Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves

 ,
, 

Abstract
:1. Introduction
2. Results and Discussion
2.1. Inhibitory Effects of Crude Extracts of Rosemary Organs on Lettuce Growth
2.2. The Content of Pure Compounds in Rosemary Organs
2.3. Plant Growth Inhibitory Effects of Pure Compounds Present in Rosemary
2.4. Estimation of Contributions of the Various Compounds to Rosemary Growth Inhibitory Activity
2.5. Growth Inhibition and Changes in Filter Paper Coloration by Carnosic Acid
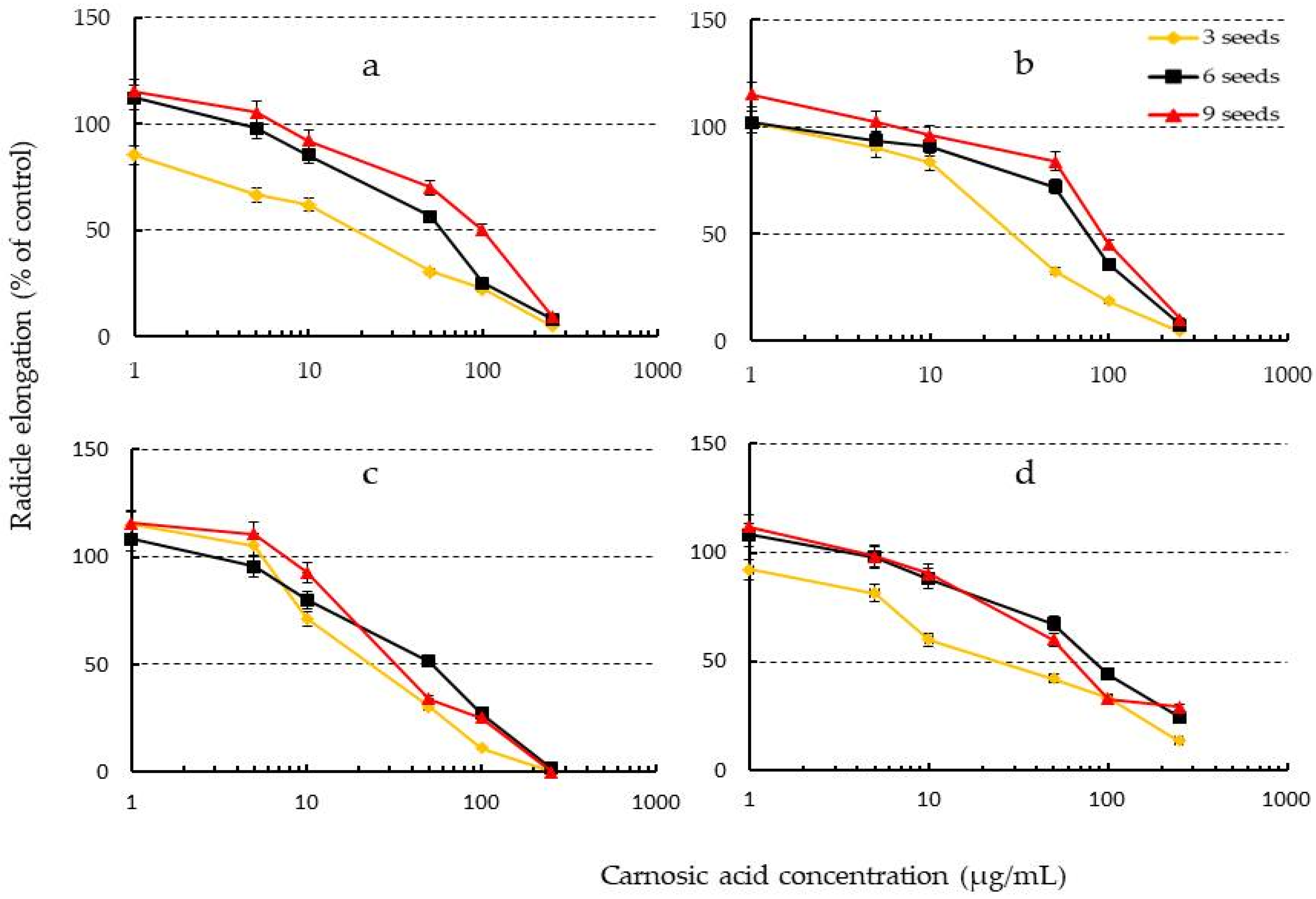
2.6. Effects of Increasing Seed Number of Test Plants on Phytotoxicity of Carnosic Acid
2.7. Effect of Soil-Incorporated with Rosemary Leaf Debris on the Bioassay Species
3. Conclusions
4. Materials and Methods
4.1. Chemicals and Test Plants for Bioassay
4.2. Selection of Rosemary Samples and Extraction Procedure
4.3. Inhibitory Effects of Rosemary Crude Extract and Test Compounds
4.4. HPLC Analysis of Rosemary Organs
4.5. Growth Inhibition and Filter Paper Coloration Changes Induced by Carnosic Acid
4.6. Effects on Growth Inhibition Activity of Carnosic Acid by Varied Seed Number of Test Plants
4.7. Effect of Rosemary Leaf Debris Incorporated Soil on the Bioassay Species
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Willis, R.J. What Is Allelopathy? In The History of Allelopathy; Springer: Dordrecht, The Netherlands, 2007; pp. 1–13. [Google Scholar]
- Inderjit; Weston, L.; Duke, S. Challenges, Achievements and Opportunities in Allelopathy Research. J. Plant Interact. 2005, 1, 69–81. [Google Scholar] [CrossRef]
- Weir, T.L.; Park, S.W.; Vivanco, J.M. Biochemical and Physiological Mechanisms Mediated by Allelochemicals. Curr. Opin. Plant Biol. 2004, 7, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Xuan, T.D.; Shinkichi, T.; Khanh, T.D.; Chung, I.M. Biological Control of Weeds and Plant Pathogens in Paddy Rice by Exploiting Plant Allelopathy: An Overview. Crop Prot. 2005, 24, 197–206. [Google Scholar] [CrossRef]
- Fujii, Y.; Shibuya, T.; Tamaki, Y. Allelopathy of Velvetbean: Determination and Identification of L-DOPA as a Candidate of Allelopathic Substances. Japan Agric. Res. Q. 1992, 25, 238–247. [Google Scholar]
- Kamo, T.; Hiradate, S.; Fujii, Y. First Isolation of Natural Cyanamide as a Possible Allelochemical from Hairy Vetch Vicia villosa. J. Chem. Ecol. 2003, 29, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Hiradate, S.; Ohse, K.; Furubayashi, A.; Fujii, Y. Quantitative Evaluation of Allelopathic Potentials in Soils: Total Activity Approach. Weed Sci. 2010, 58, 258–264. [Google Scholar] [CrossRef]
- Golisz, A.; Lata, B.; Gawronski, S.W.; Fujii, Y. Specific and Total Activities of the Allelochemicals Identified in Buckwheat. Weed Biol. Manag. 2007, 7, 164–171. [Google Scholar] [CrossRef]
- Mishyna, M.; Laman, N.; Prokhorov, V.; Fujii, Y. Angelicin as the Principal Allelochemical in Heracleum sosnowskyi Fruit. Nat. Prod. Commun. 2015, 10, 1–5. [Google Scholar]
- Vyvyan, J.R. Allelochemicals as Leads for New Herbicides and Agrochemicals. Tetrahedron 2002, 58, 1631–1646. [Google Scholar] [CrossRef]
- Barney, J.N.; Hay, A.G.; Weston, L.A. Isolation and Characterization of Allelopathic Volatiles from Mugwort (Artemisia vulgaris). J. Chem. Ecol. 2005, 31, 247–265. [Google Scholar] [CrossRef] [PubMed]
- Weston, L.A.; Duke, S.O. Weed and Crop Allelopathy. CRC. Crit. Rev. Plant Sci. 2003, 22, 367–389. [Google Scholar] [CrossRef]
- Soltys, D.; Krasuska, U.; Bogatek, R.; Gniazdowsk, A. Allelochemicals as Bioherbicides—Present and Perspectives. Herbic. Curr. Res. Case Stud. Use 2013. [Google Scholar] [CrossRef]
- Heap, I. Global Perspective of Herbicide-Resistant Weeds. Global perspective of herbicide-resistant weeds. Pest Manag. Sci. 2014, 70, 1306–1315. [Google Scholar] [CrossRef] [PubMed]
- Duke, S.O.; Romagni, J.G.; Dayan, F.E. Natural Products as Sources for New Mechanisms of Herbicidal Action. Crop Prot. 2000, 19, 583–589. [Google Scholar] [CrossRef]
- Dayan, F.E.; Owens, D.K.; Duke, S.O. Rationale for a Natural Products Approach to Herbicide Discovery. Pest Manag. Sci. 2012, 68, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Gerwick, B.C.; Sparks, T.C. Natural Products for Pest Control: An Analysis of Their Role, Value and Future. Pest Manag. Sci. 2014, 70, 1169–1185. [Google Scholar] [CrossRef] [PubMed]
- Terpinc, P.; Bezjak, M.; Abramovič, H. A Kinetic Model for Evaluation of the Antioxidant Activity of Several Rosemary Extracts. Food Chem. 2009, 115, 740–744. [Google Scholar] [CrossRef]
- Napoli, E.M.; Curcuruto, G.; Ruberto, G. Screening of the Essential Oil Composition of Wild Sicilian Rosemary. Biochem. Syst. Ecol. 2010, 38, 659–670. [Google Scholar] [CrossRef]
- Kontogianni, V.G.; Tomic, G.; Nikolic, I.; Nerantzaki, A.A.; Sayyad, N.; Stosic-Grujicic, S.; Stojanovic, I.; Gerothanassis, I.P.; Tzakos, A.G. Phytochemical Profile of Rosmarinus officinalis and Salvia officinalis Extracts and Correlation to their Antioxidant and Anti-Proliferative Activity. Food Chem. 2013, 136, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Sotelo-Felix, J.I.; Matinez-Fong, D.; Muriel, P.; Santillan, R.L.; Castillo, D.; Yahuaca, P. Evaluation of the Effectiveness of Rosmarinus officinalis (Lamiaceae) in the Alleviation of Carbon Tetrachloride-Induced Acute Hepatotoxicity. J. Ethnopharmacol. 2002, 81, 145–154. [Google Scholar] [CrossRef]
- Sasaki, K.; El Omri, A.; Kondo, S.; Han, J.; Isoda, H. Rosmarinus officinalis Polyphenols Produce Anti-Depressant like Effect through Monoaminergic and Cholinergic Functions Modulation. Behav. Brain Res. 2013, 238, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Bakirel, T.; Bakirel, U.; Keleş, O.Ü.; Ülgen, S.G.; Yardibi, H. In Vivo Assessment of Antidiabetic and Antioxidant Activities of Rosemary (Rosmarinus officinalis) in Alloxan-Diabetic Rabbits. J. Ethnopharmacol. 2008, 116, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Appiah, K.; Mardani, H.; Osivand, A.; Kpabitey, S.; Amoatey, C.; Oikawa, Y.; Fujii, Y. Exploring Alternative Use of Medicinal Plants for Sustainable Weed Management. Sustainability 2017, 9, 1468. [Google Scholar] [CrossRef]
- Takemura, T.; Sakuno, E.; Kamo, T.; Hiradate, S.; Fujii, Y. Screening of the Growth-Inhibitory Effects of 168 Plant Species against Lettuce Seedlings. Am. J. Plant Sci. 2013, 04, 1095–1104. [Google Scholar] [CrossRef]
- Morikawa, C.I.O.; Miyaura, R.; Kamo, T.; Hiradate, S.; Chávez Pérez, J.A.; Fujii, Y. Isolation of Umbelliferone as a Principal Allelochemical from the Peruvian Medicinal Plant Diplostephium foliosissimum (Asteraceae). Rev. Soc. Quim. Peru. 2011, 77, 285–291. [Google Scholar]
- Rahimi, M.; Bidarnamani, F.; Shabanipoor, M. Effects of Allelopathic Three Medicinal Plants on Germination and Seeding Growth of Portulaca oleracea. Forum 2015, 7, 1520–1523. [Google Scholar]
- Hiradate, S.; Morita, S.; Sugie, H.; Fujii, Y.; Harada, J. Phytotoxic Cis-Cinnamoyl Glucosides from Spiraea thunbergii. Phytochemistry 2004, 65, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Hiradate, S.; Morita, S.; Furubayashi, A.; Fujii, Y.; Harada, J. Plant Growth Inhibition by Cis-Cinnamoyl Glucosides and Cis-Cinnamic Acid. J. Chem. Ecol. 2005, 31, 591–601. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Hiradate, S. A Critical Survey of Allelochemicals in Action—the Importance of Total Activity and the Weed Suppression Equation. In Establishing the Scientific Base; Harper, J.D.I., An, M., Wu, H., Kent, J.H., Eds.; Centre for Rural Social Research, Charles Sturt University: Wagga Wagga, Australia, 2005; pp. 73–76. [Google Scholar]
- Belel, M.D.; Belel, R.D. Allelopathic Effect of Leaf and Seed Extract of Nutgrass (Cyperus tuberosus) on the Germination of Beans (Vigna unguiculata (L.) Walp). Cogent Food Agric. 2015, 1, 1–7. [Google Scholar] [CrossRef]
- Cipollini, K.; Greenawalt Bohrer, M. Comparison of Allelopathic Effects of Five Invasive Species on Two Native Species. J. Torrey Bot. Soc. 2016, 143, 427–436. [Google Scholar] [CrossRef]
- Wu, A.P.; Li, Z.L.; He, F.F.; Wang, Y.H.; Dong, M. Screening Allelochemical-Resistant Species of the Alien Invasive Mikania micrantha for Restoration in South China. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Cheng, Z.; Meng, H.; Tang, X. The Garlic Allelochemical Diallyl Disulfide Affects Tomato Root Growth by Influencing Cell Division, Phytohormone Balance and Expansin Gene Expression. Front. Plant Sci. 2016, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Turk, M.A.; Tawaha, A.M. Allelopathic Effect of Black Mustard (Brassica nigra L.) on Germination and Growth of Wild Oat (Avena fatua L.). Crop Prot. 2003, 22, 673–677. [Google Scholar] [CrossRef]
- Takemura, T.; Kamo, T.; Sakuno, E.; Hiradate, S.; Fujii, Y. Discovery of Coumarin as the Predominant Allelochemical in Gliricidia sepium. J. Trop. For. Sci. 2013, 25, 268–272. [Google Scholar]
- Kobayashi, A.; Kato-Noguchi, H. The Seasonal Variations of Allelopathic Activity and Allelopathic Substances in Brachiaria brizantha. Bot. Stud. 2015, 56, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Hu, H. Isolation and Characterization of a Novel Antialgal Allelochemical from Phragmites communis. J. Appl. Environ. Microbiol. 2005, 71, 6545–6553. [Google Scholar] [CrossRef] [PubMed]
- Del Baño, M.J.; Lorente, J.; Castillo, J.; Benavente-García, O.; Del Río, J.A.; Ortuño, A.; Quirin, K.W.; Gerard, D. Phenolic Diterpenes, Flavones, and Rosmarinic Acid Distribution during the Development of Leaves, Flowers, Stems, and Roots of Rosmarinus officinalis. Antioxidant Activity. J. Agric. Food Chem. 2003, 51, 4247–4253. [Google Scholar] [CrossRef] [PubMed]
- Luis, J.C.; Johnson, C.B. Seasonal Variations of Rosmarinic and Carnosic Acids in Rosemary Extracts. Analysis of Their in Vitro Antiradical Activity. Spanish J. Agric. Res. 2005, 3, 106–112. [Google Scholar] [CrossRef]
- Wellwood, C.R.L.; Cole, R.A. Relevance of Carnosic Acid Concentrations to the Selection of Rosemary, Rosmarinus officinalis (L.), Accessions for Optimization of Antioxidant Yield. J. Agric. Food Chem. 2004, 52, 6101–6107. [Google Scholar] [CrossRef] [PubMed]
- Borrás-Linares, I.; Stojanović, Z.; Quirantes-Piné, R.; Arráez-Román, D.; Švarc-Gajić, J.; Fernández-Gutiérrez, A.; Segura-Carretero, A. Rosmarinus officinalis Leaves as a Natural Source of Bioactive Compounds. Int. J. Mol. Sci. 2014, 15, 20585–20606. [Google Scholar] [CrossRef] [PubMed]
- Vallverdú-Queralt, A.; Regueiro, J.; Martínez-Huélamo, M.; Rinaldi Alvarenga, J.F.; Leal, L.N.; Lamuela-Raventos, R.M. A Comprehensive Study on the Phenolic Profile of Widely Used Culinary Herbs and Spices: Rosemary, Thyme, Oregano, Cinnamon, Cumin and Bay. Food Chem. 2014, 154, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Chung, I.M.; Ahn, J.K.; Yun, S.J. Identification of Allelopathic Compounds from Rice (Oryza sativa L.) Straw and Their Biological Activity. Can. J. Plant Sci. 2001, 81, 815–819. [Google Scholar] [CrossRef]
- Ishikura, Y.; Kojima, Y.; Terrazawa, M. Effects of Phenolic Compounds on Seed Germination of Shirakamba birch, Betula platyphylla Var. Japonica. Eurasian J. For. Res. 2001, 2, 17–25. [Google Scholar]
- Xuan, T.D.; Tsuzuki, E.; Terao, H.; Matsuo, M.; Khanh, T.D. Correlation between Growth Inhibitory Exhibition and Suspected Allelochemicals (Phenolic Compounds) in the Extract of Alfalfa (Medicago sativa L.). Plant Prod. Sci. 2003, 6, 165–171. [Google Scholar] [CrossRef]
- Chon, S.U.; Kim, J.D. Biological Activity and Quantification of Suspected Allelochemicals from Alfalfa Plant Parts. J. Agron. Crop Sci. 2002, 188, 281–285. [Google Scholar] [CrossRef]
- Masuda, T.; Inaba, Y.; Maekawa, T.; Takeda, Y.; Tamura, H.; Yamaguchi, H. Recovery Mechanism of the Antioxidant Activity from Carnosic Acid Quinone, an Oxidized Sage and Rosemary Antioxidant. J. Agric. Food Chem. 2002, 50, 5863–5869. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef] [Green Version]
- Ternes, W.; Schwarz, K. Antioxidative Constituents of Rosmarinus officinalis and Salvia officinalis. Z Leb. Unters Forch 1995, 201, 548–550. [Google Scholar] [CrossRef]
- Smith, R. Weed Control in Carrot Family Crops. 2015. Available online: https://ucanr.edu/blogs/blogcore/postdetail.cfm?postnum=17941 (accessed on 18 September 2018).
- Nishihara, E.; Parvez, M.M.; Araya, H.; Kawashima, S.; Fujii, Y. L-3-(3,4-Dihydroxyphenyl)Alanine (L-DOPA), an Allelochemical Exuded from Velvetbean (Mucuna pruriens) Roots. Plant Growth Regul. 2005, 45, 113–120. [Google Scholar] [CrossRef]
- Schwarz, K.; Ternes, W.; Schmauderer, E. Antioxidative Constituents of Rosmarinus officinalis and Salvia officinalis with Antioxidative Activity amongst Tocochromanols Using HPLC. Leb. Und-forsch. 1992, 104–107. [Google Scholar] [CrossRef]
- Masuda, T.; Inaba, Y.; Takeda, Y. Antioxidant Mechanism of Carnosic Acid: Structural Identification of Two Oxidation Products. J. Agric. Food Chem. 2001, 49, 5560–5565. [Google Scholar] [CrossRef] [PubMed]
- Mijangos, F.; Varona, F.; Villota, N. Changes in Solution Color during Phenol Oxidation by Fenton Reagent. Environ. Sci. Technol. 2006, 40, 5538–5543. [Google Scholar] [CrossRef] [PubMed]
- Phipyapong, P.; Hunt, M.D.; Steffens, J.C. Systemic Wound Induction of Potato (Solanum tuberosum) Polyphenol Oxidase. Phytochemistry 1995, 40, 673–676. [Google Scholar] [CrossRef]
- Storch, A.; Blessing, H.; Bareiss, M.; Jankowski, S.; Ling, Z.A.O.D.; Carvey, P.; Schwarz, J. Catechol-O-Methyltransferase Inhibition Attenuates Levodopa Toxicity in Mesencephalic Dopamine Neurons. Mol. Pharmacol. 2000, 594, 589–594. [Google Scholar]
- Cheema, Z.A.; Khaliq, A. Use of Sorghum Allelopathic Properties to Control Weeds in Irrigated Wheat in a Semi Arid Region of Punjab. Agric. Ecosyst. Environ. 2000, 79, 105–112. [Google Scholar] [CrossRef]
- Ismail, B.S.; Siddique, M.A.B. The Inhibitory Effect of Grasshopper’s Cyperus (Cyperus iria L.) on the Seedling Growth of Five Malaysian Rice Varieties. Trop. Life Sci. Res. 2011, 22, 81–89. [Google Scholar] [PubMed]
- Iqbal, Z.; Furubayashi, A.; Fujii, Y. Allelopathic Effect of Leaf Debris, Leaf Aqueous Extract and Rhizosphere Soil of Ophiopogon japonicus Ker-Gawler on the Growth of Plants. Weed Biol. Manag. 2004, 4, 43–48. [Google Scholar] [CrossRef]
- Kumar, V.; Brainard, D.C.; Bellinder, R.R. Suppression of Powell Amaranth (Amaranthus powellii) by Buckwheat Residues: Role of Allelopathy. Weed Sci. 2009, 57, 66–73. [Google Scholar] [CrossRef]
- Chandra, S.; Chatterjee, P.; Dey, P.; Bhattacharya, S. Allelopathic Effect of Ashwagandha against the Germination and Radicle Growth of Cicer Arietinum and Triticum Aestivum. Pharmacognosy Res. 2012, 4, 166. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Total Amount in Each Organs (mg g−1 DW) | Concentration of Compounds in Examined Organs of Rosemary (mg g−1 DW) | |||
|---|---|---|---|---|---|
| Leaves | Inflorescences | Stems | Roots | ||
| Caffeic acid | 0.45 ± 0.01 | 0.02 ± 0.01 | 0.01 ± 0.00 | ND | 0.48a |
| Ferulic acid | 0.81 ± 0.03 | 0.01 ± 0.00 | 0.02 ± 0.01 | 0.01 ± 0.00 | 0.85a |
| Gallic acid | 1.72 ± 0.31 | 0.02 ± 0.00 | 0.01 ± 0.00 | ND | 1.72a |
| Carnosic acid | 25.3 ± 3.21 | 12.5 ± 1.36 | 8.31 ± 0.96 | 5.32 ± 0.85 | 51.4b |
| Chlorogenic acid | 0.92 ± 0.12 | 0.01 ± 0.00 | ND | 0.02 ± 0.00 | 0.95a |
| Rosmarinic acid | 11.9 ± 1.21 | 4.52 ± 0.65 | 3.21 ± 0.31 | 3.51 ± 0.65 | 23.1c |
| Amount in Organ | 41.1b | 17.1a | 11.6a | 8.86a | - |
| Family | Common Name | Scientific Name | Elongation (% of Control) | Paper Coloration | |
|---|---|---|---|---|---|
| Radicle | Hypocotyl | ||||
| Amaranthaceae | Globe amaranth | Gomphrena globosa L. | 23.5 ± 2.05 | 45.3 ± 4.33 | No change |
| Common cockscomb | Celosia cristata L. | 26.4 ± 2.13 | 65.0 ± 10.3 | No change | |
| Apiaceae | Fennel | Foeniculum vulgare Mill. | 67.6 ± 3.15 | 83.8 ± 12.3 | Reddish |
| Carrot | Daucus carota L. | 74.5 ± 4.35 | 86.3 ± 13.5 | Reddish | |
| Asteraceae | Lettuce | Lactuca sativa L. | 6.0 ± 1.05 | 13.2 ± 0.89 | Grey |
| Crown daisy | Glebionis coronaria (L). Cass. Ex. Spach | 28.7 ± 2.19 | 32.7 ± 2.56 | No change | |
| Brassicaceae | Komatsuna | Brassica rapa var. perviridis L.H. Bailey | 10.0 ±1.56 | 24.8 ± 2.01 | Grey |
| Pak choi | Brassica rapa subsp. Chinensis (L.) Hanelt | 9.1 ± 1.15 | 37.9 ± 2.65 | Grey | |
| Radish | Raphanus raphanistrum subsp. Sativus (L.) Domin | 15.4 ± 1.86 | 28.4 ± 2.89 | Grey | |
| Turnip | Brassica rapa subsp. Rapa L. | 7.7 ± 0.11 | 31.9 ± 2.66 | No change | |
| Arugula | Eruca sativa Mill. | 7.3 ± 0.15 | 21.7 ± 1.95 | Grey | |
| Fabaceae | White clover | Trifolium repens L. | 25.6 ± 2.25 | 21.2 ± 1.80 | Reddish |
| Hairy vetch | Vicia villosa Roth | 28.9 ± 2.6 | 40.9 ± 3.10 | Reddish | |
| Alfalfa | Medicago sativa L. | 31.7 ± 2.65 | 34.7 ± 5.33 | Yellowish | |
| Lamiaceae | Basil | Ocimum basilicum L. | 26.9 ± 1.56 | 35.4 ± 4.22 | No change |
| Rosemary | Rosmarinus officinalis L. | 22.1 ± 1.31 | 28.3 ± 3.33 | No change | |
| Poaceae | Meadow fescue | Festuca pratensis Huds. | 41.2 ± 3.01 | 59.5 ± 7.56 | Reddish |
| Perennial ryegrass | Lolium perenne L. | 26.6 ± 2.66 | 38.9 ± 6.44 | Reddish | |
| Italian ryegrass | Lolium multiflorum Lam. | 6.2 ± 0.01 | 49.1 ± 7.56 | Reddish | |
| Kentucky bluegrass | Poa pratensis L. | 12.4 ± 1.23 | 37.8 ± 5.66 | Reddish | |
| Timothy grass | Phleum pratense L. | 3.8 ± 0.02 | 7.0 ± 0.89 | No change | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Appiah, K.S.; Mardani, H.K.; Omari, R.A.; Eziah, V.Y.; Ofosu-Anim, J.; Onwona-Agyeman, S.; Amoatey, C.A.; Kawada, K.; Katsura, K.; Oikawa, Y.; et al. Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves. Toxins 2018, 10, 498. https://doi.org/10.3390/toxins10120498
Appiah KS, Mardani HK, Omari RA, Eziah VY, Ofosu-Anim J, Onwona-Agyeman S, Amoatey CA, Kawada K, Katsura K, Oikawa Y, et al. Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves. Toxins. 2018; 10(12):498. https://doi.org/10.3390/toxins10120498
Chicago/Turabian StyleAppiah, Kwame Sarpong, Hossein Korrani Mardani, Richard Ansong Omari, Vincent Yao Eziah, John Ofosu-Anim, Siaw Onwona-Agyeman, Christiana Adukwei Amoatey, Kiyokazu Kawada, Keisuke Katsura, Yosei Oikawa, and et al. 2018. "Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves" Toxins 10, no. 12: 498. https://doi.org/10.3390/toxins10120498
APA StyleAppiah, K. S., Mardani, H. K., Omari, R. A., Eziah, V. Y., Ofosu-Anim, J., Onwona-Agyeman, S., Amoatey, C. A., Kawada, K., Katsura, K., Oikawa, Y., & Fujii, Y. (2018). Involvement of Carnosic Acid in the Phytotoxicity of Rosmarinus officinalis Leaves. Toxins, 10(12), 498. https://doi.org/10.3390/toxins10120498