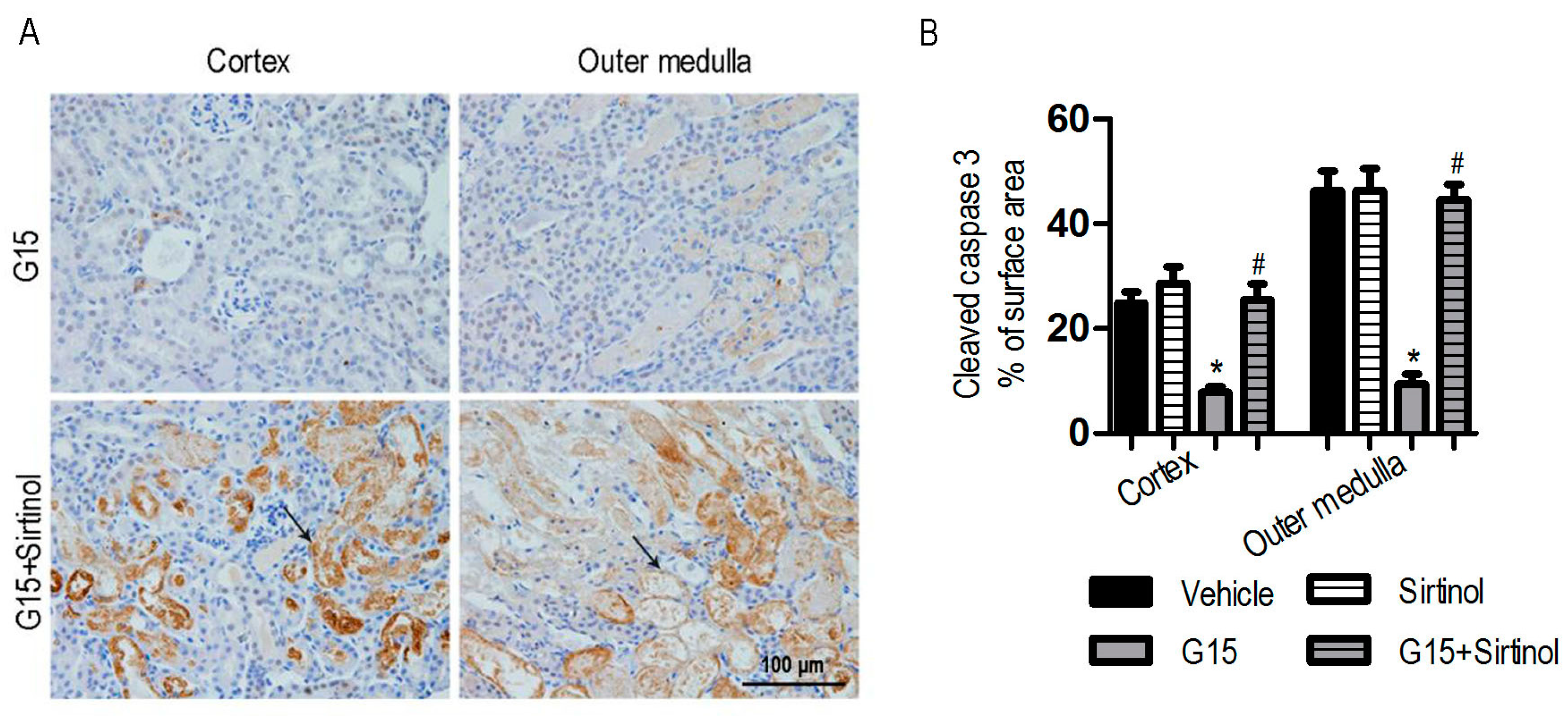
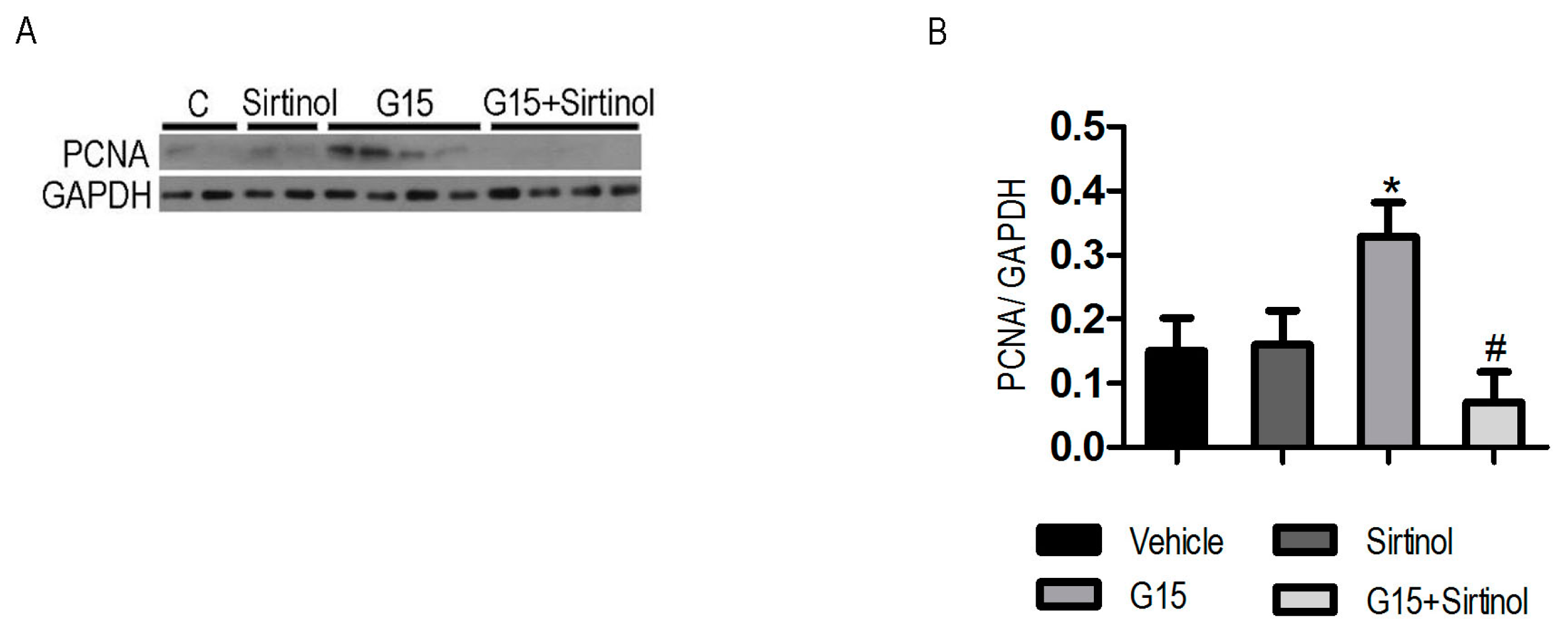
3.4. The SIRT1 Inhibitor Abolishes the Protective Effects of Genistein on I/R-Induced Injury
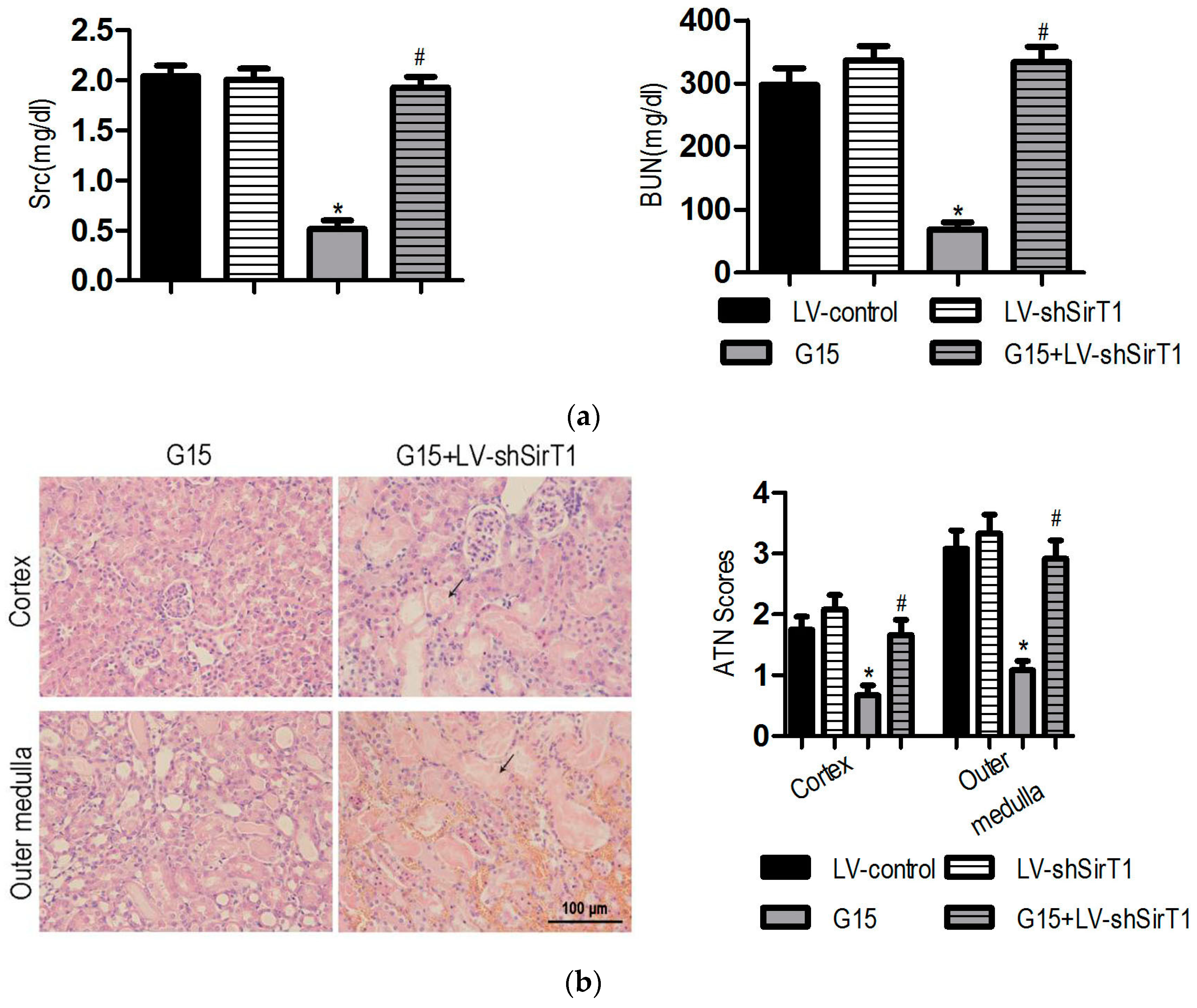
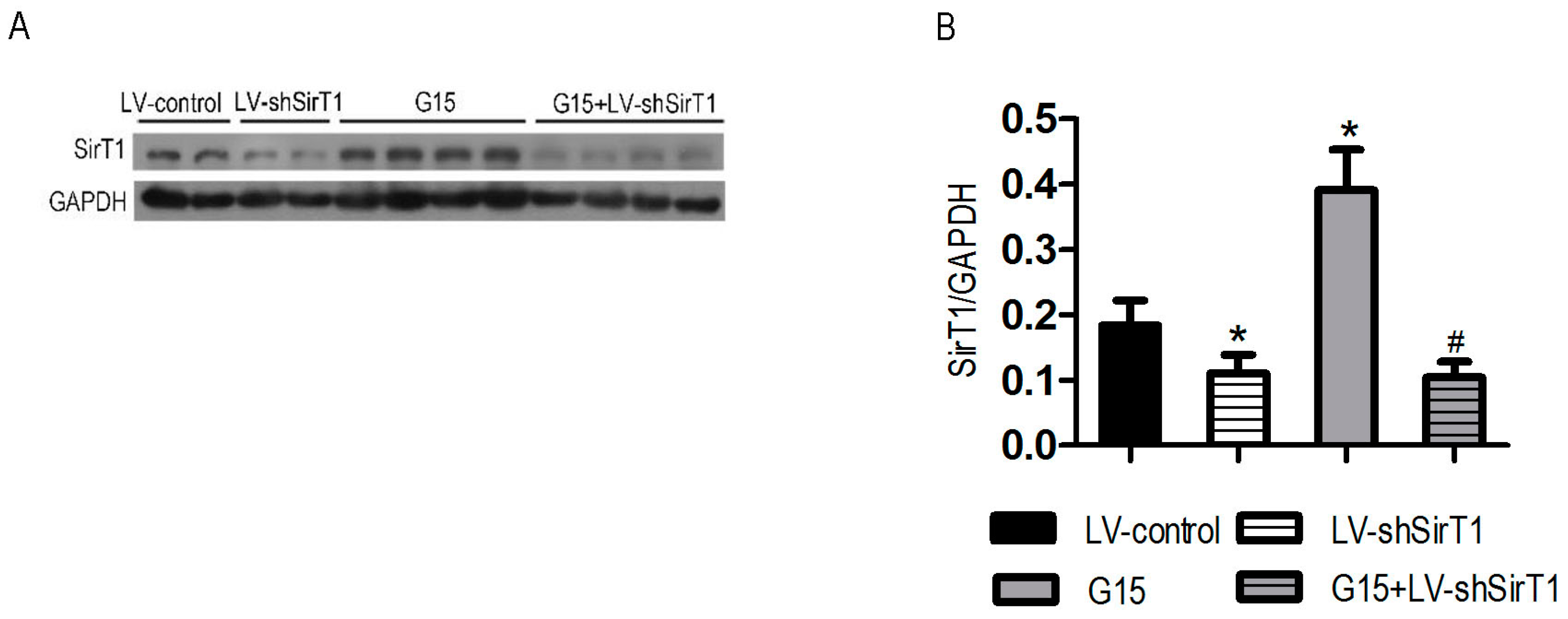
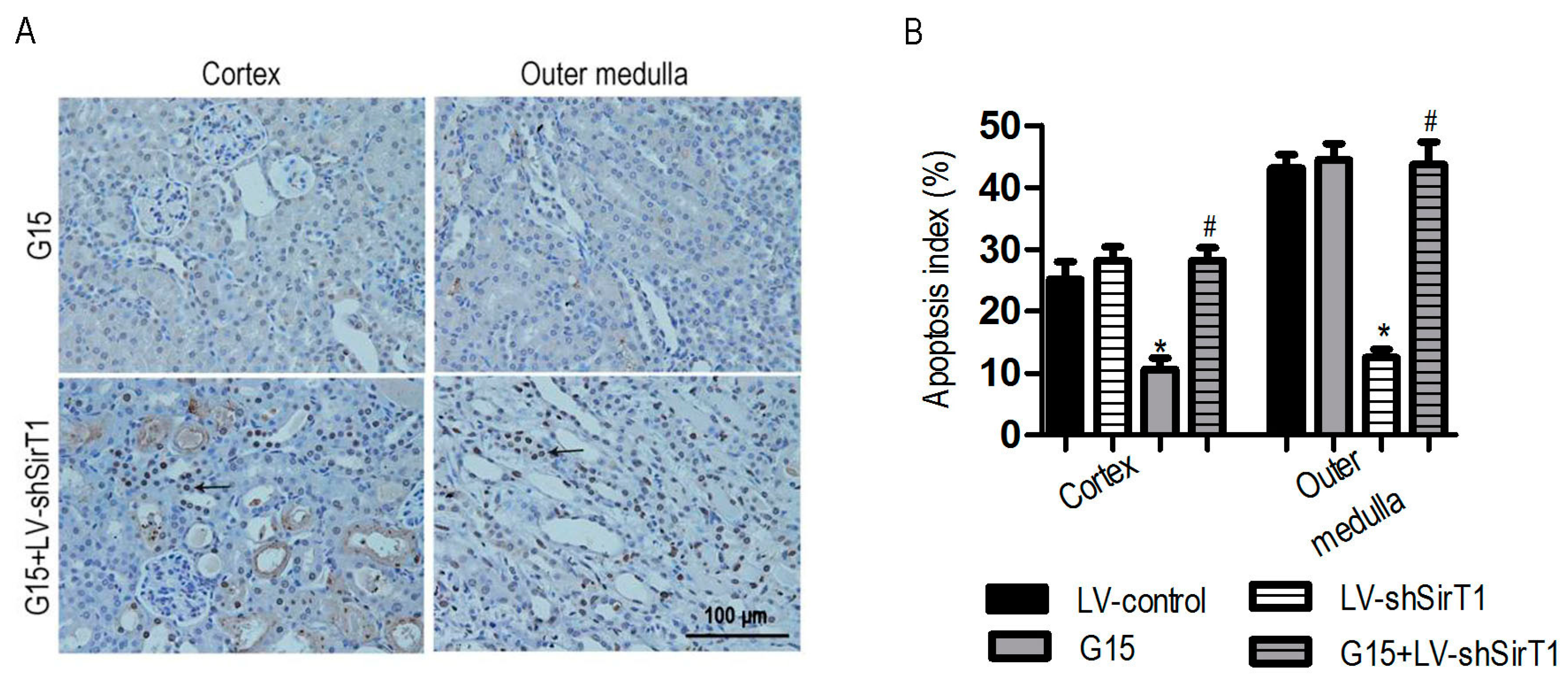
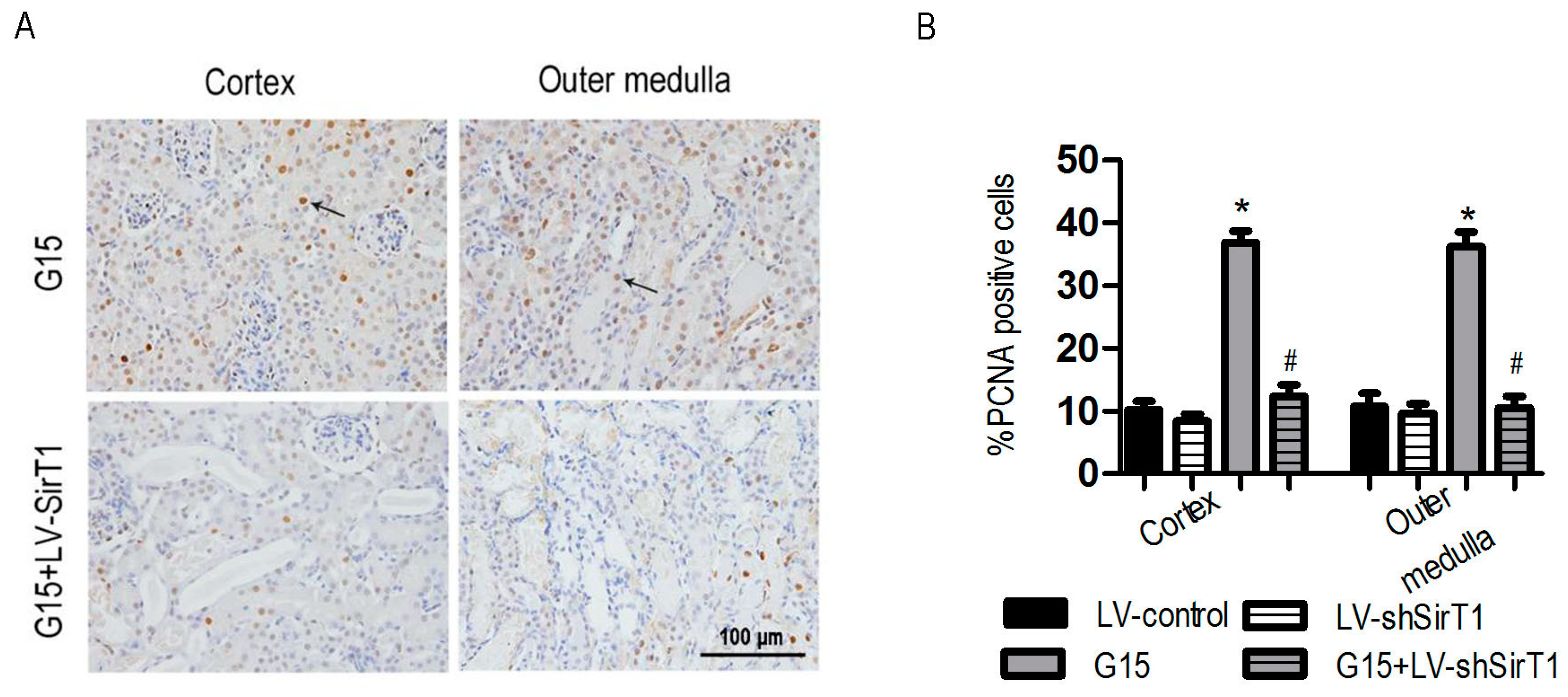
3.5. SIRT1 Depletion Eliminates the Protective Effects of Genistein on I/R-Induced Injury
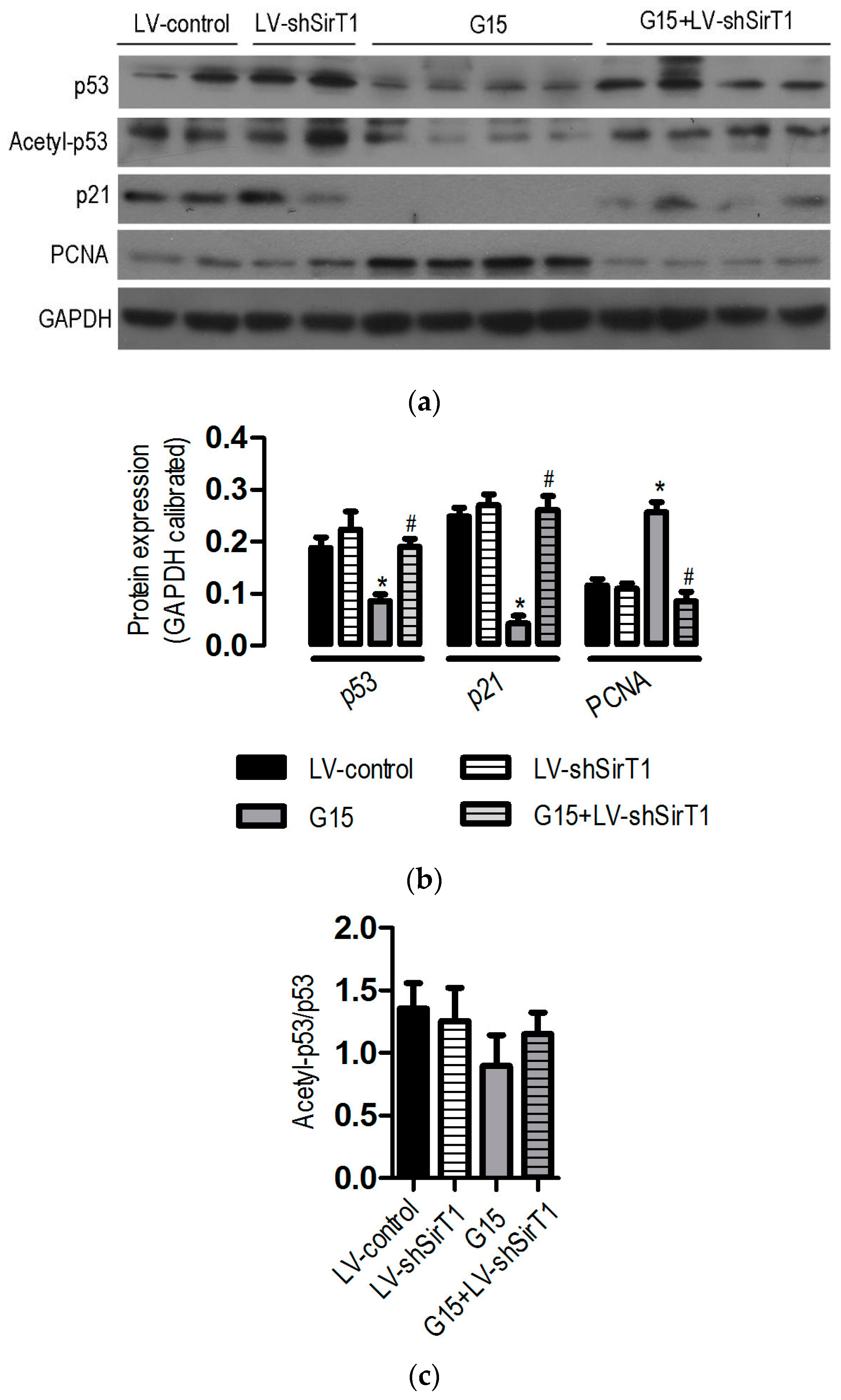
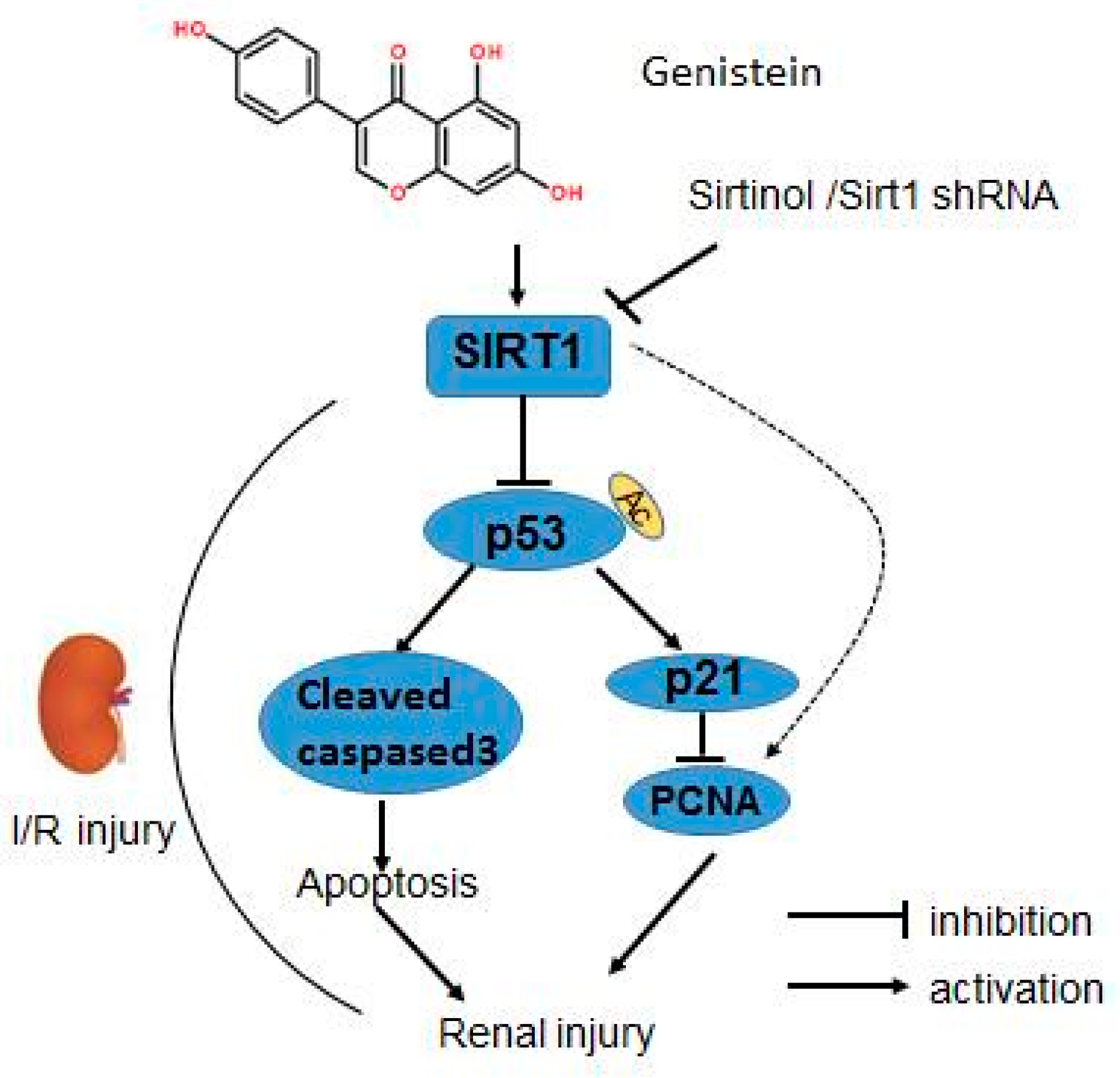
3.6. The Protective Effects of Genistein Is Associated with the SIRT1/p53 Axis
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Appendix A
Appendix A.1. Lentivirus Preparation
Appendix A.2. Quantitative Real-Time RT-PCR
Appendix B









References
- Ichai, C.; Vinsonneau, C.; Souweine, B.; Armando, F.; Canet, E.; Clec’h, C.; Constantin, J.M.; Darmon, M.; Duranteau, J.; Gaillot, T.; et al. Acute kidney injury in the perioperative period and in intensive care units (excluding renal replacement therapies). Ann. Intensive Care 2016, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Waikar, S.S.; Liu, K.D.; Chertow, G.M. Diagnosis, epidemiology and outcomes of acute kidney injury. Clin. J. Am. Soc. Nephrol. 2008, 3, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Gueler, F.; Gwinner, W.; Schwarz, A.; Haller, H. Long-term effects of acute ischemia and reperfusion injury. Kidney Int. 2004, 66, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Lempiainen, J.; Finckenberg, P.; Levijoki, J.; Mervaala, E. AMPK activator AICAR ameliorates ischaemia reperfusion injury in the rat kidney. Br. J. Pharmacol. 2012, 166, 1905–1915. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, T.; Tsutsui, H.; Miura, T.; Nishinaka, T.; Terada, T.; Takama, M.; Yoshida, S.; Tanba, T.; Tojo, A.; Yamagata, M.; et al. Renoprotective effect of yohimbine on ischaemia/reperfusion-induced acute kidney injury through alpha2C-adrenoceptors in rats. Eur. J. Pharmacol. 2016, 781, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.P.; Singh, N.; Bedi, P.M. Pioglitazone ameliorates renal ischemia reperfusion injury through NMDA receptor antagonism in rats. Mol. Cell. Biochem. 2016, 417, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Kelly, G. A review of the Sirtuin system, its clinical implications, and the potential role of dietary activators like resveratrol: Part 1. Altern. Med. Rev. 2010, 15, 245–263. [Google Scholar] [PubMed]
- Kumar, A.; Chauhan, S. How much successful are the medicinal chemists in modulation of SIRT1: A critical review. Eur. J. Med. Chem. 2016, 119, 45–69. [Google Scholar] [CrossRef] [PubMed]
- Wakino, S.; Hasegawa, K.; Itoh, H. Sirtuin and metabolic kidney disease. Kidney Int. 2015, 88, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Wakino, S.; Simic, P.; Sakamaki, Y.; Minakuchi, H.; Fujimura, K.; Hosoya, K.; Komatsu, M.; Kaneko, Y.; Kanda, T.; et al. Renal tubular SIRT1 attenuates diabetic albuminuria by epigenetically suppressing Claudin-1 overexpression in podocytes. Nat. Med. 2013, 19, 1496–1504. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, Y.; Zhang, M.Z.; You, L.; Davis, L.S.; Fan, H.; Yang, H.C.; Fogo, A.B.; Zent, R.; Harris, R.C. SIRT1 activation protects the mouse renal medulla from oxidative injury. J. Clin. Investig. 2010, 120, 1056–1068. [Google Scholar] [CrossRef] [PubMed]
- Kume, S.; Haneda, M.; Kanasaki, K.; Sugimoto, T.; Araki, S.; Isono, M.; Isshiki, K.; Uzu, T.; Kashiwagi, A.; Koya, D. Silent information regulator 2 (SIRT1) attenuates oxidative stress-induced mesangial cell apoptosis via p53 deacetylation. Free. Radic. Biol. Med. 2006, 40, 2175–2182. [Google Scholar] [CrossRef] [PubMed]
- Kume, S.; Haneda, M.; Kanasaki, K.; Sugimoto, T.; Araki, S.; Isshiki, K.; Isono, M.; Uzu, T.; Guarente, L.; Kashiwagi, A.; et al. SIRT1 inhibits transforming growth factor beta-induced apoptosis in glomerular mesangial cells via Smad7 deacetylation. J. Biol. Chem. 2007, 282, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.; Yang, H.C.; You, L.; Wang, Y.Y.; He, W.J.; Hao, C.M. The histone deacetylase, SIRT1, contributes to the resistance of young mice to ischemia/reperfusion-induced acute kidney injury. Kidney Int. 2013, 83, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, Y.; Wei, Q.; Huo, Y.; Liu, K.; Liu, F.; Dong, Z. Tubular p53 regulates multiple genes to mediate AKI. J. Am. Soc. Nephrol. 2014, 25, 2278–2289. [Google Scholar] [CrossRef] [PubMed]
- Ying, Y.; Kim, J.; Westphal, S.N.; Long, K.E.; Padanilam, B.J. Targeted deletion of p53 in the proximal tubule prevents ischemic renal injury. J. Am. Soc. Nephrol. 2014, 25, 2707–2716. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.R.; Verweij, M.; Brand, K.; van de Ven, M.; Goemaere, N.; van den Engel, S.; Chu, T.; Forrer, F.; Müller, C.; de Jong, M.; et al. Short-term dietary restriction and fasting precondition against ischemia reperfusion injury in mice. Aging Cell 2010, 9, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Li, Y.; Wang, Z.; Sarkar, F.H. Multi-targeted therapy of cancer by genistein. Cancer Lett. 2008, 269, 226–242. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Wan, C.; Luo, Q.; Huang, Z.; Luo, Q. Genistein-inhibited cancer stem cell-like properties and reduced chemoresistance of gastric cancer. Int. J. Mol. Sci. 2014, 15, 3432–3443. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.W.; Qiu, S.F.; Shen, J.; Sun, L.N.; Wang, J.Y.; Bruce, I.C.; Xia, Q. Genistein attenuates oxidative stress and neuronal damage following transient global cerebral ischemia in rat hippocampus. Neurosci. Lett. 2008, 438, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Itagaki, S.; Oikawa, S.; Ogura, J.; Kobayashi, M.; Hirano, T.; Sugawara, M.; Iseki, K. Protective effect of soy isoflavone genistein on ischemia-reperfusion in the rat small intestine. Biol. Pharm. Bull. 2011, 34, 1448–1454. [Google Scholar] [CrossRef] [PubMed]
- Canyilmaz, E.; Uslu, G.H.; Bahat, Z.; Kandaz, M.; Mungan, S.; Haciislamoglu, E.; Mentese, A.; Yoney, A. Comparison of the effects of melatonin and genistein on radiation-induced nephrotoxicity: Results of an experimental study. Biomed. Rep. 2016, 4, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Hirasaka, K.; Maeda, T.; Ikeda, C.; Haruna, M.; Kohno, S.; Abe, T.; Ochi, A.; Mukai, R.; Oarada, M.; Eshima-Kondo, S.; et al. Isoflavones derived from soy beans prevent MuRF1-mediated muscle atrophy in C2C12 myotubes through SIRT1 activation. J. Nutr. Sci. Vitaminol. 2013, 59, 317–324. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Lu, M.; Ma, L.; Zhang, S.; Qiu, M.; Wang, Y. Osthole ameliorates renal ischemia-reperfusion injury in rats. J. Surg. Res. 2013, 183, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Bellner, L.; Zhao, H.; Ratliff, B.B.; Darzynkiewicz, Z.; Vio, C.P.; Ferreri, N.R. NFAT5 is protective against ischemic acute kidney injury. Hypertension 2014, 63, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Yao, H.; Caito, S.; Hwang, J.W.; Arunachalam, G.; Rahman, I. Regulation of SIRT1 in cellular functions: Role of polyphenols. Arch. Biochem. Biophys. 2010, 501, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Havasi, A.; Borkan, S.C. Apoptosis and acute kidney injury. Kidney Int. 2011, 80, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.T.; Gallos, G.; Nasr, S.H.; Emala, C.W. A1 adenosine receptor activation inhibits inflammation, necrosis, and apoptosis after renal ischemia-reperfusion injury in mice. J. Am. Soc. Nephrol. 2004, 15, 102–111. [Google Scholar] [CrossRef] [PubMed]
- Migita, H.; Yoshitake, S.; Tange, Y.; Choijookhuu, N.; Hishikawa, Y. Hyperbaric Oxygen Therapy Suppresses Apoptosis and Promotes Renal Tubular Regeneration after Renal Ischemia/Reperfusion Injury in Rats. Nephro-Urol. Mon. 2016, 8, e34421. [Google Scholar] [CrossRef] [PubMed]
- Cortina, B.; Torregrosa, G.; Castelló-Ruiz, M.; Burguete, M.C.; Moscardó, A.; Latorre, A.; Salom, J.B.; Vallés, J.; Santos, M.T.; Alborch, E. Improvement of the circulatory function partially accounts for the neuroprotective action of the phytoestrogen genistein in experimental ischemic stroke. Eur. J. Pharmacol. 2013, 708, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Imai, S. The NAD World: A new systemic regulatory network for metabolism and aging—SIRT1, systemic NAD biosynthesis, and their importance. Cell Biochem. Biophys. 2009, 53, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Wakino, S.; Yoshioka, K.; Tatematsu, S.; Hara, Y.; Minakuchi, H.; Sueyasu, K.; Washida, N.; Tokuyama, H.; Tzukerman, M.; et al. Kidney-specific overexpression of SIRT1 protects against acute kidney injury by retaining peroxisome function. J. Biol. Chem. 2010, 285, 13045–13056. [Google Scholar] [CrossRef] [PubMed]
- Nadal-Serrano, M.; Pons, D.G.; Sastre-Serra, J.; Blanquer-Rosselló Mdel, M.; Roca, P.; Oliver, J. Genistein modulates oxidative stress in breast cancer cell lines according to ERalpha/ERbeta ratio: Effects on mitochondrial functionality, Sirtuins, uncoupling protein 2 and antioxidant enzymes. Int. J. Biochem. Cell Biol. 2013, 45, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Hsu, M.H.; Savas, U.; Lasker, J.M.; Johnson, E.F. Genistein, resveratrol, and 5-aminoimidazole-4-carboxamide-1-beta-d-ribofuranoside induce cytochrome P450 4F2 expression through an AMP-activated protein kinase-dependent pathway. J. Pharmacol. Exp. Ther. 2011, 337, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Kikuno, N.; Shiina, H.; Urakami, S.; Kawamoto, K.; Hirata, H.; Tanaka, Y.; Majid, S.; Igawa, M.; Dahiya, R. Genistein mediated histone acetylation and demethylation activates tumor suppressor genes in prostate cancer cells. Int. J. Cancer 2008, 123, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, C.; Liu, B.; Jin, X.; Li, P.; Zheng, X.; Zhao, T.; Li, F.; Li, Q. Genistein mediates the selective radiosensitizing effect in NSCLC A549 cells via inhibiting methylation of the keap1 gene promoter region. Oncotarget 2016, 7, 27267–27279. [Google Scholar] [CrossRef] [PubMed]
- Sung, M.J.; Kim, D.H.; Jung, Y.J.; Kang, K.P.; Lee, A.S.; Lee, S.; Kim, W.; Davaatseren, M.; Hwang, J.T.; Kim, H.J.; et al. Genistein protects the kidney from cisplatin-induced injury. Kidney Int. 2008, 74, 1538–1547. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Trotman, L.C.; Shaffer, D.; Lin, H.K.; Dotan, Z.A.; Niki, M.; Koutcher, J.A.; Scher, H.I.; Ludwig, T.; Gerald, W.; et al. Crucial role of p53-dependent cellular senescence in suppression of Pten-deficient tumorigenesis. Nature 2005, 436, 725–730. [Google Scholar] [CrossRef] [PubMed]
- Rasbach, K.A.; Schnellmann, R.G. Isoflavones promote mitochondrial biogenesis. J. Pharmacol. Exp. Ther. 2008, 325, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Witko-Sarsat, V.; Ohayon, D. Proliferating cell nuclear antigen in neutrophil fate. Immunol. Rev. 2016, 273, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Reiser, J. Production and concentration of pseudotyped HIV-1-based gene transfer vectors. Gene Ther. 2000, 7, 910–913. [Google Scholar] [CrossRef] [PubMed]








© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.-F.; Yang, K.; Zhu, P.; Zhao, H.-Q.; Song, Y.-H.; Liu, K.-C.; Huang, W.-F. Genistein Ameliorates Ischemia/Reperfusion-Induced Renal Injury in a SIRT1-Dependent Manner. Nutrients 2017, 9, 403. https://doi.org/10.3390/nu9040403
Li W-F, Yang K, Zhu P, Zhao H-Q, Song Y-H, Liu K-C, Huang W-F. Genistein Ameliorates Ischemia/Reperfusion-Induced Renal Injury in a SIRT1-Dependent Manner. Nutrients. 2017; 9(4):403. https://doi.org/10.3390/nu9040403
Chicago/Turabian StyleLi, Wei-Fang, Kang Yang, Ping Zhu, Hong-Qian Zhao, Yin-Hong Song, Kuan-Can Liu, and Wei-Feng Huang. 2017. "Genistein Ameliorates Ischemia/Reperfusion-Induced Renal Injury in a SIRT1-Dependent Manner" Nutrients 9, no. 4: 403. https://doi.org/10.3390/nu9040403
APA StyleLi, W.-F., Yang, K., Zhu, P., Zhao, H.-Q., Song, Y.-H., Liu, K.-C., & Huang, W.-F. (2017). Genistein Ameliorates Ischemia/Reperfusion-Induced Renal Injury in a SIRT1-Dependent Manner. Nutrients, 9(4), 403. https://doi.org/10.3390/nu9040403