Docosahexaenoic Acid Inhibits Cerulein-Induced Acute Pancreatitis in Rats



Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Measurement of Pancreatic Edema
2.4. Determination of LPO and MPO Activity
2.5. Real-Time Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
2.6. Western Blot Analysis
2.7. Enzyme-Linked Immunosorbent Assay (ELISA)
2.8. Electrophoretic Mobility Shift Assay (EMSA)
2.9. Immunohistochemical Analysis
2.10. Histological Observation
2.11. Statistical Analysis
3. Results
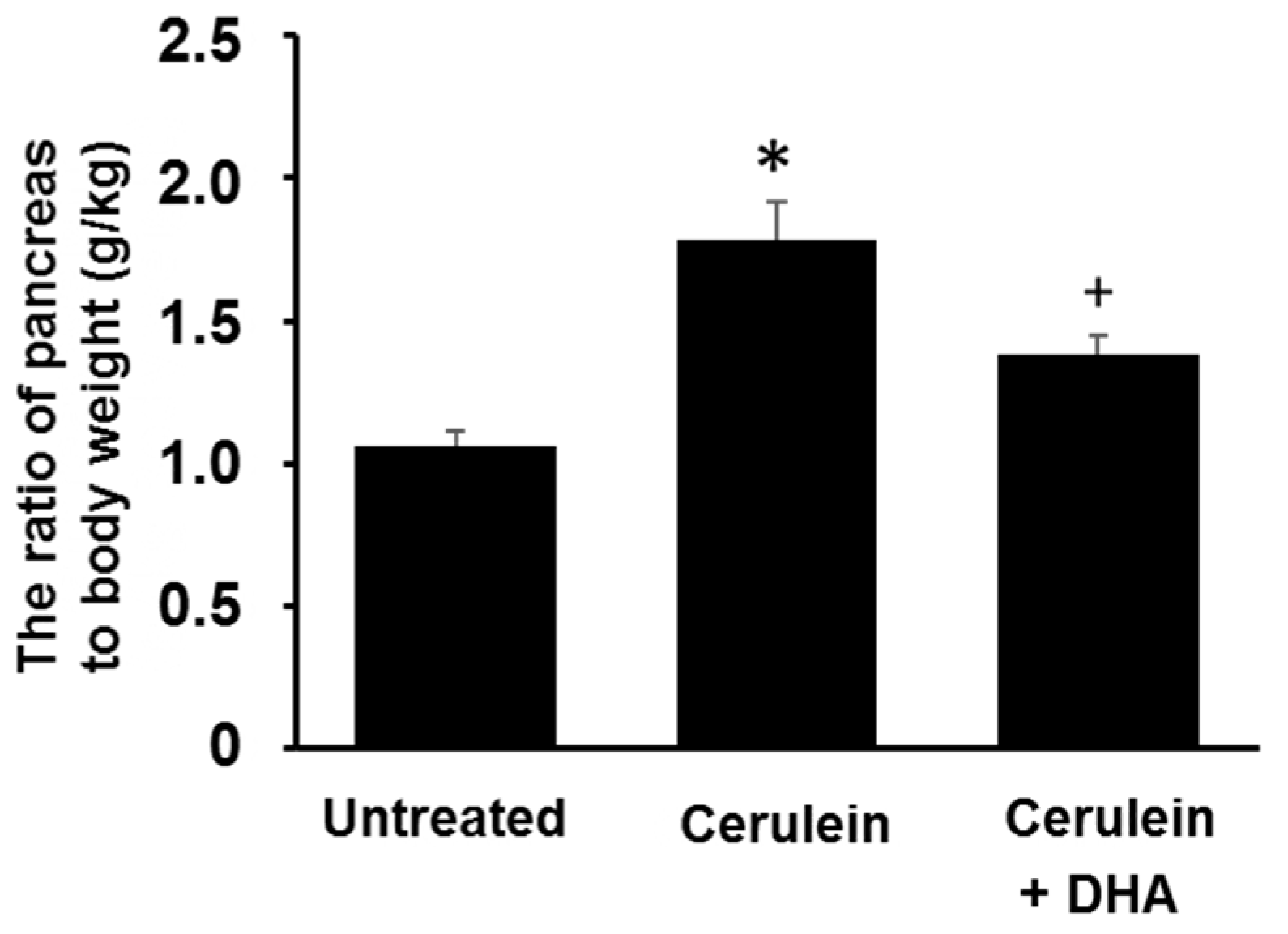
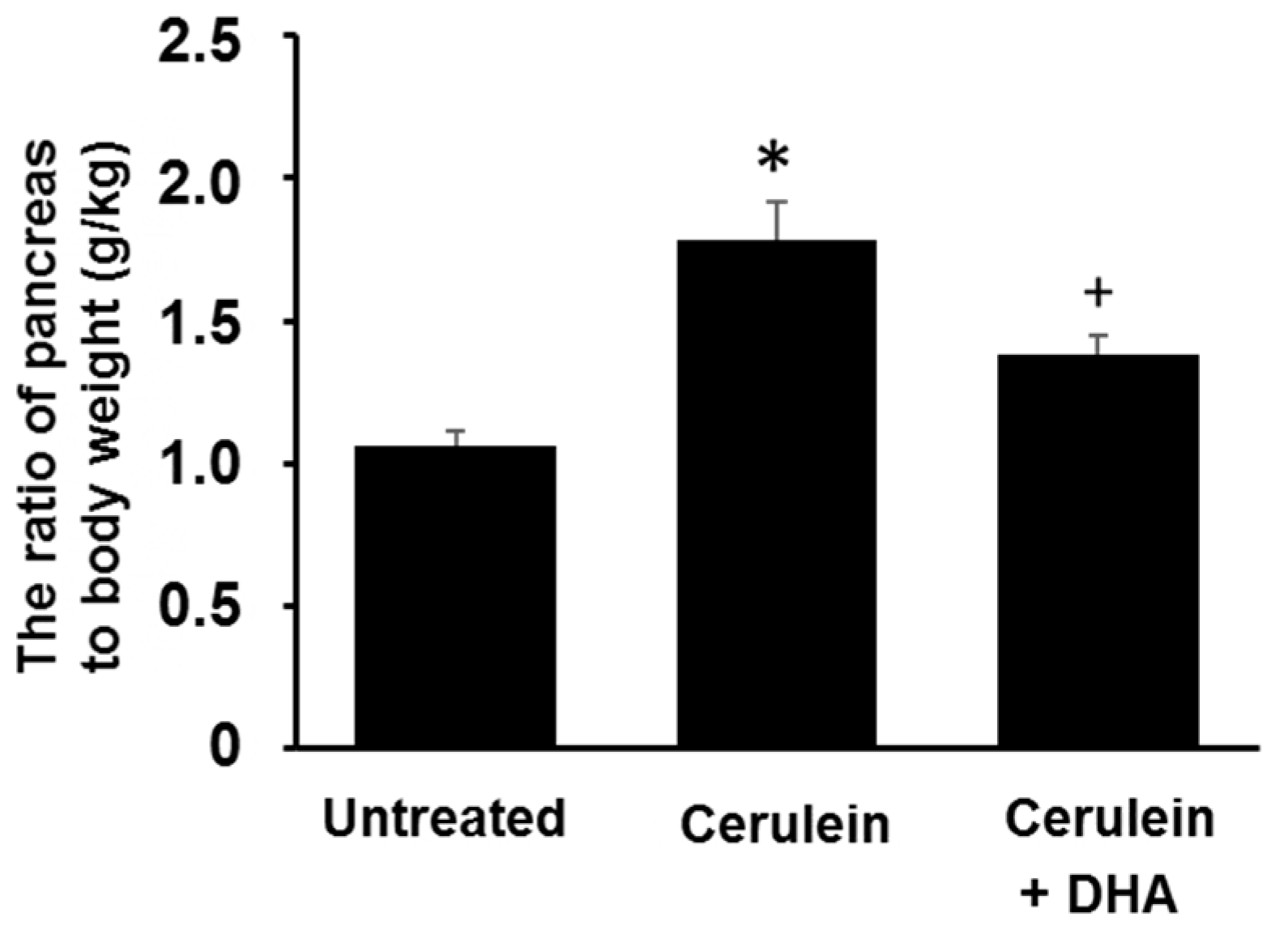
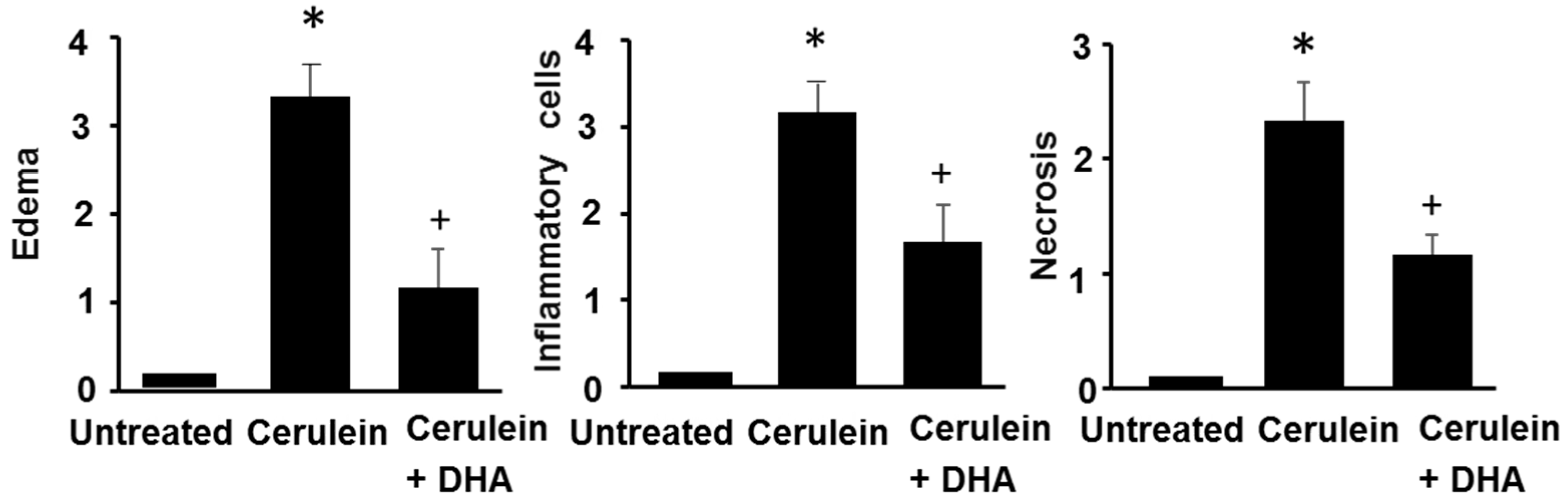
3.1. DHA Reduced Cerulein-Induced Pancreatic Edema in Rats
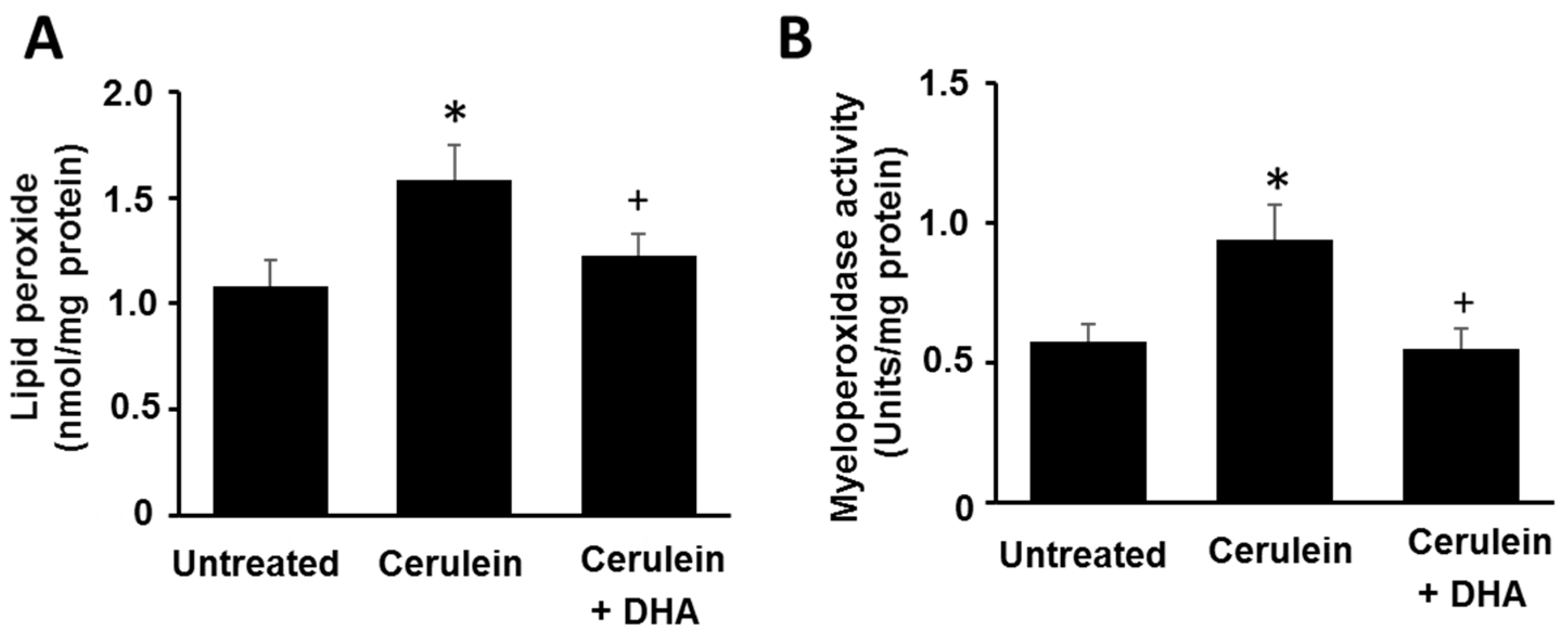
3.2. DHA Reduced the Abundance of LPO and Activity of MPO in Pancreas of Cerulein-Stimulated Rats
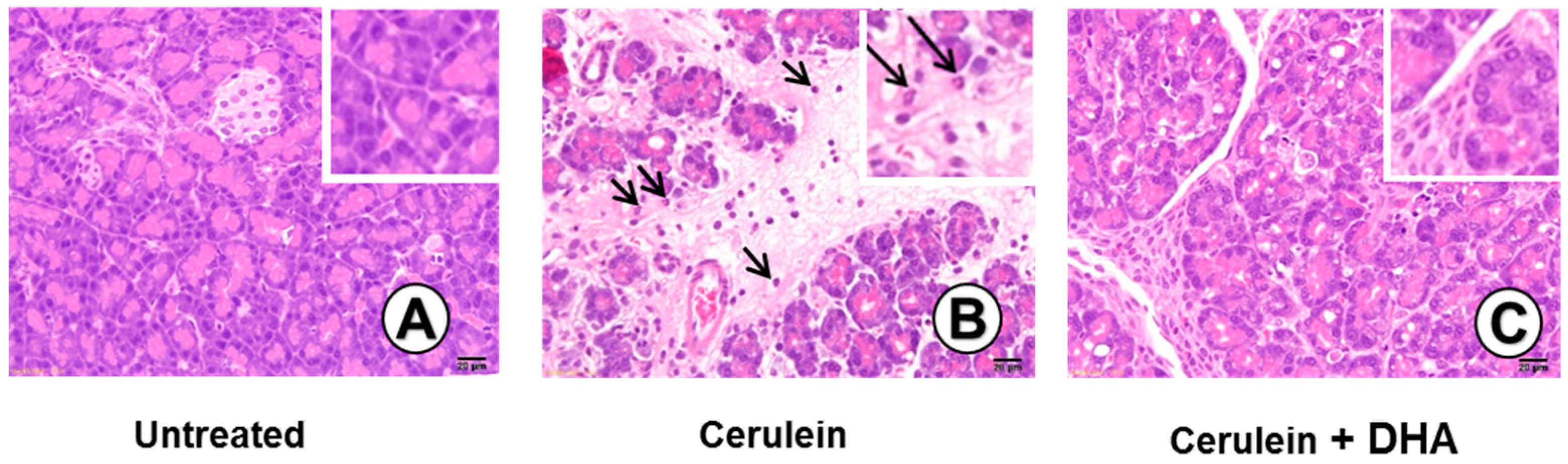
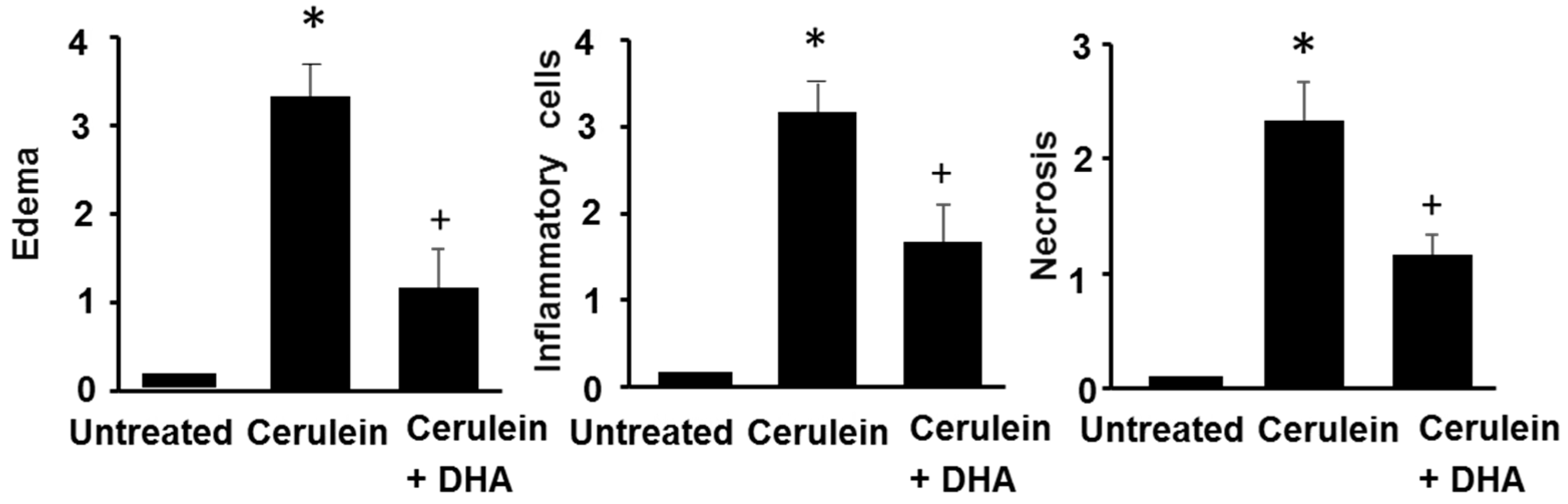
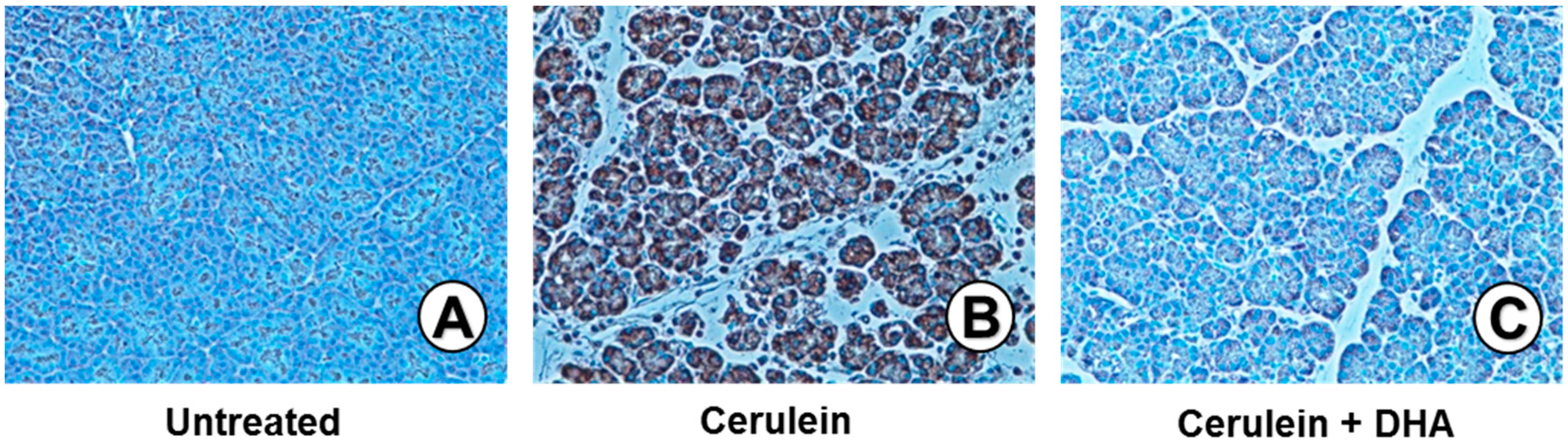
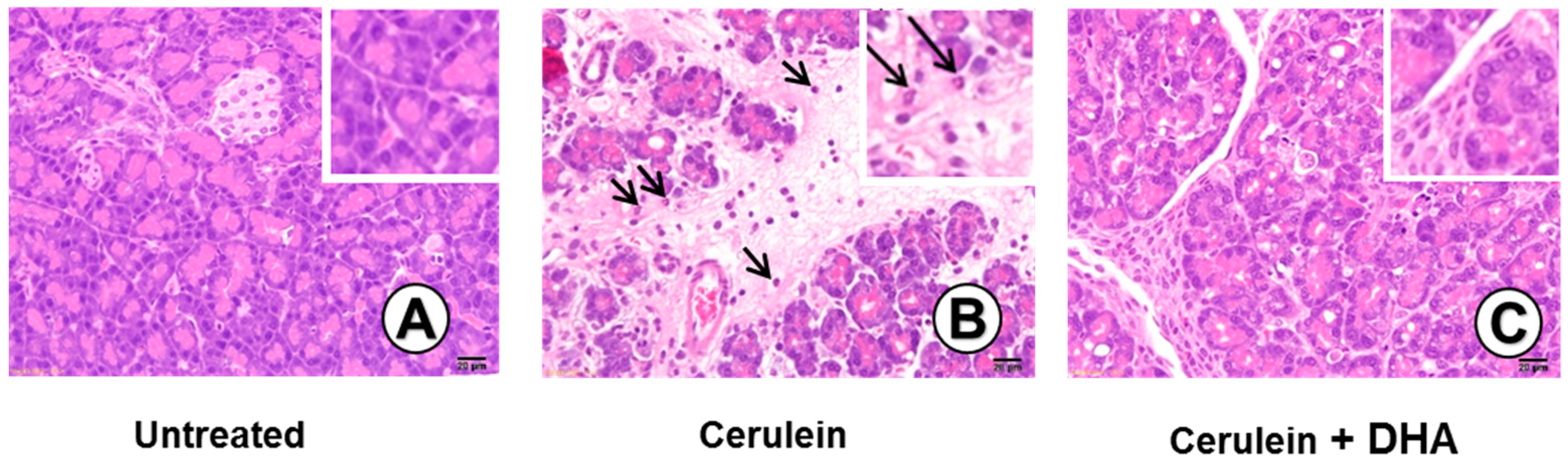
3.3. DHA Inhibited Cerulein-Induced Histopathologic Changes in Rat Pancreas
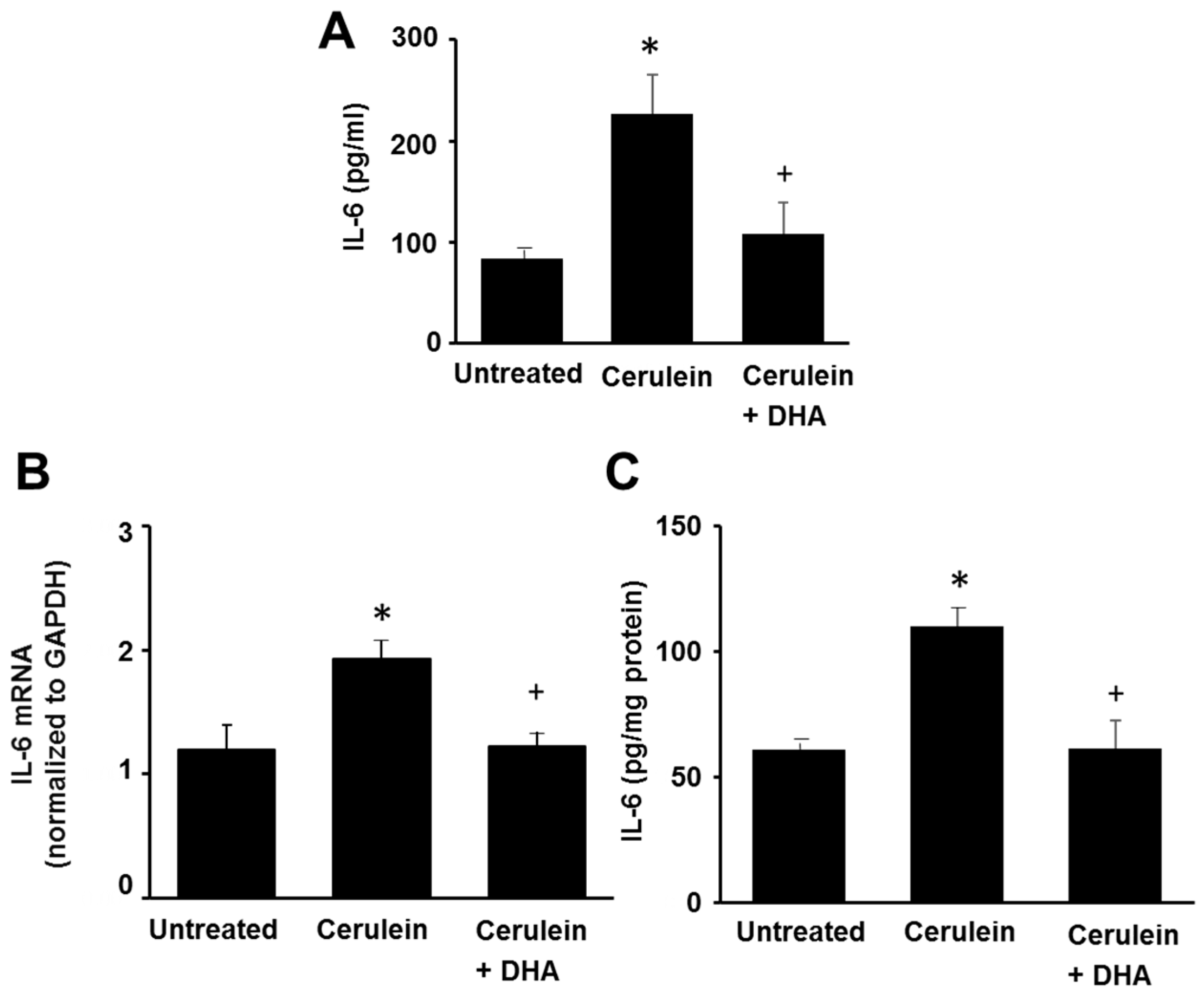
3.4. DHA Reduced IL-6 Expression in the Pancreas of Cerulein-Induced Rats
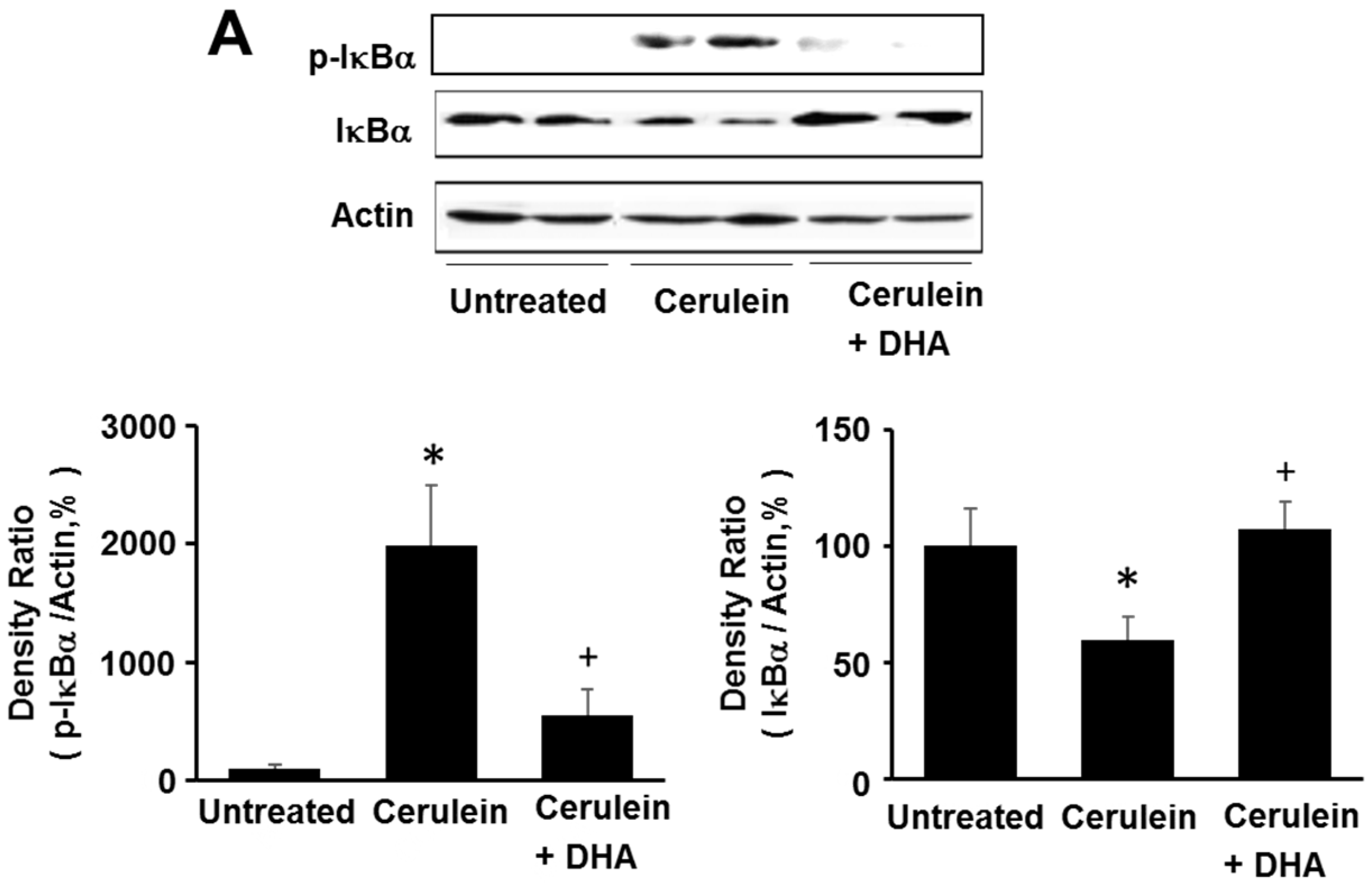
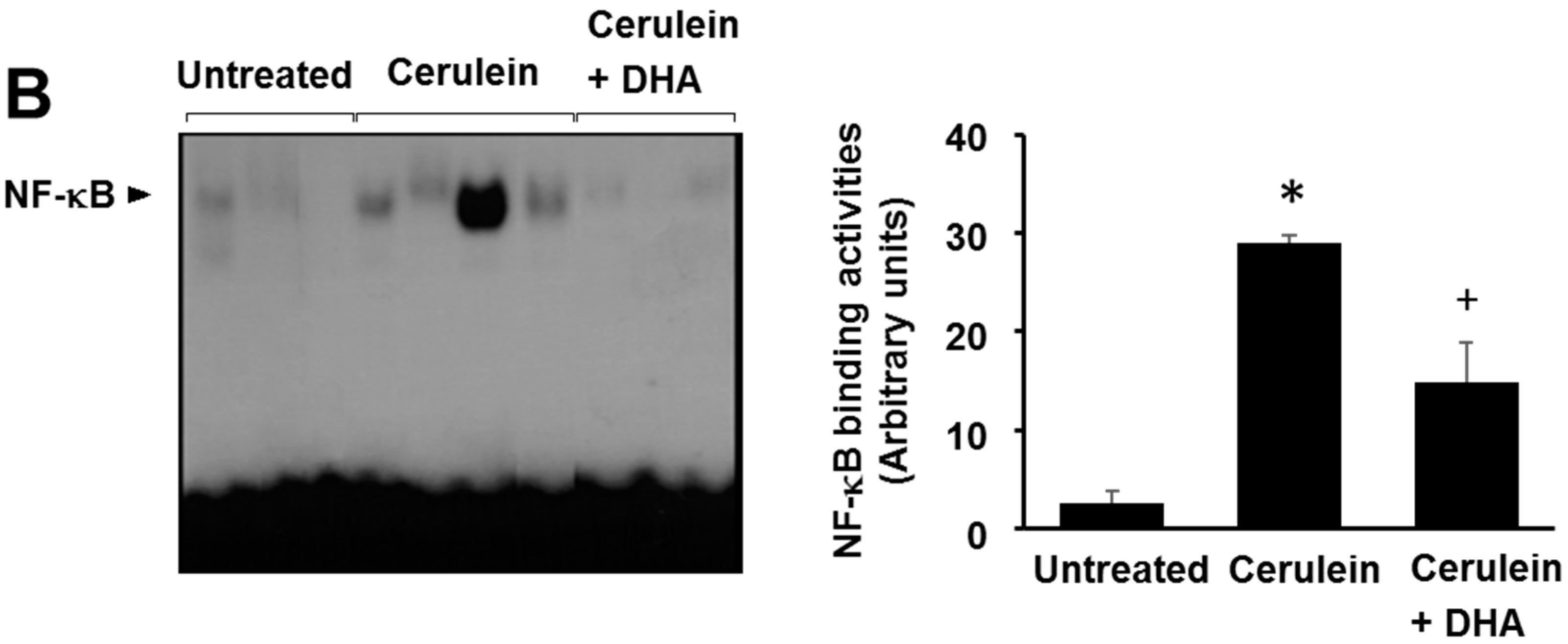
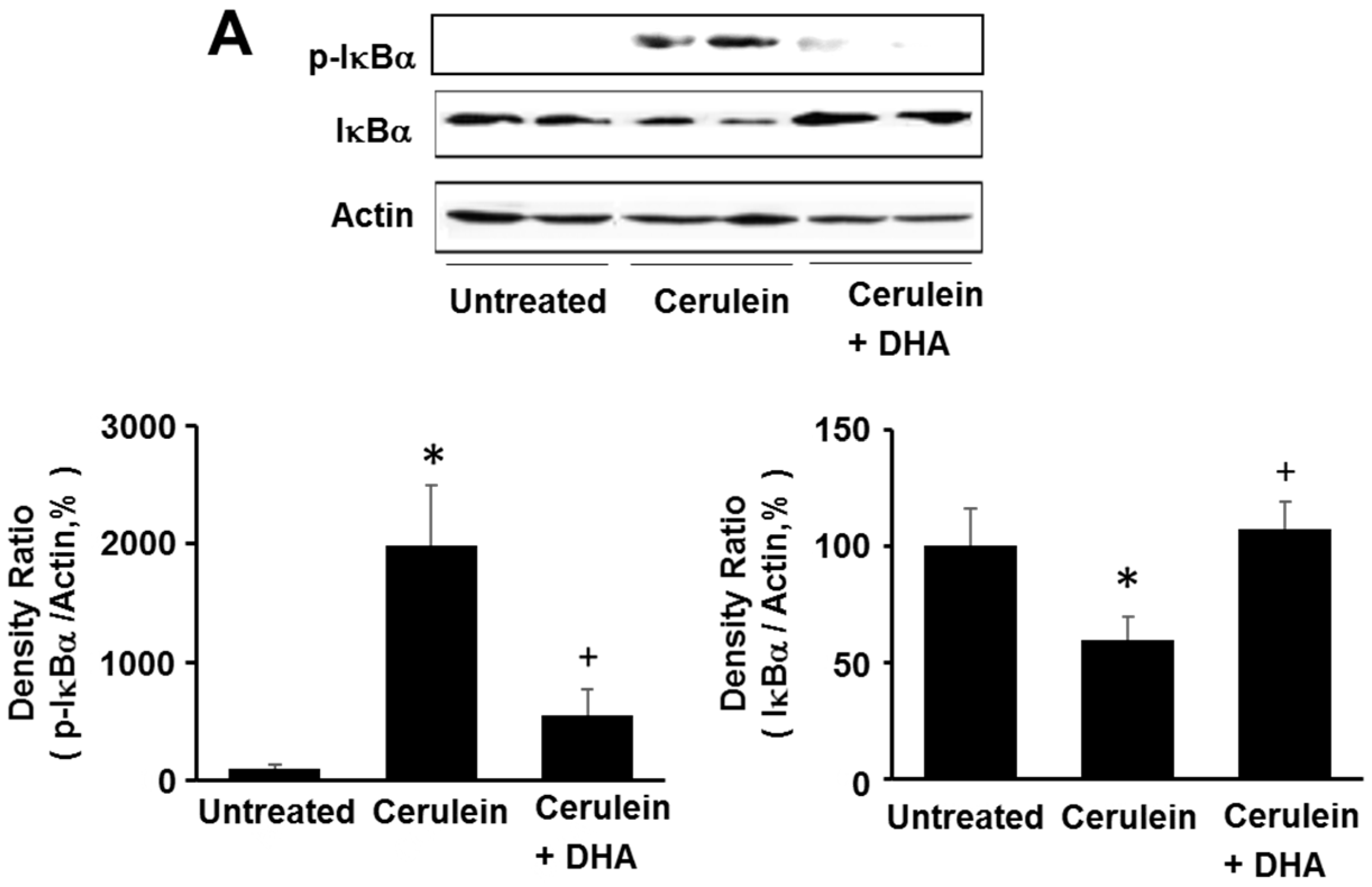
3.5. DHA Inhibited Cerulein-Induced Phosphorylation and Degradation of IκBα and Activation of NF-kB in the Rat Pancreas
3.6. DHA Suppressed Cerulein-Induced Activation of PKCδ in Rat Pancreas
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bhatia, M.; Wong, F.L.; Cao, Y.; Lau, H.Y.; Huang, J.; Puneet, P.; Chevali, L. Pathophysiology of acute pancreatitis. Pancreatology 2005, 5, 132–144. [Google Scholar] [CrossRef] [PubMed]
- Saluja, A.K.; Steer, M.L. Pathophysiology of pancreatitis. Digestion 1999, 60, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Frossard, J.L.; Hadengue, A.; Pastor, C.M. New serum markers for the detection of severe acute pancreatitis in humans. Am. J. Respir. Crit. Care Med. 2001, 164, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, B.; Saluja, A.K.; Lerch, M.M.; Bhagat, L.; Bhatia, M.; Lee, H.S.; Frossard, J.L.; Adler, G.; Steer, M.L. Intra-acinar cell activation of trypsinogen during caerulein-induced pancreatitis in rats. Am. J. Physiol. 1998, 275, 352–362. [Google Scholar]
- Lerch, M.M.; Adler, G. Experimental animal models of acute pancreatitis. Int. J. Pancreatol. 1994, 15, 159–170. [Google Scholar] [PubMed]
- Kim, H. Cerulein pancreatitis: Oxidative stress, inflammation, and apoptosis. Gut Liver 2008, 2, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Schoenberg, M.H.; Büchler, M.; Gaspar, M.; Stinner, A.; Younes, M.; Melzner, I.; Bültmann, B.; Beger, H.G. Oxygen free radicals in acute pancreatitis of the rat. Gut 1990, 31, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Gough, D.B.; Boyle, B.; Joyce, W.P.; Delaney, C.P.; McGeeney, K.F.; Gorey, T.F.; Fitzpatrick, J.M. Free radical inhibition and serial chemiluminescence in evolving experimental pancreatitis. Br. J. Surg. 1990, 77, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Niki, E.; Eguchi, J.; Kamiya, Y.; Shimasaki, H. Oxidation of biological membranes and its inhibition. Free radical chain oxidation of erythrocyte ghost membranes by oxygen. Biochim. Biophys. Acta 1985, 819, 29–36. [Google Scholar] [CrossRef]
- Koster, J.F.; Slee, R.G. Lipid peroxidation of human erythrocyte ghosts induced by organic hydroperoxides. Biochim. Biophys. Acta 1983, 752, 233–239. [Google Scholar] [CrossRef]
- De Groot, H.; Littauer, A. Hypoxia, reactive oxygen, and cell injury. Free Radic. Biol. Med. 1989, 6, 541–551. [Google Scholar] [CrossRef]
- Farber, J.L.; Kyle, M.E.; Coleman, J.B. Mechanisms of cell injury by activated oxygen species. Lab. Investig. 1990, 62, 670. [Google Scholar] [PubMed]
- Gómez-Cambronero, L.; Camps, B.; de La Asunción, J.G.; Cerdá, M.; Pellín, A.; Pallardó, F.V.; Calvete, J.; Sweiry, J.H.; Mann, G.E.; Viña, J.; et al. Pentoxifylline ameliorates cerulein-induced pancreatitis in rats: Role of glutathione and nitric oxide. J. Pharmacol. Exp. Ther. 2000, 293, 670–676. [Google Scholar] [PubMed]
- Alsfasser, G.; Gock, M.; Herzog, L.; Gebhard, M.M.; Herfarth, C.; Klar, E.; Schmidt, J. Glutathione depletion with l-buthionine-(S,R)-sulfoximine demonstrates deleterious effects in acute pancreatitis of the rat. Dig. Dis. Sci. 2002, 47, 1793–1799. [Google Scholar] [CrossRef] [PubMed]
- Curran, F.J.; Sattar, N.; Talwar, D.; Baxter, J.N.; Imrie, C.W. Relationship of carotenoid and vitamins A and E with the acute inflammatory response in acute pancreatitis. Br. J. Surg. 2000, 87, 301–305. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.J.; Mitros, F.A.; Oberley, L.W. Expression of antioxidant enzymes in diseases of the human pancreas: Another link between chronic pancreatitis and pancreatic cancer. Pancreas 2003, 26, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Schölmerich, J. Interleukins in acute pancreatitis. Scand. J. Gastroenterol. Suppl. 1996, 219, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Leser, H.G.; Gross, V.; Scheibenbogen, C.; Heinisch, A.; Salm, R.; Lausen, M.; Rückauer, K.; Andreesen, R.; Farthmann, E.H.; Schölmerich, J. Elevation of serum interleukin-6 concentration precedes acute-phase response and reflects severity in acute pancreatitis. Gastroenterology 1991, 101, 782–785. [Google Scholar] [CrossRef]
- Yu, J.H.; Lim, J.W.; Kim, H.; Kim, K.H. NADPH oxidase mediates interleukin-6 expression in cerulein-stimulated pancreatic acinar cells. Int. J. Biochem. Cell Biol. 2005, 37, 1458–1469. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Kim, K.H.; Kim, H. SOCS 3 and PPAR-lignads inhibits the expression of IL-6 and TGF-β by regulating JAK2/STAT3 signaling in pancreas. Int. J. Biochem. Cell Biol. 2008, 40, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, E.C.; Newton, A.C.; Mochly-Rosen, D.; Fields, A.P.; Reyland, M.E.; Insel, P.A.; Messing, R.O. Protein kinase C isozymes and the regulation of diverse cell responses. Am. J. Physiol. Lung Cell. Mol. Physiol. 2000, 279, L429–L438. [Google Scholar] [PubMed]
- Ron, D.; Kazanietz, M.G. New insights into the regulation of protein kinase C and novel phorbol ester receptors. FASEB J. 1999, 13, 1658–1676. [Google Scholar] [PubMed]
- Bastani, B.; Yang, L.; Baldassare, J.J.; Pollo, D.A.; Gardner, J.D. Cellular distribution of isoforms of protein kinase C (PKC) in pancreatic acini. Biochim. Biophys. Acta 1995, 1269, 307–315. [Google Scholar] [CrossRef]
- Pollo, D.A.; Baldassare, J.J.; Honda, T.; Henderson, P.A.; Talkad, V.D.; Gardner, J.D. Effects of cholecystokinin (CCK) and other secretagogues on isoforms of protein kinase C (PKC) in pancreatic acini. Biochim. Biophys. Acta 1994, 1224, 127–138. [Google Scholar] [CrossRef]
- Satoh, A.; Gukovskaya, A.S.; Nieto, J.M.; Cheng, J.H.; Gukovsky, I.; Reeve, J.R., Jr.; Shimosegawa, T.; Pandol, S.J. PKC-delta and -epsilon regulate NF-kappaB activation induced by cholecystokinin and TNF-alpha in pancreatic acinar cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Moyad, M.A. An introduction to dietary/supplemental omega-3 fatty acids for general health and prevention: Part II. Urol. Oncol. 2005, 23, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Lim, J.W.; Kim, H. Inhibitory mechanism of omega-3 fatty acids in pancreatic inflammation and apoptosis. Ann. N. Y. Acad. Sci. 2009, 1171, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Malleo, G.; Mazzon, E.; Genovese, T.; Di Paola, R.; Muià, C.; Centorrino, T.; Siriwardena, A.K.; Cuzzocrea, S. Etanercept attenuates the development of cerulein-induced acute pancreatitis in mice: A comparison with TNF-alpha genetic deletion. Shock 2007, 27, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Bettaieb, A.; Chahed, S.; Bachaalany, S.; Griffey, S.; Hammock, B.D.; Haj, F.G. Soluble epoxide hydrolase pharmacological inhibition ameliorates experimental acute pancreatitis in mice. Mol. Pharmacol. 2015, 88, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Qui, B.; Mei, Q.B.; Ma, J.J.; Korsten, M.A. Susceptibility to cerulein-induced pancreatitis in inducible nitric oxide synthase-deficient mice. Pancreas 2001, 23, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Cuzzocrea, S.; Rossi, A.; Serraino, I.; Di Paola, R.; Dugo, L.; Genovese, T.; Britti, D.; Sciarra, G.; De Sarro, A.; Caputi, A.P.; et al. 5-Lipoxygenase knockout mice exhibit a resistance to acute pancreatitis induced by cerulein. Immunology 2003, 110, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Krawisz, J.E.; Sharon, P.; Stenson, W.F. Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 1984, 87, 1344–1350. [Google Scholar] [PubMed]
- Chen, P.; Sun, B.; Chen, H.; Wang, G.; Pan, S.; Kong, R.; Bai, X.; Wang, S. Effects of carbon monoxide releasing molecule-liberated CO on severe acute pancreatitis in rats. Cytokine 2010, 49, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.; Rattner, D.W.; Lewandrowski, K.; Compton, C.C.; Mandavilli, U.; Knoefel, W.T.; Warshaw, A.L. A better model of acute pancreatitis for evaluating therapy. Ann. Surg. 1992, 215, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice Hall: New Jersey, NY, USA, 1984. [Google Scholar]
- Siriwardhana, N.; Kalupahana, N.S.; Moustaid-Moussa, N. Health benefits of n-3 polyunsaturated fatty acids: eicosapentaenoic acid and docosahexaenoic acid. Adv. Food Nutr. Res. 2012, 65, 211–222. [Google Scholar] [PubMed]
- Riediger, N.D.; Othman, R.A.; Suh, M.; Moghadasian, M.H. A systemic review of the roles of n-3 fatty acids in health and disease. J. Am. Diet Assoc. 2009, 109, 668–679. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta 2015, 1851, 469–484. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Chung, H.Y. Antioxidative and anti-inflammatory actions of docosahexaenoic acid and eicosapentaenoic acid in renal epithelial cells and macrophages. J. Med. Food 2007, 10, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Véricel, E.; Polette, A.; Bacot, S.; Calzada, C.; Lagarde, M. Pro- and antioxidant activities of docosahexaenoic acid on human blood platelets. J. Thromb. Haemost. 2005, 1, 566–572. [Google Scholar] [CrossRef]
- Song, E.A.; Lim, J.W.; Kim, H. Docosahexaenoic acid inhibits IL-6 expression via PPARγ-mediated expression of catalase in cerulein-stimulated pancreatic acinar cells. Int. J. Biochem. Cell Biol. 2017, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Das, M.; Das, S. Identification of cytotoxic mediators and their putative role in the signaling pathways during docosahexaenoic acid (DHA)-induced apoptosis of cancer cells. Apoptosis 2016, 21, 1408–1421. [Google Scholar] [CrossRef] [PubMed]
- Merendino, N.; Loppi, B.; D’Aquino, M.; Molinari, R.; Pessina, G.; Romano, C.; Velotti, F. Docosahexaenoic acid induces apoptosis in the human PaCa-44 pancreatic cancer cell line by active reduced glutathione extrusion and lipid peroxidation. Nutr. Cancer 2005, 52, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Trépanier, M.O.; Lim, J.; Lai, T.K.; Cho, H.J.; Domenichiello, A.F.; Chen, C.T.; Taha, A.Y.; Bazinet, R.P.; Burnham, W.M. Intraperitoneal administration of docosahexaenoic acid for 14 days increases serum unesterified DHA and seizure latency in the maximal pentylenetetrazol model. Epilepsy Behav. 2014, 33, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Tsukamoto, A.; Sugimoto, T.; Onuki, Y.; Shinoda, H.; Mihara, T.; Hori, M.; Inomata, T. The 5-HT3 receptor antagonist ondansetron attenuates pancreatic injury in cerulein-induced acute pancreatitis model. Inflammation 2017, 40, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Li, L.; Liu, H.; Hu, L.; Dai, Y.; Chen, J.; Xu, S.; Chen, W.; Xu, X.; et al. Propylene glycol alginate sodium sulfate alleviates cerulein-induced acute pancreatitis by modulating the MEK/ERK pathway in mice. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [PubMed]
- Kosekli, M.A.; Sungurtekin, U.; Cobankara, V.; Ozmen, O.; Sahinduran, S.; Yilmaz, M. Effects of certolizumab on cerulein-induced acute pancreatitis in rats. Pancreas 2016, 45, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Hirota, M.; Nozawa, F.; Okabe, A.; Shibata, M.; Beppu, T.; Shimada, S.; Egami, H.; Yamaguchi, Y.; Ikei, S.; Okajima, T.; et al. Relationship between plasma cytokine concentration and multiple organ failure in patients with acute pancreatitis. Pancreas 2000, 21, 141–146. [Google Scholar] [CrossRef] [PubMed]
- Panday, A.; Sahoo, M.K.; Osorio, D.; Batra, S. NADPH oxidases: An overview from structure to innate immunity-associated pathologies. Cell. Mol. Immunol. 2015, 12, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Xu, B.; Xia, T.; He, W.; Gao, P.; Guo, L.; Wang, Z.; Niu, Q.; Wang, A. Relationship between intracellular Ca2⁺ and ROS during fluoride-induced injury in SH-SY5Y cells. Environ. Toxicol. 2013, 28, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Lim, J.W.; Kim, K.H.; Morio, T.; Kim, H. NADPH oxidase and apoptosis in cerulein-stimulated pancreatic acinar AR42J cells. Free Radic. Biol. Med. 2005, 39, 590–602. [Google Scholar] [CrossRef] [PubMed]
- Gukovskaya, A.S.; Hosseini, S.; Satoh, A.; Cheng, J.H.; Nam, K.J.; Gukovsky, I.; Pandol, S.J. Ethanol differentially regulates NF-kappaB activation in pancreatic acinar cells through calcium and protein kinase C pathways. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 286, G204–G213. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Logsdon, C.D. CCK stimulates mob-1 expression and NF-kappaB activation via protein kinase C and intracellular Ca2+. Am. J. Physiol. Cell Physiol. 2000, 278, C344–C351. [Google Scholar] [PubMed]
- Yuan, J.; Lugea, A.; Zheng, L.; Gukovsky, I.; Edderkaoui, M.; Rozengurt, E.; Pandol, S.J. Protein kinase D1 mediates NF-kappaB activation induced by cholecystokinin and cholinergic signaling in pancreatic acinar cells. Am. J. Physiol. Gastrointest. Liver Physiol. 2008, 295, G1190–G1201. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, X.; Westlund, K.N. Restoration of spontaneous exploratory behaviors with an intrathecal NMDA receptor antagonist or a PKC inhibitor in rats with acute pancreatitis. Pharmacol. Biochem. Behav. 2004, 77, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Jakkampudi, A.; Jangala, R.; Reddy, B.R.; Mitnala, S.; Nageshwar Reddy, D.; Talukdar, R. NF-κB in acute pancreatitis: Mechanisms and therapeutic potential. Pancreatology 2016, 16, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.E.; Lim, J.W.; Kim, J.M.; Kim, H. Anti-inflammatory mechanism of polyunsaturated fatty acids in Helicobacter pylori-infected gastric epithelial cells. Mediat. Inflamm. 2014, 128919. [Google Scholar] [CrossRef]
- Lian, S.; Xia, Y.; Nguyen, T.T.; Ung, T.T.; Yoon, H.J.; Kim, N.H.; Kim, K.K.; Jung, Y.D. Docosahexaenoic Acid Inhibits Tumor Promoter-Induced Urokinase-Type Plasminogen Activator Receptor by Suppressing PKCδ- and MAPKs-Mediated Pathways in ECV304 Human Endothelial Cells. PLoS ONE 2016, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Dunn, J.A.; Li, C.; Ha, T.; Kao, R.L.; Browder, W. Therapeutic modification of nuclear factor kappa B binding activity and tumor necrosis factor-alpha gene expression during acute biliary pancreatitis. Am. Surg. 1997, 63, 1036–1043. [Google Scholar] [PubMed]
- Grady, T.; Liang, P.; Ernst, S.A.; Logsdon, C.D. Chemokine gene expression in rat pancreatic acinar cells is an early event associated with acute pancreatitis. Gastroenterology 1997, 113, 1966–1975. [Google Scholar] [CrossRef]
- Gukovsky, I.; Gukovskaya, A.S.; Blinman, T.A.; Zaninovic, V.; Pandol, S.J. Early NF-kappaB activation is associated with hormone-induced pancreatitis. Am. J. Physiol. 1998, 275, G1402–G1414. [Google Scholar] [PubMed]
- Satoh, A.; Shimosegawa, T.; Fujita, M.; Kimura, K.; Masamune, A.; Koizumi, M.; Toyota, T. Inhibition of nuclear factor-kappaB activation improves the survival of rats with taurocholate pancreatitis. Gut 1999, 44, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Ji, B.; Han, B.; Ernst, S.A.; Simeone, D.; Logsdon, C.D. NF-kappaB activation in pancreas induces pancreatic and systemic inflammatory response. Gastroenterology 2002, 122, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Baumann, B.; Wagner, M.; Aleksic, T.; von Wichert, G.; Weber, C.K.; Adler, G.; Wirth, T. Constitutive IKK2 activation in acinar cells is sufficient to induce pancreatitis in vivo. J. Clin. Investig. 2007, 117, 1502–1513. [Google Scholar] [CrossRef] [PubMed]
- Algül, H.; Treiber, M.; Lesina, M.J.; Nakhai, H.; Saur, D.; Geisler, F.; Pfeifer, A.; Paxian, S.; Schmid, R.M. Pancreas-specific RelA/p65 truncation increases susceptibility of acini to inflammation-associated cell death following cerulein pancreatitis. J. Clin. Investig. 2007, 117, 1490–1501. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer | Sequence (5′-3′) |
|---|---|---|
| GAPDH | Forward primer | GAAGGTGAAGGTCGGAGT |
| Reverse primer | GAAGATGGTGATGGGATTC | |
| IL-6 | Forward primer | GAGAGGAGACTTCACAGAGGATACCAC |
| Reverse primer | ACCACAGTGAGGAATGTCCACAA |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, Y.K.; Lee, S.; Lim, J.W.; Kim, H. Docosahexaenoic Acid Inhibits Cerulein-Induced Acute Pancreatitis in Rats. Nutrients 2017, 9, 744. https://doi.org/10.3390/nu9070744
Jeong YK, Lee S, Lim JW, Kim H. Docosahexaenoic Acid Inhibits Cerulein-Induced Acute Pancreatitis in Rats. Nutrients. 2017; 9(7):744. https://doi.org/10.3390/nu9070744
Chicago/Turabian StyleJeong, Yoo Kyung, Sle Lee, Joo Weon Lim, and Hyeyoung Kim. 2017. "Docosahexaenoic Acid Inhibits Cerulein-Induced Acute Pancreatitis in Rats" Nutrients 9, no. 7: 744. https://doi.org/10.3390/nu9070744
APA StyleJeong, Y. K., Lee, S., Lim, J. W., & Kim, H. (2017). Docosahexaenoic Acid Inhibits Cerulein-Induced Acute Pancreatitis in Rats. Nutrients, 9(7), 744. https://doi.org/10.3390/nu9070744