Genetic Variants Involved in One-Carbon Metabolism: Polymorphism Frequencies and Differences in Homocysteine Concentrations in the Folic Acid Fortification Era

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study and Sample Population
2.2. Data Collection and Processing
2.3. Diet
2.4. Anthropometric Measures
2.5. Blood Collection
2.6. Biochemical Analysis
2.7. DNA Extraction and Genotyping
2.8. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ulrich, C.M. Nutrigenetics in cancer research—Folate metabolism and colorectal cancer. J. Nutr. 2005, 135, 2698–2702. [Google Scholar] [PubMed]
- Stover, P.J. Polymorphisms in 1-carbon metabolism, epigenetics and folate-related pathologies. J. Nutr. Nutr. 2011, 4, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.T.; Schalinske, K.L. New insights into the regulation of methyl group and homocysteine metabolism. J. Nutr. 2007, 137, 311–314. [Google Scholar] [PubMed]
- Eldibany, M.M.; Caprini, J.A. Hyperhomocysteinemia and thrombosis: An overview. Arch. Pathol. Lab. Med. 2007, 131, 872–884. [Google Scholar] [PubMed]
- Clarke, R.; Halsey, J.; Lewington, S.; Lonn, E.; Armitage, J.; Manson, J.E.; Bønaa, K.H.; Spence, J.D.; Nygård, O.; Jamison, R.; et al. Effects of lowering homocysteine levels with B vitamins on cardiovascular disease, cancer, and cause-specific mortality: Meta-analysis of 8 randomized trials involving 37 485 individuals. Arch. Intern. Med. 2010, 170, 1622–1631. [Google Scholar] [PubMed]
- DeVos, L.; Chanson, A.; Liu, Z.; Ciappio, E.D.; Parnell, L.D.; Mason, J.B.; Tucker, K.L.; Crott, J.W. Associations between single nucleotide polymorphisms in folate uptake and metabolizing genes with blood folate, homocysteine, and DNA uracil concentrations. Am. J. Clin. Nutr. 2008, 88, 1149–1158. [Google Scholar] [PubMed]
- Jacques, P.F.; Selhub, J.; Bostom, A.G.; Wilson, P.W.F.; Rosenberg, I.H. The effect of folic acid fortification on plasma folate and total homocysteine concentrations. N. Engl. J. Med. 1999, 340, 1449–1454. [Google Scholar] [CrossRef] [PubMed]
- Ganji, V.; Kafai, M.R. Trends in serum folate, RBC folate, and circulating total homocysteine concentrations in the United States: Analysis of data from National Health and Nutrition Examination Surveys, 1988–1994, 1999–2000, and 2001–2002. J. Nutr. 2006, 136, 153–158. [Google Scholar] [PubMed]
- Marchioni, D.M.L.; Fisberg, R.M. Manual de Avaliação do Consumo Alimentar em Estudos Populacionais: A Experiência do Inquérito de Saúde em São Paulo (ISA); Faculdade de Saúde Pública da USP: São Paulo, Brazil, 2012. [Google Scholar]
- Verly, E., Jr.; Steluti, J.; Fisberg, R.M.; Marchioni, D.M. A quantile regression approach can reveal the effect of fruit and vegetable consumption on plasma homocysteine levels. PLoS ONE 2014, 9, e111619. [Google Scholar] [CrossRef]
- Haubrock, J.; Noethlings, U.; Volatier, J.L.; Dekkers, A.; Ocke, M.; Harttig, U.; Illner, A.K.; Knüppel, S.; Andersen, L.F.; Boeing, H.; et al. Estimating Usual Food Intake Distributions by Using the Multiple Source Method in the EPIC-Potsdam Calibration Study. J. Nutr. 2011, 141, 914–920. [Google Scholar]
- WHO—World Health Organization. Obesity: Preventing and Managing the Global Epidemic Report of a World Health Organization Consultation; World Health Organization: Geneva, Switzerland, 2000; p. 256. [Google Scholar]
- Bagley, P.J.; Selhub, J. Analysis of folate from distribution by affinity followed by reversed-phase chromatography with electrical detection. Clin. Chem. 2000, 46, 404–411. [Google Scholar] [PubMed]
- Rybak, M.E.; Jain, R.B.; Pfeiffer, C.M. Clinical vitamin B6 analysis: An interlaboratory comparison of pyridoxal 5′-phosphate measurements in serum. Clin. Chem. 2005, 51, 1223–1231. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.; Dykes, D.; Polesky, H. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [PubMed]
- Myakishev, M.V.; Khripin, Y.; Hu, S.; Hamer, D.H. High-throughput SNP genotyping by allele-specific PCR with universal energy-transfer-labeled primers. Genome Res. 2001, 11, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Schneider, J.A.; Rees, D.C.; Liu, Y.T.; Clegg, J.B. Worldwide distribution of a common methylenetetrahydrofolate reductase mutation. Am. J. Hum. Genet. 1998, 62, 1258–1260. [Google Scholar] [CrossRef] [PubMed]
- Botto, L.D.; Yang, Q. 5,10-Methylenetetrahydrofolate reductase gene variants and congenital anomalies: A HuGE review. Am. J. Epidemiol. 2000, 151, 862–877. [Google Scholar] [CrossRef] [PubMed]
- Shinjo, S.K.; Oba-Shinjo, S.M.; da Silva, R.; Barbosa, K.C.; Yamamoto, F.; Scaff, M. Methylenetetrahydrofolate reductase gene polymorphism is not related to the risk of ischemic cerebrovascular disease in a Brazilian population. Clinics 2007, 63, 295–300. [Google Scholar] [CrossRef]
- Voetsch, B.; Damasceno, B.P.; Camargo, E.C.; Massaro, A.; Bacheschi, L.A.; Scaff, M.; Annichino-Bizzacchi, J.M.; Arruda, V.R. Inherited thrombophilia as a risk factor for the development of ischaemic stroke in young adults. Thromb. Haemost. 2000, 83, 229–233. [Google Scholar] [PubMed]
- Oliveira, K.C.; Bianco, B.; Verreschi, I.T.N.; Guedes, A.D.; Galera, B.B.; Galera, M.F.; Barbosa, C.P.; Lipay, M.V. Prevalence of the Polymorphism MTHFR A1298C and not MTHFR C677T Is Related to Chromosomal Aneuploidy in Brazilian Turner Syndrome Patients. Arq. Bras. Endocrinol. Metabol. 2008, 52, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Devlin, A.M.; Clarke, R.; Birks, J.; Evans, J.G.; Halsted, C.H. Interactions among polymorphisms in folate-metabolizing genes and serum total homocysteine concentrations in a healthy elderly population. Am. J. Clin. Nutr. 2006, 3, 708–713. [Google Scholar]
- Lima, C.S.; Ortega, M.M.; Ozelo, M.C.; Araujo, R.C.; De Souza, C.A.; Lorand-Metze, I.; Annichino-Bizzacchi, J.M.; Costa, F.F. Polymorphisms of methylenetetrahydrofolate reductase (MTHFR), methionine synthase (MTR), methionine synthase reductase (MTRR), and thymidylate synthase (TYMS) in multiple myeloma risk. Leuk. Res. 2008, 32, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Sharp, L.; Little, J. Polymorphisms in genes involved in folate metabolism and colorectal neoplasia: A HuGE review. Am. J. Epidemiol. 2004, 159, 423–443. [Google Scholar] [CrossRef] [PubMed]
- Barnabé, A.; Aléssio, A.C.; Bittar, L.F.; de MoraesMazetto, B.; Bicudo, A.M.; de Paula, E.V.; Höehr, N.F.; Annichino-Bizzacchi, J.M. Folate, vitamin B12 and Homocysteine status in the post-folic acid fortification era in different subgroups of the Brazilian population attended to at a public health care center. Nutr. J. 2015, 14, 19. [Google Scholar] [CrossRef] [PubMed]
- Kalmbach, R.D.; Choumenkovitch, S.F.; Troen, A.P.; Jacques, P.F.; D’Agostino, R.; Selhub, J. A 19-base pair deletion polymorphism in dihydrofolate reductase is associated with increased unmetabolized folic acid in plasma and decreased red blood cell folate. J. Nutr. 2008, 138, 2323–2327. [Google Scholar] [CrossRef] [PubMed]
- Mendes, C.C.; Raimundo, A.M.; Oliveira, L.D.; Zampieri, B.L.; Marucci, G.H.; Biselli, J.M.; Goloni-Bertollo, E.M.; Eberlin, M.N.; Haddad, R.; Riccio, M.F.; et al. DHFR 19-bp Deletion and SHMT C1420T Polymorphisms and Metabolite Concentrations of the Folate Pathway in Individuals with Down Syndrome. Genet. Test. Mol. Biomark. 2013, 17, 274–277. [Google Scholar] [CrossRef] [PubMed]
- Hoey, L.; McNulty, H.; Askin, N.; Dunne, A.; Ward, M.; Pentieva, K.; Strain, J.; Molloy, A.M.; Flynn, C.A.; Scott, J.M. Effect of a voluntary food fortification policy on folate, related B vitamin status, and homocysteine in healthy adults. Am. J. Clin. Nutr. 2007, 86, 1405–1413. [Google Scholar] [PubMed]
- Girelli, D.; Friso, S.; Trabetti, E.; Olivieri, O.; Russo, C.; Pessotto, R.; Faccini, G.; Pignatti, P.F.; Mazzucco, A.; Corrocher, R. Methylenetetrahydrofolate reductase C677T mutation plasma homocysteine, and folate in subjects from northern Italy with or without angiographically documented severe coronary atherosclerotic disease: Evidence for an important genetic-environmental interaction. Blood 1998, 9, 4158–4163. [Google Scholar]
- Crider, K.S.; Zhu, J.H.; Hao, L.; Yang, Q.H.; Yang, T.P.; Gindler, J.; Maneval, D.R.; Quinlivan, E.P.; Li, Z.; Bailey, L.B.; et al. MTHFR 677C→T genotype is associated with folate and homocysteine concentrations in a large, population-based, double-blind trial of folic acid supplementation. Am. J. Clin. Nutr. 2011, 93, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Rozen, R. Genetic predisposition to hyperhomocysteinemia: Deficiency of methylenetetrahydrofolate reductase (MTHFR). Thromb. Haemost. 1997, 78, 523–526. [Google Scholar] [PubMed]
- Marchioni, D.M.L.; Verly, E., Jr.; Steluti, J.; Cesar, C.L.G.; Fisberg, R.M. Ingestão de folato nos períodos pré e pós-fortificação mandatória: Estudo de base populacional em São Paulo, Brasil Folic acid intake before and after mandatory fortification: A population-based study in Sao Paulo, Brazil. Cad. Saúde Pública 2013, 29, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO—Food and Agriculture Organization/World Health Organization. Folate and Folic Acid. In FAO/WHO Expert Consultation on Human Vitamin and Mineral Requirements; FAO: Rome, Italy, 2001; pp. 53–63. [Google Scholar]
- Parra, F.C.; Amado, R.C.; Lambertucci, J.R.; Rocha, J.; Antunes, C.M.; Pena, S.D. Color and genomic ancestry in Brazilians. Proc. Natl. Acad. Sci. USA 2003, 100, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Kurtz, G. Pharmacogenomics and the genetic diversity of the Brazilian population. Cad. Saude Publica 2009, 25, 1650–1651. [Google Scholar] [CrossRef] [PubMed]
- Rady, P.L.; Szucs, S.; Grady, J.; Hudnall, S.D.; Kellner, L.H.; Nitowsky, H.; Tyring, S.K.; Matalon, R.K. Genetic polymorphisms of methylenetetrahydrofolate reductase (MTHFR) and methionine synthase reductase (MTRR) in ethnic populations in Texas; a report of a novel MTHFR polymorphic site, G1793A. Am. J. Med. Genet. 2002, 107, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.-H.; Botto, L.D.; Gallagher, M.; Friedman, J.; Sanders, C.L.; Koontz, D.; Nikolova, S.; Erickson, J.D.; Steinberg, K. Prevalence and effects of gene-gene and gene-nutrient interactions on serum folate and serum total homocysteine concentrations in the United States: Findings from the third National Health and Nutrition Examination Survey DNA Bank. Am. J. Clin. Nutr. 2008, 88, 232–246. [Google Scholar] [PubMed]
- Pangilinan, F.; Molloy, A.M.; Mills, J.L.; Troendle, J.F.; Parle-McDermott, A.; Signore, C.; O’Leary, V.B.; Chines, P.; Seay, J.M.; Geiler-Samerotte, K.; et al. Evaluation of common genetic variants in 82 candidate genes as risk factors for neural tube defects. BMC Med. Genet. 2012, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Mehlig, K.; Leander, K.; De Faire, U.; Nyberg, F.; Berg, C.; Rosengren, A.; Björck, L.; Zetterberg, H.; Blennow, K.; Tognon, G.; et al. The association between plasma homocysteine and coronary heart disease is modified by the MTHFR 677C>T polymorphism. Heart 2013, 99, 1761–1765. [Google Scholar] [CrossRef] [PubMed]
- Muntjewerff, J.W.; Kahn, R.S.; Blom, H.J.; Heijer, M. Homocysteine, methylenetetrahydrofolate reductase and risk of schizophrenia: A meta-analysis. Mol. Psychiatry 2006, 11, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Safarinejad, M.R.; Shafiei, N.; Safarinejad, N. Relationship between three polymorphisms of Methylenetetrahydrofolate reductase (MTHFR C677T, A1298C, and G1793A) gene and risk of Prostate cancer: A case control study. Prostate 2010, 70, 1645–1657. [Google Scholar] [CrossRef] [PubMed]
- Arslan, S.; Karadayi, S.; Yildirim, M.E.; Ozdemir, O.; Akkurt, I. The association between methylene-tetrahydrofolate reductase gene polymorphism and lung cancer risk. Mol. Biol. Rep. 2011, 38, 991–996. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Chen, J. Association of MHTFR Ala222Val (rs1801133) polymorphism and breast cancer susceptibility: An update meta-analysis based on 51 research studies. Diagn. Pathol. 2012, 7, 171. [Google Scholar] [CrossRef] [PubMed]
- Husemoen, L.L.N.; Skaaby, T.; Jørgensen, T.; Thuesen, B.H.; Fenger, M.; Grarup, N.; Sandholt, C.H.; Hansen, T.; Pedersen, O.; Linneberg, A. MTHFR C677T genotype and cardiovascular risk in a general population without mandatory folic acid fortification. Eur. J. Nutr. 2014, 53, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.; Platt, R.; Wu, Q.; Leclerc, D.; Christensen, B.; Yang, H.; Gravel, R.A.; Rozen, R. A Common Variant in Methionine Synthase Reductase Combined with Low Cobalamin (Vitamin B12) Increases Risk for Spina Bifida. Mol. Genet. Metab. 1999, 67, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Zhou, G.W.; Wang, N.; Wang, Y.J. MTRR A66G polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat. 2010, 124, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.G.; Wang, J.L.; Zhang, J.; Zhao, L.X.; Zhai, G.X.; Xiang, Y.Z.; Chang, P. Reduced folate carrier A80G polymorphism and susceptibility to neural tube defects: A meta-analysis. Gene 2012, 510, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Biselli, J.M.; Zampieri, B.L.; Goloni-Bertollo, E.M.; Haddad., R.; Fonseca, M.F.; Eberlin, M.N.; Vannucchi, H.; Carvalho, V.M.; Pavarino, E.C. Genetic polymorphisms modulate the folate metabolism of Brazilian individuals with Down syndrome. Mol. Biol. Rep. 2012, 39, 9277–9284. [Google Scholar] [CrossRef] [PubMed]
- Laguardia, J. O uso da variável “raça” na pesquisa em saúde. Physis 2004, 4, 197–234. [Google Scholar] [CrossRef]


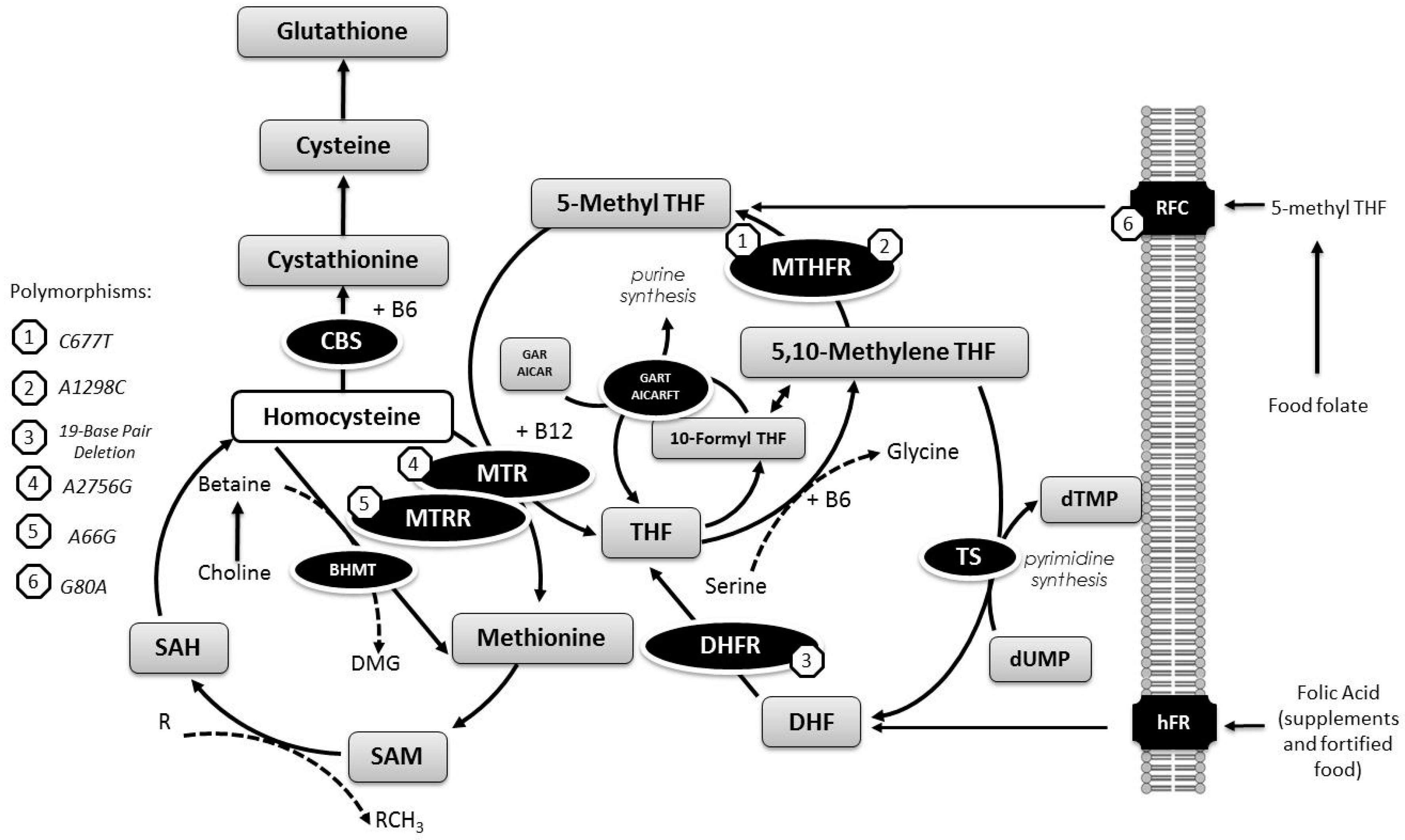{kind=link}
| Polymorphisms | Location | Gene | Changes | p a | MAF | |
|---|---|---|---|---|---|---|
| DNA | Amino acids | |||||
| rs1801131 | 1p36.3 | MTHFR | A→C | Glu→Ala | 0.392 | 0.24 |
| rs1801133 | 1p36.3 | MTHFR | C→T | Ala→Val | 0.428 | 0.33 |
| rs1805087 | 1q43 | MTR | A→G | Asp→Gly | 0.333 | 0.19 |
| rs1801394 | 5p15.31 | MTRR | A→G | - | 0.154 | 0.42 |
| rs1051266 | 21q22.3 | RFC1 | G→A | His→Arg | 0.141 | 0.46 |
| 19-bp deletion | 5q11.2–q13.2 | DHFR | - | - | 0.807 | 0.47 |
| SNP | MTHFR 677C>T | p-Value | MTHFR 1298A>C | p-Value | MTR 2756A>G | p-Value | MTRR 66A>G | p-Value | RFC1 80G>A | p-Value | DHFR Deletion | p-Value | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | C:C | C:T | T:T | A:A | A:C | C:C | A:A | A:G | G:G | A:A | A:G | G:G | G:G | G:A | A:A | WT: WT | WT:del | del:del | ||||||
| Total (%) | 46.0 | 42.7 | 11.3 | 58.6 | 34.2 | 6.2 | 66.0 | 29.8 | 4.2 | 35.6 | 44.8 | 19.6 | 30.5 | 47.0 | 22.5 | 28.4 | 50.2 | 21.4 | ||||||
| Sex (%) 1 | ||||||||||||||||||||||||
| male | 43.4 | 44.3 | 12.3 | 0.462 | 60.6 | 34.5 | 4.9 | 0.376 | 67.8 | 27.7 | 4.6 | 0.548 | 35.0 | 45.2 | 19.8 | 0.961 | 31.6 | 45.1 | 23.4 | 0.682 | 20.1 | 52.9 | 27.0 | 0.455 |
| female | 47.8 | 41.6 | 10.6 | 57.2 | 35.6 | 7.21 | 64.7 | 31.3 | 3.9 | 36.0 | 44.5 | 19.5 | 29.7 | 48.4 | 21.9 | 22.4 | 48.2 | 29.4 | ||||||
| Age group (%) 1 | ||||||||||||||||||||||||
| 12–19 years | 42.0 | 44.0 | 14.0 | 0.662 | 61.9 | 33.6 | 4.5 | 0.593 | 69.5 | 26.6 | 3.9 | 0.139 | 33.3 | 44.4 | 22.2 | 0.193 | 28.5 | 45.7 | 25.8 | 0.763 | 24.4 | 48.1 | 27.6 | 0.801 |
| 20–59 years | 48.2 | 41.2 | 10.6 | 56.0 | 36.3 | 7.7 | 69.5 | 26.2 | 4.4 | 36.9 | 47.8 | 15.4 | 32.1 | 45.7 | 22.2 | 19.4 | 52.0 | 28.6 | ||||||
| 60+ years | 45.8 | 43.7 | 10.6 | 59.6 | 34.8 | 5.7 | 60.5 | 35.3 | 4.2 | 35.5 | 41.8 | 22.7 | 29.9 | 48.9 | 21.1 | 21.9 | 49.5 | 28.6 | ||||||
| Race (%) 1 | ||||||||||||||||||||||||
| White | 42.4 | 44.4 | 13.2 | 0.035 * | 54.9 | 37.5 | 7.6 | 0.056 | 68.4 | 28.9 | 2.8 | 0.183 | 28.7 | 46.2 | 25.2 | 0.000 * | 32.4 | 47.3 | 20.3 | 0.003 * | 30.3 | 51.3 | 18.5 | 0.120 |
| Black | 59.7 | 38.7 | 1.6 | 69.4 | 27.4 | 3.2 | 65.6 | 29.5 | 4.9 | 54.8 | 38.7 | 6.5 | 29.5 | 57.4 | 13.1 | 16.1 | 51.6 | 32.3 | ||||||
| Mixed (white/black) | 49.3 | 41.0 | 9.7 | 63.7 | 32.3 | 4.0 | 61.2 | 31.7 | 7.1 | 43.1 | 43.5 | 13.5 | 28.4 | 44.4 | 27.1 | 28.1 | 47.8 | 24.1 | ||||||
| Asian/Indigenous | 42.9 | 35.7 | 21.4 | 41.7 | 41.7 | 16.7 | 71.4 | 28.6 | 0.0 | 42.9 | 50.0 | 7.1 | 7.7 | 30.8 | 61.5 | 28.6 | 50.0 | 21.4 | ||||||
| Hcy, µmol/L (median) 2 | 8.3 | 9.0 | 9.3 | 0.026 * | 8.7 | 8.9 | 7.5 | 0.180 | 8.6 | 8.9 | 8.7 | 0.569 | 9.0 | 8.2 | 9.4 | 0.097 | 8.9 | 8.8 | 8.5 | 0.401 | 8.4 | 8.9 | 8.8 | 0.355 |
| Folate, nmol/L (median) 2 | 27.9 | 27.9 | 24.1 | 0.088 | 26.5 | 28.5 | 22.8 | 0.565 | 28.0 | 25.1 | 21.0 | 0.015 * | 26.6 | 27.0 | 28.1 | 0.423 | 29.0 | 27.4 | 25.4 | 0.396 | 27.5 | 28.0 | 25.6 | 0.935 |
| Homocysteine 1 | Total | Folate Concentration | p-Interaction 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| First Tertile | Second Tertile | Third Tertile | |||||||
| Mean | SEM | Mean | SEM | Mean | SEM | Mean | SEM | ||
| MTHFR 677C>T | |||||||||
| C:C | 9.3 | 0.2 | 9.7 | 0.4 | 9.6 | 0.4 | 8.9 | 0.3 | 0.208 |
| C:T | 10.3 | 0.4 | 10.5 | 0.8 | 10.0 | 0.4 | 10.5 | 1.0 | |
| T:T | 12.6 | 1.3 | 12.6 | 1.3 | 10.2 | 1.7 | 13.9 | 3.9 | |
| p-value 2 | 0.000 * | 0.006 * | 0.162 | 0.038 * | |||||
| MTHFR 1298A>C | |||||||||
| A:A | 10.5 | 0.4 | 11.0 | 0.6 | 9.5 | 0.4 | 10.9 | 1.0 | 0.327 |
| A:C | 9.7 | 0.2 | 10.3 | 0.5 | 10.0 | 0.5 | 9.0 | 0.3 | |
| C:C | 9.2 | 0.7 | 7.1 | 0.6 | 12.1 | 1.6 | 8.4 | 1.3 | |
| p-value 2 | 0.304 | 0.121 | 0.190 | 0.187 | |||||
| MTR 2756A>G | |||||||||
| A:A | 10.0 | 0.3 | 10.1 | 0.4 | 9.5 | 0.3 | 10.4 | 0.8 | 0.397 |
| A:G | 10.4 | 0.5 | 11.5 | 1.2 | 10.5 | 0.7 | 9.2 | 0.4 | |
| G:G | 9.1 | 0.5 | 9.6 | 0.7 | 8.5 | 1.0 | 9.8 | 1.5 | |
| p-value 2 | 0.439 | 0.583 | 0.859 | 0.374 | |||||
| MTRR 66A>G | |||||||||
| A:A | 10.2 | 0.4 | 10.4 | 0.5 | 10.0 | 0.6 | 10.6 | 1.1 | 0.825 |
| A:G | 10.0 | 0.4 | 10.6 | 0.8 | 9.5 | 0.5 | 9.6 | 0.9 | |
| G:G | 10.1 | 0.4 | 9.8 | 0.9 | 10.1 | 0.6 | 10.4 | 0.7 | |
| p-value 2 | 0.576 | 0.59 | 0.612 | 0.388 | |||||
| RFC1 80G>A | |||||||||
| G:G | 10.7 | 0.5 | 11.7 | 1.2 | 11.0 | 0.8 | 9.2 | 0.5 | 0.214 |
| G:A | 10.1 | 0.4 | 9.6 | 0.4 | 9.7 | 0.4 | 11.2 | 1.2 | |
| A:A | 9.4 | 0.3 | 10.5 | 0.7 | 8.5 | 0.5 | 9.2 | 0.5 | |
| p-value 2 | 0.281 | 0.920 | 0.001 * | 0.396 | |||||
| deletion DHFR | |||||||||
| WT:WT | 10.4 | 0.6 | 10.5 | 1.3 | 9.7 | 0.8 | 10.8 | 1.3 | 0.535 |
| WT:del | 10.1 | 0.3 | 10.2 | 0.4 | 9.8 | 0.4 | 10.1 | 0.8 | |
| del:del | 10.1 | 0.4 | 11.2 | 0.8 | 10.3 | 0.6 | 8.9 | 0.5 | |
| p-value 2 | 0.312 | 0.071 | 0.667 | 0.836 | |||||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steluti, J.; Carvalho, A.M.; Carioca, A.A.F.; Miranda, A.; Gattás, G.J.F.; Fisberg, R.M.; Marchioni, D.M. Genetic Variants Involved in One-Carbon Metabolism: Polymorphism Frequencies and Differences in Homocysteine Concentrations in the Folic Acid Fortification Era. Nutrients 2017, 9, 539. https://doi.org/10.3390/nu9060539
Steluti J, Carvalho AM, Carioca AAF, Miranda A, Gattás GJF, Fisberg RM, Marchioni DM. Genetic Variants Involved in One-Carbon Metabolism: Polymorphism Frequencies and Differences in Homocysteine Concentrations in the Folic Acid Fortification Era. Nutrients. 2017; 9(6):539. https://doi.org/10.3390/nu9060539
Chicago/Turabian StyleSteluti, Josiane, Aline M. Carvalho, Antonio A. F. Carioca, Andreia Miranda, Gilka J. F. Gattás, Regina M. Fisberg, and Dirce M. Marchioni. 2017. "Genetic Variants Involved in One-Carbon Metabolism: Polymorphism Frequencies and Differences in Homocysteine Concentrations in the Folic Acid Fortification Era" Nutrients 9, no. 6: 539. https://doi.org/10.3390/nu9060539
APA StyleSteluti, J., Carvalho, A. M., Carioca, A. A. F., Miranda, A., Gattás, G. J. F., Fisberg, R. M., & Marchioni, D. M. (2017). Genetic Variants Involved in One-Carbon Metabolism: Polymorphism Frequencies and Differences in Homocysteine Concentrations in the Folic Acid Fortification Era. Nutrients, 9(6), 539. https://doi.org/10.3390/nu9060539