Brain Functional Connectivity Is Modified by a Hypocaloric Mediterranean Diet and Physical Activity in Obese Women


 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Intervention
2.3. Study Design
2.4. Image Acquisition
2.5. Image Processing
2.6. Statistical Analysis
3. Results
3.1. Demographic and Clinical Characteristics
3.2. Neuroimaging Connectivity
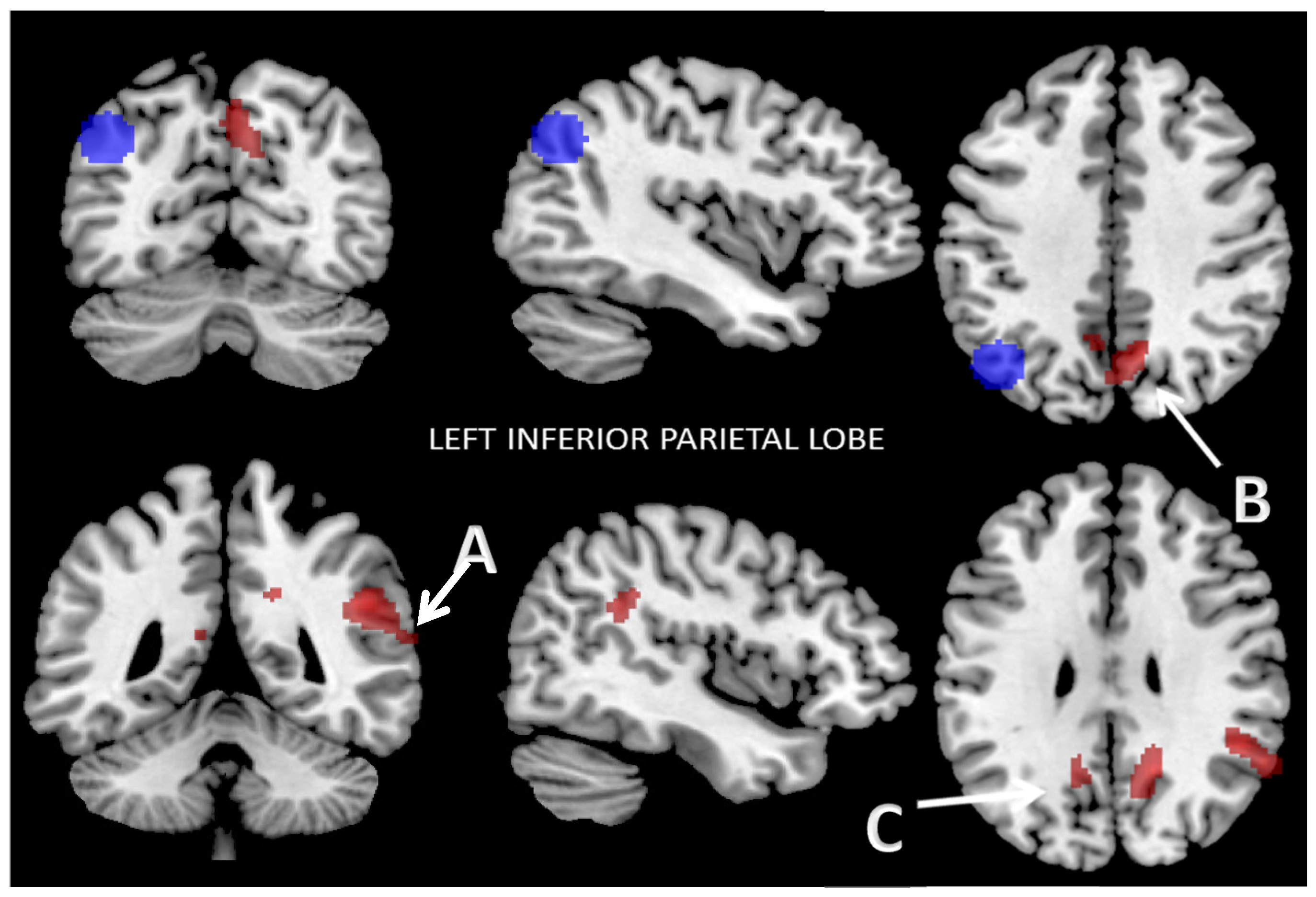
3.2.1. Functional Connectivity of the Left Inferior Parietal Lobe
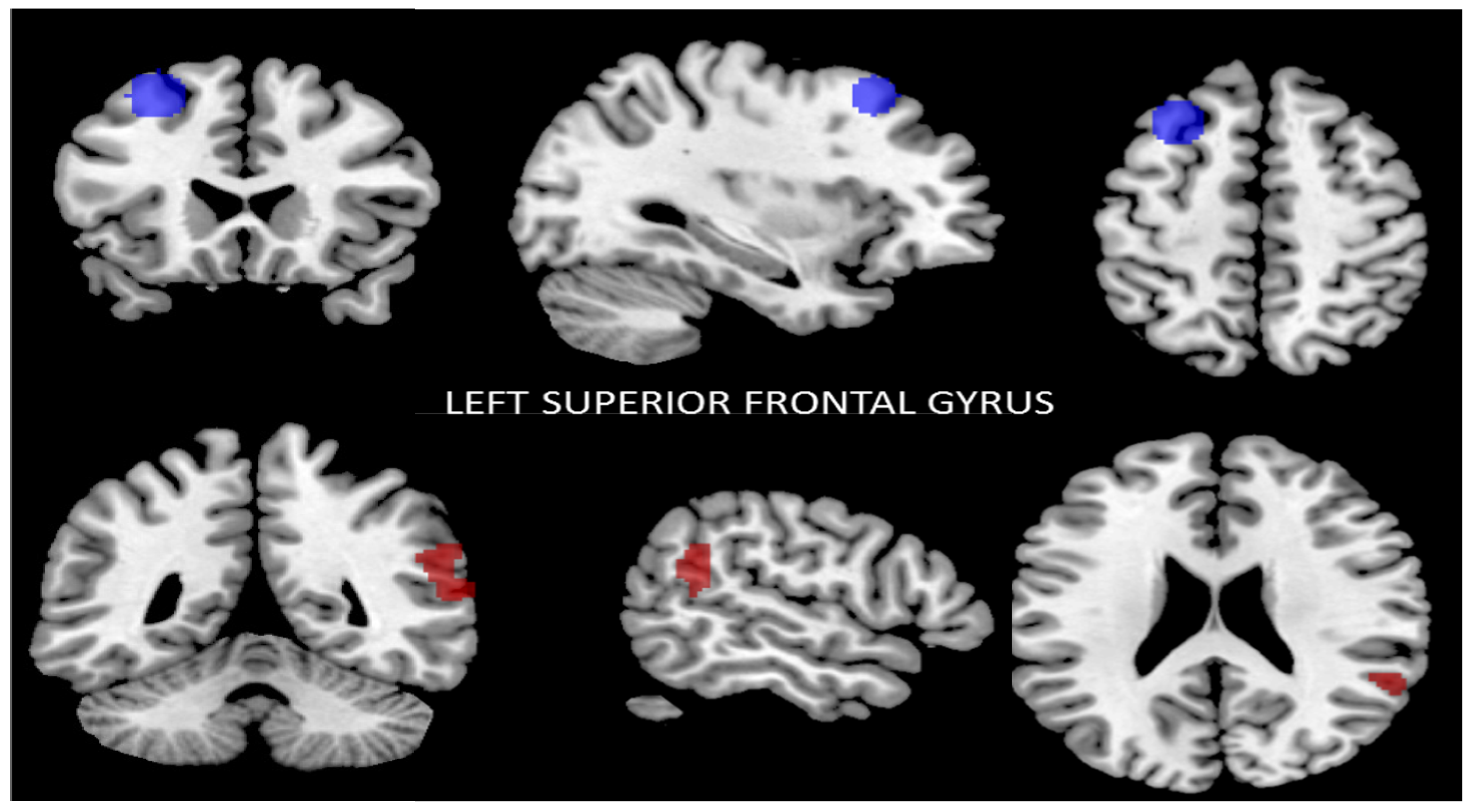
3.2.2. Functional Connectivity of the Left Superior Frontal Gyrus
3.2.3. Functional Connectivity of the Right Posterior Superior Temporal Gyrus
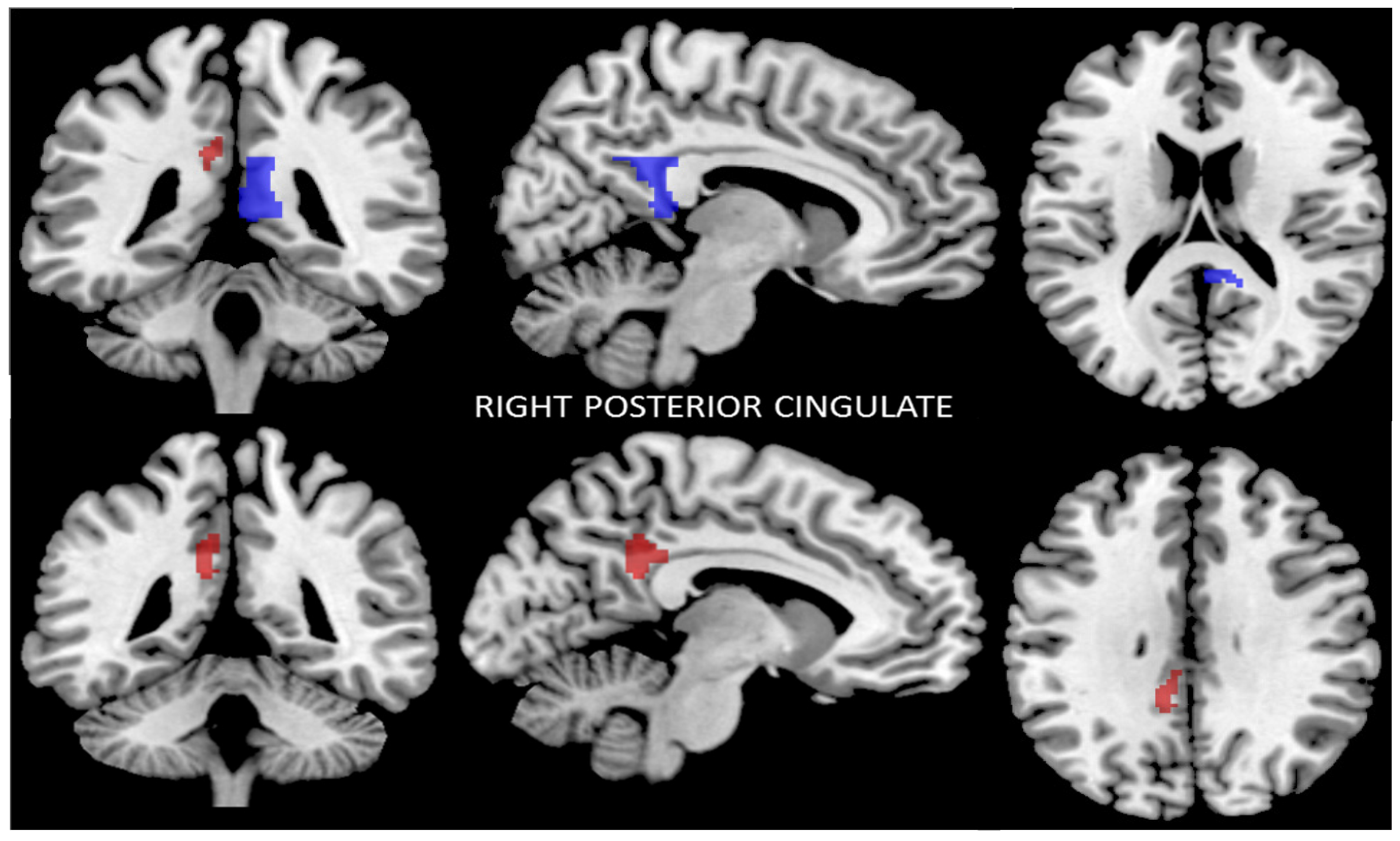
3.2.4. Functional Connectivity of the Right Posterior Cingulate Cortex
4. Discussion
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carnell, S.; Gibson, C.; Benson, L.; Ochner, C.N.; Geliebter, A. Neuroimaging and obesity: Current knowledge and future directions. Obes. Rev. 2012, 13, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Burger, K.S.; Berner, L.A. A functional neuroimaging review of obesity, appetitive hormones and ingestive behavior. Physiol. Behav. 2014, 136, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.T.; Davis, C.R.; Dyer, K.A.; Hodgson, J.M.; Woodman, R.J.; Keage, H.A.; Murphy, K.J. A Mediterranean Diet to Improve Cardiovascular and Cognitive Health: Protocol for a Randomised Controlled Intervention Study. Nutrients 2017, 9, E145. [Google Scholar] [CrossRef] [PubMed]
- Buckland, G.; Bach, A.; Serra-Majem, L. Obesity and the Mediterranean diet: A systematic review of observational and intervention studies. Obes. Rev. 2008, 9, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Kastorini, C.M.; Panagiotakos, D.B.; Giugliano, D. Mediterranean diet and weight loss: Meta-analysis of randomized controlled trials. Metab. Syndr. Relat. Disord. 2011, 9, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Luciano, M.; Corley, J.; Cox, S.R.; Valdés Hernández, M.C.; Craig, L.C.; Dickie, D.A.; Karama, S.; McNeill, G.M.; Bastin, M.E.; Wardlaw, J.M.; et al. Mediterranean-type diet and brain structural change from 73 to 76 years in a Scottish cohort. Neurology 2017, 88, 449–455. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Brickman, A.M.; Stern, Y.; Habeck, C.G.; Razlighi, Q.R.; Luchsinger, J.A.; Manly, J.J.; Schupf, N.; Mayeux, R.; Scarmeas, N. Mediterranean diet and brain structure in a multiethnic elderly cohort. Neurology 2015, 85, 1744–1751. [Google Scholar] [CrossRef] [PubMed]
- Titova, O.E.; Ax, E.; Brooks, S.J.; Sjögren, P.; Cederholm, T.; Kilander, L.; Kullberg, J.; Larsson, E.M.; Johansson, L.; Ahlström, H.; et al. Mediterranean diet habits in older individuals: Associations with cognitive functioning and brain volumes. Exp. Gerontol. 2013, 48, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Staubo, S.C.; Aakre, J.A.; Vemuri, P.; Syrjanen, J.A.; Mielke, M.M.; Geda, Y.E.; Kremers, W.K.; Machulda, M.M.; Knopman, D.S.; Petersen, R.C.; et al. Mediterranean diet, micronutrients and macronutrients, and MRI measures of cortical thickness. Alzheimer Dement. 2017, 13, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Mosconi, L.; Murray, J.; Tsui, W.H.; Li, Y.; Davies, M.; Williams, S.; Pirraglia, E.; Spector, N.; Osorio, R.S.; Glodzik, L.; et al. Mediterranean diet and magnetic resonance imaging-assessed brain atrophy in cognitively normal individuals at risk for Alzheimer’s disease. J. Prev. Alzheimer Dis. 2014, 1, 23–32. [Google Scholar]
- Pelletier, A.; Barul, C.; Féart, C.; Helmer, C.; Bernard, C.; Periot, O.; Dilharreguy, B.; Dartigues, J.F.; Allard, M.; Barberger-Gateau, P.; et al. Mediterranean diet and preserved brain structural connectivity in older subjects. Alzheimer Dement. 2015, 11, 1023–1031. [Google Scholar] [CrossRef] [PubMed]
- Honea, R.A.; Szabo-Reed, A.N.; Lepping, R.J.; Perea, R.; Breslin, F.; Martin, L.E.; Brooks, W.M.; Donnelly, J.E.; Savage, C.R. Voxel-based morphometry reveals brain gray matter volume changes in successful dieters. Obesity 2016, 24, 1842–1848. [Google Scholar] [CrossRef] [PubMed]
- Olivo, G.; Wiemerslage, L.; Nilsson, E.K.; Solstrand Dahlberg, L.; Larsen, A.L.; Olaya Búcaro, M.; Gustafsson, V.P.; Titova, O.E.; Bandstein, M.; Larsson, E.M.; et al. Resting-state brain and the FTO obesity risk allele: Default mode, sensorimotor, and salience network connectivity underlying different somatosensory integration and reward processing between genotypes. Front. Hum. Neurosci. 2016, 10. [Google Scholar] [CrossRef] [PubMed]
- Tregellas, J.R.; Wylie, K.P.; Rojas, D.C.; Tanabe, J.; Martin, J.; Kronberg, E.; Cordes, D.; Cornier, M. Altered Default Network Activity in Obesity. Obesity (Silver Spring) 2011, 19, 2316–2321. [Google Scholar] [CrossRef] [PubMed]
- García-García, I.; Jurado, M.A.; Garolera, M.; Segura, B.; Sala-Llonch, R.; Marqués-Iturria, I.; Pueyo, R.; Sender-Palacios, M.J.; Vernet-Vernet, M.; Narberhaus, A.; et al. Alterations of the salience network in obesity: A resting-state fMRI study. Hum. Brain Mapp. 2013, 34, 2786–2797. [Google Scholar] [CrossRef] [PubMed]
- McFadden, K.L.; Cornier, M.A.; Melanson, E.L.; Bechtell, J.L.; Tregellas, J.R. Effects of exercise on resting-state default mode and salience network activity in overweight/obese adults. Neuroreport 2013, 24, 866–871. [Google Scholar] [CrossRef] [PubMed]
- Lepping, R.J.; Bruce, A.S.; Francisco, A.; Yeh, H.W.; Martin, L.E.; Powell, J.N.; Hancock, L.; Patrician, T.M.; Breslin, F.J.; Selim, N.; et al. Resting-state brain connectivity after surgical and behavioral weight loss. Obesity (Silver Spring) 2015, 23, 1422–1428. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Andrews-Hanna, J.R.; Schacter, D.L. The brain’s default network: Anatomy, function, and relevance to disease. Ann. N. Y. Acad. Sci. 2008, 1124, 1–38. [Google Scholar] [CrossRef] [PubMed]
- Seeley, W.W.; Menon, V.; Schatzberg, A.F.; Keller, J.; Glover, G.H.; Kenna, H.; Reiss, A.L.; Greicius, M.D. Dissociable intrinsic connectivity networks for salience process in gand executive control. J. Neurosci. 2007, 27, 2349–2356. [Google Scholar] [CrossRef] [PubMed]
- Steward, T.; Picó-Pérez, M.; Mata, F.; Martínez-Zalacaín, I.; Cano, M.; Contreras-Rodríguez, O.; Fernández-Aranda, F.; Yucel, M.; Soriano-Mas, C.; Verdejo-García, A. Emotion Regulation and Excess Weight: Impaired Affective Processing Characterized by Dysfunctional Insula Activation and Connectivity. PLoS ONE 2016, 11, e0152150. [Google Scholar] [CrossRef] [PubMed]
- Kullmann, S.; Heni, M.; Veit, R.; Ketterer, C.; Schick, F.; Haring, H.U.; Fritsche, A.; Preissl, H. The obese brain: Association of body mass index and insulin sensitivity with resting state network functional connectivity. Hum. Brain Mapp. 2012, 33, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Coveleskie, K.; Gupta, A.; Kilpatrick, L.A.; Mayer, E.D.; Ashe-McNalley, C.; Stains, J.; Labus, J.S.; Mayer, E.A. Altered functional connectivity within the central reward network in overweight and obese women. Nutr. Diabetes 2015, 5, e148. [Google Scholar] [CrossRef] [PubMed]
- Evero, N.; Hackett, L.C.; Clark, R.D.; Phelan, S.; Hagobian, T.A. Aerobic exercise reduces neuronal responses in food reward brain regions. J. Appl. Physiol. 2012, 112, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.M.; Fox, P.T.; Miller, K.L.; Glahn, D.C.; Fox, P.M.; Mackay, C.E.; Filippini, N.; Watkins, K.E.; Toro, R.; Laird, A.R.; et al. Correspondence of the brain’s functional architecture during activation and rest. Proc. Natl. Acad. Sci. USA 2009, 106, 13040–13045. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.J.; Volkow, N.D.; Felder, C.; Fowler, J.S.; Levy, A.V.; Pappas, N.R.; Wong, C.T.; Zhu, W.; Netusil, N. Enhanced resting activity of the oral somatosensory cortex in obese subjects. NeuroReport 2002, 13, 1151–1155. [Google Scholar] [CrossRef] [PubMed]
- Babio, N.; Toledo, E.; Estruch, R.; Ros, E.; Martínez-González, M.A.; Castañer, O.; Bulló, M.; Corella, D.; Arós, F.; Gómez-Gracia, E.; et al. Mediterranean diets and metabolic syndrome status in the PREDIMED randomized trial. CMAJ 2014, 186, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Mente, A.; de Koning, L.; Shannon, H.S.; Anand, S.S. A systematic review of the evidence supporting a causal link between dietary factors and coronary heart disease. Arch. Intern. Med. 2009, 169, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Ballart, J.D.; Pinol, J.L.; Zazpe, I.; Corella, D.; Carrasco, P.; Toledo, E.; Perez-Bauer, M.; Martínez-González, M.A.; Salas-Salvadó, J.; Martín-Moreno, J.M. Relative validity of a semi-quantitative food-frequency questionnaire in an elderly Mediterranean population of Spain. Br. J. Nutr. 2009, 103, 1808–1816. [Google Scholar] [CrossRef] [PubMed]
- De la Fuente-Arrillaga, C.; Vazquez, Z.; Bes-Rastrollo, M.; Sampson, L.; Martınez-Gonzalez, M.A. Reproducibility of a Food Frequency Questionnaire (FFQ) Validated in Spain. Public Health Nutr. 2009, 13, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, M.A.; Garcia-Arellano, A.; Toledo, E.; Salas-Salvadó, J.; Buil-Cosiales, P.; Corella, D.; Covas, M.I.; Schröder, H.; Arós, F.; Gómez-Gracia, E.; et al. A 14-item Mediterranean diet assessment tool and obesity indexes among high-risk subjects: The PREDIMED trial. PLoS ONE 2012, 7, e43134. [Google Scholar] [CrossRef] [PubMed]
- The Diabetes Prevention Program Research Group. Reduction in the incidence of type 2 diabetes with lifestyle intervention or metformin. N. Engl. J. Med. 2002, 346, 393–403. [Google Scholar]
- Maldjian, J.A.; Laurienti, P.J.; Burdette, J.B.; Kraft, R.A. An Automated Method for Neuroanatomic and Cytoarchitectonic Atlas-based Interrogation of fMRI Data Sets. NeuroImage 2003, 19, 1233–1239. [Google Scholar] [CrossRef]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Greicius, M.D.; Krasnow, B.; Reiss, A.L.; Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. Proc. Natl. Acad. Sci. USA 2003, 100, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Fransson, P.; Marrelec, G. The precuneus/posterior cingulate cortex plays a pivotal role in the default mode network: Evidence from a partial correlation network analysis. Neuroimage 2008, 42, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Tomasi, D.; Baler, R. Food and drug reward: Overlapping circuits in human obesity and addiction. Curr. Top. Behav. Neurosci. 2012, 11, 1–24. [Google Scholar] [PubMed]
- Menon, V.; Uddin, L.Q. Saliency, switching, attention and control: A network model of insula function. Brain Struct. Funct. 2010, 214, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Stoeckel, L.E.; Kim, J.; Weller, R.E.; Cox, J.E.; Cook, E.W., 3rd; Horwitz, B. Effective connectivity of a reward network in obese woman. Brain Res. Bull. 2009, 79, 388–395. [Google Scholar] [CrossRef] [PubMed]
- Lips, M.A.; Wijngaarden, M.A.; van der Grond, J.; van Buchem, M.A.; de Groot, G.H.; Rombouts, S.A.; Pijl, H.; Veer, I.M. Resting-state functional connectivity of brain regions involved in cognitive control, motivation, and reward is enhanced in obese females. Am. J. Clin. Nutr. 2014, 100, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Tataranni, P.A.; Gautier, J.F.; Chen, K.; Uecker, A.; Bandy, D.; Salbe, A.D.; Pratley, R.E.; Lawson, M.; Reiman, E.M.; Ravussin, E. Neuroanatomical correlates of hunger and satiation in humans using positron emission tomography. Proc. Natl. Acad. Sci. USA 1999, 96, 4569–4574. [Google Scholar] [CrossRef] [PubMed]
- Del Parigi, A.; Gautier, J.F.; Chen, K.; Salbe, A.D.; Ravussin, E.; Reiman, E.; Tataranni, P.A. Neuroimaging and obesity: Mapping the brain responses to hunger and satiation in humans using positron emission tomography. Ann. N. Y. Acad. Sci. USA 2002, 967, 389–397. [Google Scholar] [CrossRef]
- Wijngaarden, M.A.; Veer, I.M.; Rombouts, S.A.; van Buchem, M.A.; Willems van Dijk, K.; Pijl, H.; van der Grond, J. Obesity is marked by distinct functional connectivity in brain networks involved in food reward and salience. Behav. Brain Res. 2015, 287, 127–134. [Google Scholar] [CrossRef] [PubMed]
- McFadden, K.L.; Tregellas, J.R.; Shott, M.E.; Frank, G.K. Reduced salience and default mode network activity in women with anorexia nervosa. J. Psychiatry Neurosci. 2014, 39, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.; Mayhew, S.D.; Chechlacz, M.; Bagary, M.; Bagshaw, A.P. The structural and functional connectivity of the posterior cingulate cortex: Comparison between deterministic and probabilistic tractography for the investigation of structure-function relationships. Neuroimage 2014, 102, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Legget, K.T.; Wylie, K.P.; Cornier, M.A.; Melanson, E.L.; Paschall, C.J.; Tregellas, J.R. Exercise-related changes in between-network connectivity in overweight/obese adults. Physiol. Behav. 2016, 158, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Hillman, CH.; Erickson, K.I.; Kramer, A.F. Be smart, exercise your heart: Exercise effects on brain and cognition. Nat. Rev. Neurosci. 2008, 9, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef] [PubMed]
- Vaynman, S.; Gomez-Pinilla, F. License to run: Exercise impacts functional plasticity in the intact and injured central nervous system by using neurotrophins. Neurorehabiliation Neural Repair 2005, 19, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Jung, T.W.; Lee, J.Y.; Shim, W.S.; Kang, E.S.; Kim, J.S.; Ahn, C.W.; Lee, H.C.; Cha, B.S. Adiponectin protects human neuroblastoma SH-SY5Y cells against acetaldehyde-induced cytotoxicity. Biochem. Pharmacol. 2006, 72, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Qiu, G.; Wan, R.; Hu, J.; Mattson, M.P.; Spangler, E.; Liu, S.; Yau, S.Y.; Lee, T.M.; Gleichmann, M.; Ingram, D.K.; et al. Adiponectin protects rat hippocampal neurons against excitotoxicity. Age 2011, 33, 155–165. [Google Scholar] [CrossRef] [PubMed]
- García-Casares, N.; García-Arnés, J.A.; Rioja, J.; Ariza, M.J.; Gutiérrez, A.; Alfaro, F.; Nabrozidis, A.; González-Alegre, P.; González-Santos, P. Alzheimer’s like brain changes correlate with low adiponectin plasma levels in type 2 diabetic patients. J. Diabetes Complicat. 2016, 30, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Tian, D.; Yu, C.; Zhang, J.; Tian, X.; von Deneen, K.M.; Zang, Y.; Walter, M.; Liu, Y. Altered baseline brain activities before food intake in obese men: A resting state fMRI study. Neurosci. Lett. 2015, 584, 156–161. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, J.; Dimitropoulos, A. Influence of feeding state on neurofunctional differences between individuals who are obese and normal weight: A meta-analysis of neuroimaging studies. Appetite 2014, 75, 103–109. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Before the Mediterranean Diet/Physical Activity Program | After the Mediterranean Diet/Physical Activity Program | |
|---|---|---|
| N (women) | 16 | |
| Age (years) | 46.3 ± 4.07 | |
| Weight (kg) | 98.5 ± 13.1 | 88.2 ±12.2 * |
| Body mass index (kg/m2) | 38.1 ± 4.7 | 34.1 ± 4.5 * |
| Waist circumference(cm) | 115.3 ± 11.8 | 110.5 ± 10.9 |
| Glucose (mg/dL) | 92.9 ± 12.5 | 82.5 ± 12.9 * |
| HbA1c (%) | 5.5 ± 0.4 | 5.4 ± 0.3 |
| Creatinine (mg/dL) | 0.6 ± 0.1 | 0.6 ± 0.1 |
| Uric acid (mg/dL) | 13.6 ± 34.0 | 5.0 ± 0.7 |
| Total cholesterol (mg/dL) | 202.1 ± 38.2 | 182.8 ± 34.7 * |
| LDL cholesterol (mg/dL) | 124.1 ± 33.4 | 109.4 ± 33.6 * |
| HDL cholesterol (mg/dL) | 56.0 ± 9.7 | 50.8 ± 10.7 |
| Triglycerides (mg/dL) | 93.50 (73.75–122.50) | 94.50 (74.00–150.5) |
| Insulin (µIU/mL) | 15.4 ± 5.5 | 11.5 ± 5.1 * |
| HOMA-IR index | 3.5 ± 1.3 | 2.3 ± 0.9 * |
| Weight Loss | Baseline | 6 Months | p | |
|---|---|---|---|---|
| Energy (kcal) | <5% | 2537.9 ± 100.3 | 1909.1 ± 70.9 | <0.0001 |
| ≥5–<10% | 2440.8 ± 157.1 | 1658.2 ± 90.8 | 0.001 | |
| ≥10% | 2339.8 ± 79.0 | 1621.0 ± 48.6 | <0.0001 | |
| Energy from total carbohydrate (%) | <5% | 250.1 ± 12.8 | 181.2 ± 7.4 | <0.0001 |
| ≥5–<10% | 246.1 ± 16.1 | 169.7 ± 14.6 | 0.005 | |
| ≥10% | 230.3 ± 10.5 | 172.9 ± 5.8 | <0.0001 | |
| Energy from total protein (%) | <5% | 95.9 ± 3.6 | 83.6 ± 3.6 | 0.01 |
| ≥5–<10% | 92.3 ± 3.9 | 76.4 ± 3.3 | 0.001 | |
| ≥10% | 89.4 ± 3.1 | 72.6 ± 2.1 | 0.001 | |
| Energy from total fat (%) | <5% | 122.0 ± 5.2 | 92.9 ± 4.5 | <0.0001 |
| ≥5–<10% | 124.6 ± 7.7 | 72.6 ± 4.4 | 0.001 | |
| ≥10% | 115.6 ± 4.9 | 70.4 ± 4.1 | <0.0001 | |
| SFA | <5% | 36.7 ± 2.3 | 25.6 ± 1.7 | <0.0001 |
| ≥5–<10% | 38.2 ± 3.1 | 18.9 ± 1.3 | <0.0001 | |
| ≥10% | 44.6 ± 10.1 | 17.9 ± 1.5 | <0.0001 | |
| MUFA | <5% | 59.3 ± 2.3 | 62.0 ± 14.1 | <0.0001 |
| ≥5–<10% | 59.4 ± 3.3 | 38.2 ± 2.6 | 0.001 | |
| ≥10% | 55.4 ± 2.4 | 37.4 ± 2.2 | <0.0001 | |
| PUFA | <5% | 17.2 ± 1.5 | 11.3 ± 0.6 | <0.0001 |
| ≥5–<10% | 18.6 ± 2.1 | 9.7 ± 0.9 | <0.0001 | |
| ≥10% | 16.5 ± 1.3 | 10.1 ± 0.8 | 0.001 | |
| Cholesterol (mg/d) | <5% | 382.5 ± 24.7 | 334.0 ± 25.6 | 0.3 |
| ≥5–<10% | 417.6 ± 26.5 | 238.9 ± 22.6 | 0.001 | |
| ≥10% | 395.7 ± 22.2 | 232.1 ± 12.9 | <0.0001 | |
| Fiber (g/d) | <5% | 21.9 ± 1.5 | 18.8 ± 1.2 | 0.3 |
| ≥5–<10% | 18.9 ± 1.2 | 20.2 ± 1.3 | 0.6 | |
| ≥10% | 17.2 ± 1.0 | 23.5 ± 1.4 | 0.001 | |
| Vitamin D (µg/d) | <5% | 3.7 ± 0.5 | 6.3 ± 1.2 | 0.3 |
| ≥5–<10% | 2.3 ± 0.4 | 6.1 ± 1.2 | 0.01 | |
| ≥10% | 3.7 ± 1.3 | 9.2 ± 3.3 | 0.004 |
| Seed | Left Inferior Parietal Lobe Functional Connectivity MNI (x, y, z): (−42, −68, 38) | |||
| Cluster 1 | Brodmann Area | MNI (x, y, z) | Cluster Size (K) | p-value (FWE) |
| R AG | 39 | +50, −46, +28 | 136 | <0.001 |
| R STG | 22 | 110 | ||
| R SmG | 40 | 85 | ||
| R IC | 13 | 25 | ||
| Cluster 2 | Brodmann area | MNI (x, y, z) | Cluster size (K) | p-value (FWE) |
| R DPCC | 31 | +16, −56, +30 | 170 | <0.001 |
| R SsAC | 7 | 169 | ||
| L SsAC | 7 | 100 | ||
| Cluster 3 | Brodmann area | MNI (x, y, z) | Cluster size (K) | p-value (FWE) |
| L DPCC | 31 | –08, −60, +36 | 63 | <0.06 <0.03 (FDR) |
| L SsAC | 7 | 30 | ||
| L RsCC | 29 | 15 | ||
| L CC | 30 | 5 | ||
| Seed | Left Superior Frontal Gyrus Functional Connectivity MNI: (−28, 22, 52) | |||
| Brodmann area | MNI (x, y, z) | Cluster size (K) | p-value (FWE) | |
| R SmG | 40 | +64, −50, +12 | 68 | <0.01 |
| R STG | 22 | 61 | ||
| R IC | 13 | 18 | ||
| R AG | 39 | 14 | ||
| Seed | Right Posterior Superior Temporal Gyrus (pSTG) Functional Connectivity MNI (x, y, z): (60, −30, 24) | |||
| Brodmann area | MNI (x, y, z) | Cluster size (K) | p-value (FWE) | |
| L PmC | 6 | −36, −10, +68 | 137 | <0.025 |
| L PMC | 4 | 29 | ||
| L PSsC | 3 | 9 | ||
| Seed | Right Posterior Cingulate Cortex Functional Connectivity MNI (x, y, z): (0, 6, 40) | |||
| Brodmann area | MNI (x, y, z) | Cluster size (K) | p-value (FWE) | |
| L dpCC | 31 | −10, −46, +24 | 120 | <0.04 |
| L vpCC | 23 | 2 | ||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Casares, N.; Bernal-López, M.R.; Roé-Vellvé, N.; Gutiérrez-Bedmar, M.; Fernández-García, J.C.; García-Arnés, J.A.; Ramos-Rodriguez, J.R.; Alfaro, F.; Santamaria-Fernández, S.; Steward, T.; et al. Brain Functional Connectivity Is Modified by a Hypocaloric Mediterranean Diet and Physical Activity in Obese Women. Nutrients 2017, 9, 685. https://doi.org/10.3390/nu9070685
García-Casares N, Bernal-López MR, Roé-Vellvé N, Gutiérrez-Bedmar M, Fernández-García JC, García-Arnés JA, Ramos-Rodriguez JR, Alfaro F, Santamaria-Fernández S, Steward T, et al. Brain Functional Connectivity Is Modified by a Hypocaloric Mediterranean Diet and Physical Activity in Obese Women. Nutrients. 2017; 9(7):685. https://doi.org/10.3390/nu9070685
Chicago/Turabian StyleGarcía-Casares, Natalia, María R. Bernal-López, Nuria Roé-Vellvé, Mario Gutiérrez-Bedmar, Jose C. Fernández-García, Juan A. García-Arnés, José R. Ramos-Rodriguez, Francisco Alfaro, Sonia Santamaria-Fernández, Trevor Steward, and et al. 2017. "Brain Functional Connectivity Is Modified by a Hypocaloric Mediterranean Diet and Physical Activity in Obese Women" Nutrients 9, no. 7: 685. https://doi.org/10.3390/nu9070685
APA StyleGarcía-Casares, N., Bernal-López, M. R., Roé-Vellvé, N., Gutiérrez-Bedmar, M., Fernández-García, J. C., García-Arnés, J. A., Ramos-Rodriguez, J. R., Alfaro, F., Santamaria-Fernández, S., Steward, T., Jiménez-Murcia, S., Garcia-Garcia, I., Valdivielso, P., Fernández-Aranda, F., Tinahones, F. J., & Gómez-Huelgas, R. (2017). Brain Functional Connectivity Is Modified by a Hypocaloric Mediterranean Diet and Physical Activity in Obese Women. Nutrients, 9(7), 685. https://doi.org/10.3390/nu9070685