Toxicity of Ochratoxin to Early Life Stages of Zebrafish (Danio rerio)


Abstract
:1. Introduction
2. Results
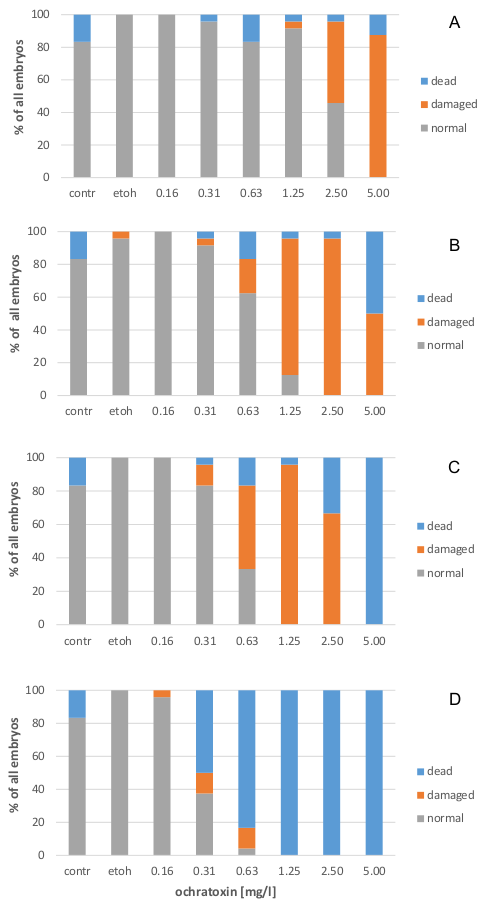
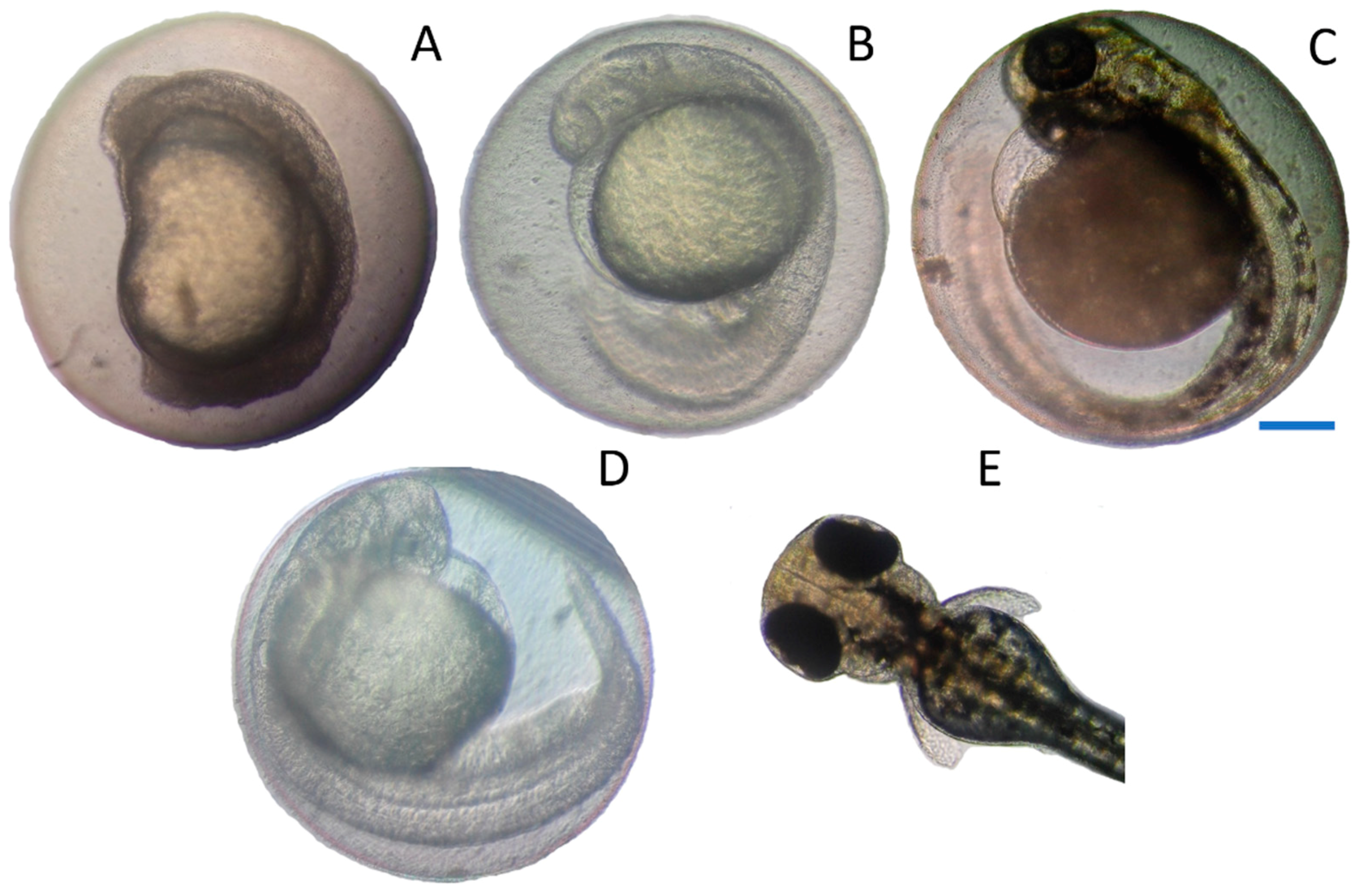
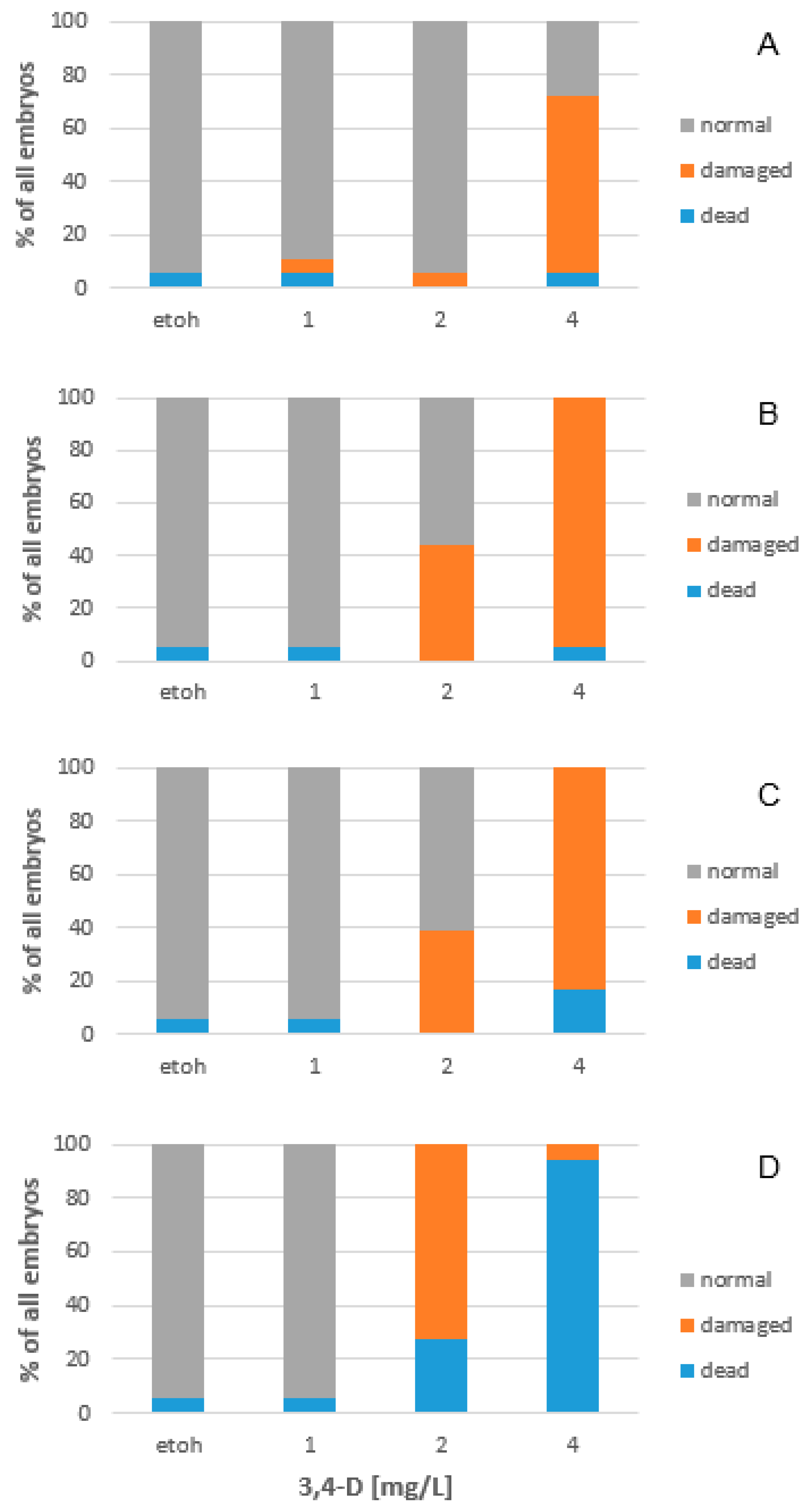
2.1. Effects on Zebrafish Embryo Development
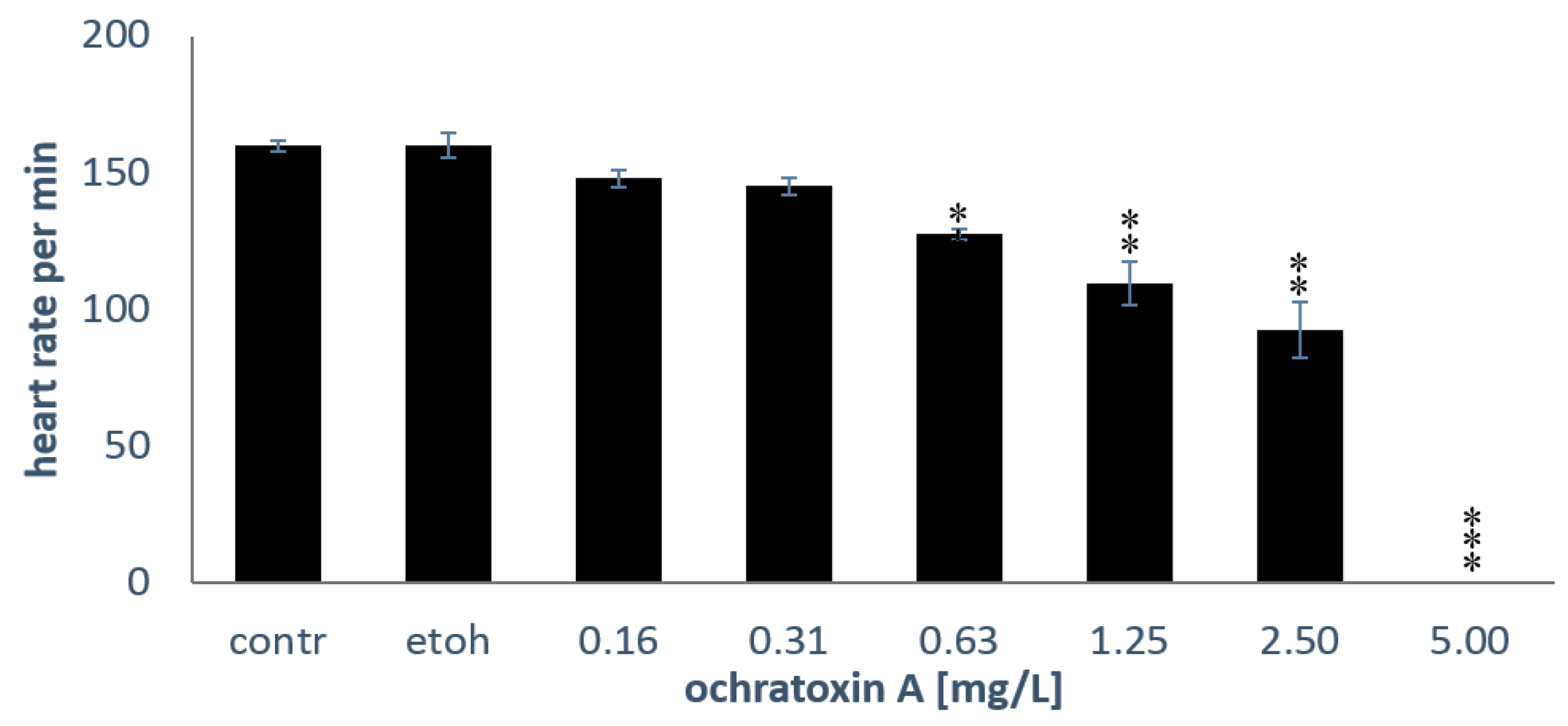
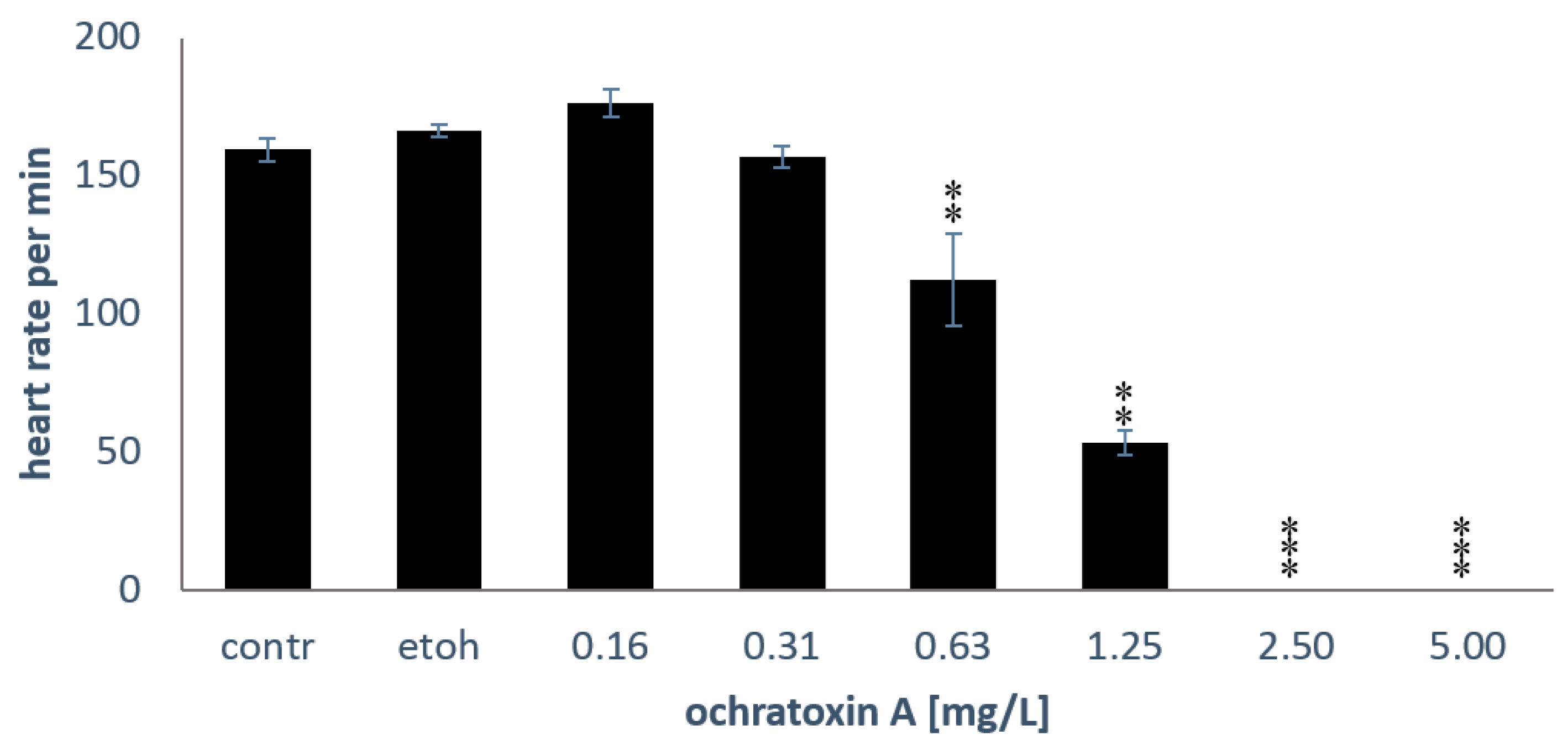
2.2. Heart Rates
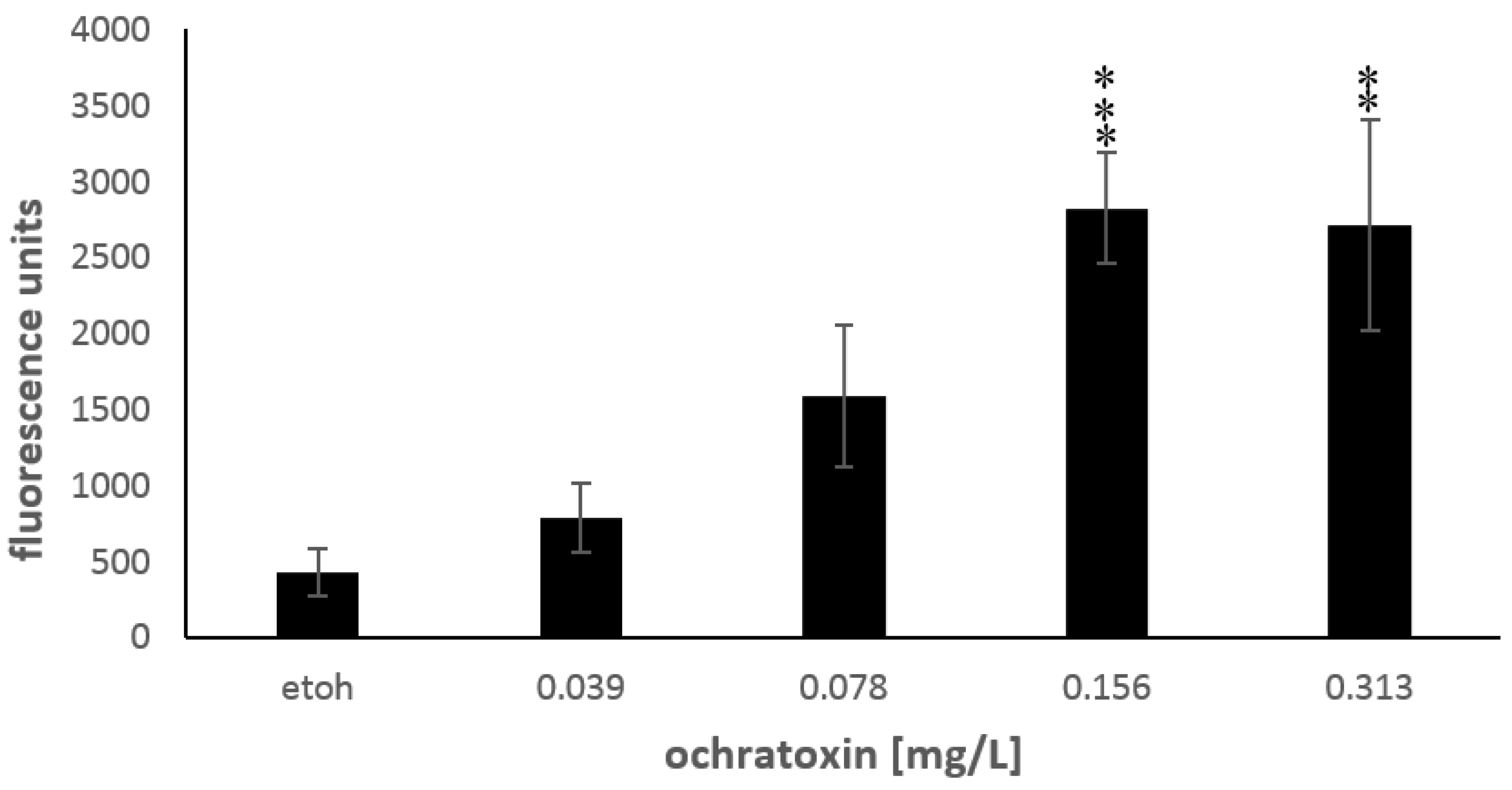
2.3. Oxidative Stress
2.4. 3,4-Dichloroaniline
3. Discussion
3.1. Toxicity of OTA to Fish Embryos
3.2. Detection of Reactive Oxygen Species (ROS)
3.3. Positive Controls
4. Conclusions
5. Materials and Methods
5.1. Chemicals
5.2. Preparation of Exposure Medium
5.3. Exposure of Fish Embryos
5.4. Assessment of Development
5.5. Measurement of Reactive Oxygen Species (ROS)
5.6. Calculation of the 50% Level for Effects and Lethality
5.7. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Microbiol. 2010, 27, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Naehrer, K. Prevalence of mycotoxins in feedstuffs and feed surveyed worldwide in 2009 and 2010. Phytopathol. Mediterr. 2012, 51, 175–192. [Google Scholar]
- Moss, M.O. Risk assessment for aflatoxins in foodstuffs. Int. Biodeterior. Biodegrad. 2002, 50, 137–142. [Google Scholar] [CrossRef]
- Manning, B.B.; Ulloa, R.M.; Li, M.H.; Robinson, E.H.; Rottinghaus, G.E. Ochratoxin A fed to channel catfish (Ictalurus punctatus) causes reduced growth and lesions of hepatopancreatic tissue. Aquaculture 2003, 219, 739–750. [Google Scholar] [CrossRef]
- Reddy, L.; Bhoola, K. Ochratoxins—Food Contaminants: Impact on Human Health. Toxins 2010, 2, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlyssides, A.G.; Barampouti, E.M.; Mai, S. Wastewater characteristics from Greek wineries and distilleries. Water Sci. Technol. 2005, 51, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, R.; Estevinho, I.; Abrunhosa, L.; Mendonc, C.; Machado, P.; Carballa, M.; Brito, A.G.; Venancio, A. Assessing the degradation of ochratoxin a using a bioassay: The case of contaminated winery wastewater. Water Sci. Technol. 2007, 56, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duarte, S.C.; Lino, C.M.; Pena, A. Ochratoxin A in feed of food-producing animals: An undesirable mycotoxin with health and performance effects. Vet. Microbiol. 2011, 154, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boorman, G.A. NTP Technical Report on the Toxicology and Carcinogenic Studies of Ochratoxin A (CAS No. 303-47-9) in F344/n Rats (Gavage Studies); NIH Publication No. 89-2813; Department of Health and Human Services, National Institute of Health: Research Triangle Park, NC, USA, 1989.
- Brown, M.H.; Szczech, G.M.; Purmalis, B.P. Teratogenic and toxic effects of ochratoxin A in rats. Toxicol. Appl. Pharmacol. 1976, 37, 331–338. [Google Scholar] [CrossRef]
- Arora, R.G.; Froelein, H.; Fellner-Feldegg, H. Inhibition of ochratoxin A teratogenesis by zearalenone and diethylstilbestrol. Food Chem. Toxicol. 1983, 21, 779–783. [Google Scholar] [CrossRef]
- Wiger, R.; Stormer, F.C. Effects of ochratoxins A and B on prechondrogenic mesenchymal cells from chick embryo limb buds. Toxicol. Lett. 1990, 54, 129–134. [Google Scholar] [CrossRef]
- Hood, R.D.; Naughton, M.J.; Hayes, A.W. Prenatal effects of ochratoxin A in hamsters. Teratology 2005, 13, 11–14. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, E.; Prietz, A.; Dietrich, D.R. Investigation of the teratogenic potential of ochratoxin A and B using the FETAX system. Birth Defects Res. B Dev. Reprod. Toxicol. 2005, 74, 417–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haq, M.; Gonzalez, N.; Mintz, K.; Jaja-Chimedza, A.; De Jesus, C.L.; Lydon, C.; Welch, A.Z.; Berry, J.P. Teratogenicity of ochratoxin A and the degradation product, ochratoxin α, in the zebrafish (Danio rerio) embryo model of vertebrate development. Toxins 2016, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Doster, R.C.; Sinnhuber, R.O.; Pawlowski, N.E. Acute intraperitoneal toxicity of ochratoxin A and B derivatives in rainbow trout (Salmo gairdneri). Food Cosmet. Toxicol. 1974, 12, 499–505. [Google Scholar] [CrossRef]
- El-Sayed, Y.S.; Khalil, R.H.; Saad, T.T. Acute toxicity of ochratoxin-A in marine water-reared sea bass (Dicentrarchus labrax L.). Chemosphere 2009, 75, 878–882. [Google Scholar] [CrossRef] [PubMed]
- Bernhoft, A.; Høgåsen, H.R.; Rosenlund, G.; Ivanova, L.; Berntssen, M.H.G.; Alexander, J.; Sundstøl Eriksen, G.; Kruse Fæste, C. Tissue distribution and elimination of deoxynivalenol and ochratoxin A in dietary exposed Atlantic salmon (Salmo salar). Food Addit. Contam. Part A 2017, 34, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Lovell, R.T. Mycotoxins: Hazardous to Farmed Fish. Feed Int. 1992, 13, 24–28. [Google Scholar]
- Zahran, E.; Manning, B.; Seo, J.-K.; Noga, E.J. The effect of Ochratoxin A on antimicrobial polypeptide expression and resistance to water mold infection in channel catfish (Ictalurus punctatus). Fish Shellfish Immunol. 2016, 57, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Srour, T.M. Effect of ochratoxin-A with or without Biogen® on growth performance, feed utilization and carcass composition of Nile tilapia (Oreochromis niloticus) fingerlings. J. Agric. Sci. Mansoura Univ. 2004, 29, 51–61. [Google Scholar]
- Supamattaya, K.; Sukrakanchana, N.; Boonyaratpalin, M.; Schatzmayr, D.; Chittiwan, V. Effects of ochratoxin A and deoxynivalenol on growth performance and immuno-physiological parameters in black tiger shrimp (Penaeus monodon). Songklanakarin J. Sci. Technol. 2005, 27 (Suppl. 1), S91–S99. [Google Scholar]
- Hauptman, B.S.; Barrows, F.T.; Block, S.S.; Gaylord, T.G.; Paterson, J.A.; Sealey, W.M. Potential for a mycotoxin deactivator to improve growth and performance of rainbow trout fed high levels of an ethanol industry co-product, grain distiller’s dried yeast. N. Am. J. Aquac. 2014, 76, 297–304. [Google Scholar] [CrossRef]
- Braunbeck, T.; Lammer, E. Background Paper on Fish Embryo Toxicity Assays; German Federal Environment Agency: Dessau-Roßlau, Germany, 2006; p. 298. [Google Scholar]
- Kuiper-Goodman, T.; Scott, P.M. Risk assessment of the mycotoxin ochratoxin A. Biomed. Environ. Sci. 1989, 2, 179–248. [Google Scholar] [PubMed]
- Peckham, J.; Doupnik, B.J.; Jones, O.J. Acute toxicity of ochratoxins A and B in chicks. Appl. Microbiol. 1971, 21, 492–494. [Google Scholar] [PubMed]
- Heussner, A.H.; Bingle, L.E.H. Comparative ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4282. [Google Scholar] [CrossRef] [PubMed]
- Okutan, H.; Aydin, G.; Ozcelik, N. Protective role of melatonin in ochratoxin A toxicity in rat heart and lung. J. Appl. Toxicol. 2004, 24, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Jameel, F.A. Pathological effects of ochratoxin A in brain, heart and lung of chicks. Al-Anbar J. Vet. Sci. 2011, 4, 93–98. [Google Scholar]
- Van Egmond, H.P.; Jonker, M.A. Worldwide Regulations for Mycotoxins in Food and Feed in 2003; Food and Nutrition Paper No. 81; Food and Agriculture Organization (FAO): Rome, Italy, 2004. [Google Scholar]
- Rafai, P.; Bata, A.; Jakab, L.; Vanyi, A. Evaluation of mycotoxin contaminated cereals for their use in animal feeds in Hungary. Food Addit. Contam. 2000, 17, 799–808. [Google Scholar] [CrossRef] [PubMed]
- Czerwiecki, L.; Czajkowska, D.; Witkowska-Gwiazdowska, A. On ochratoxin A and fungal flora in polish cereals from conventional and ecological farms. Part 2: Occurrence of ochratoxin A and fungi in cereals in 1998. Food Addit. Contam. 2002, 19, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Schaaf, G.J.; Nijmeijer, S.M.; Maas, R.F.M.; Roestenberg, P.; de Groene, E.M.; Fink-Gremmels, J. The role of oxidative stress in the ochratoxin A-mediated toxicity in proximal tubular cells. Biochim. Biophys. Acta 2002, 1588, 149–158. [Google Scholar] [CrossRef]
- Sorrenti, V.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Bognanno, M.; Galvano, F. Toxicity of ochratoxin A and its modulation by antioxidants: A review. Toxins 2013, 5, 1742–1766. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Xie, S.; Xu, F.; Liu, A.; Wang, Y.; Chen, D.; Pan, Y.; Huang, L.; Peng, D.; Wang, X.; Yuan, Z. Ochratoxin A: Toxicity, oxidative stress and metabolism. Food Chem. Toxicol. 2018, 112, 320–331. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Yu, T.; Qi, X.; Gao, J.; Huang, K.; He, X.; Luo, H.; Xu, W. Limited link between oxidative stress and ochratoxin A—Induced renal injury in an acute toxicity rat model. Toxins 2016, 8, 373. [Google Scholar] [CrossRef] [PubMed]
- Kamp, H.G.; Eisenbrand, G.; Schlatter, J.; Wurth, K.; Janzowski, C. Ochratoxin A: Induction of (oxidative) DNA damage, cytotoxicity and apoptosis in mammalian cell lines and primary cells. Toxicology 2005, 31, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Longoria, A.; Tinoco, R.; Vazquez-Duhalt, R. Chloroperoxidase-mediated transformation of highly halogenated monoaromatic compounds. Chemosphere 2008, 7, 485–490. [Google Scholar] [CrossRef] [PubMed]
- El Adlouni, C.; Pinelli, E.; Azemar, B.; Zaoui, D.; Beaune, P.; Pfohl-Leszkowicz, A. Phenobarbital increases DNA adduct and metabolites formed by ochratoxin A: Role of CYP 2C9 and microsomal glutathione-S-transferase. Environ. Mol. Mutagen. 2000, 35, 123–131. [Google Scholar] [CrossRef]
- EURL ECVAM (European Commission European Union Reference Laboratory for Alternatives to Animal Testing). Recommendation on the Zebrafish Embryo Acute Toxicity Test Method (ZFET) for Acute Fish Testing Toxicity; EUR 26710—Joint Research Centre—Institute for Health and Consumer Protection; EUR—Scientific and Technical Research Series; Publications Office of the European Union: Luxembourg, 2014; p. 34. [Google Scholar]
- Nagel, R.; Bresch, H.; Caspers, N.; Hansen, P.D.; Markert, M.; Munk, R.; Scholz, N.; ter Höfte, B.B. Effect of 3,4-dichloroaniline on the early life stages of the zebrafish (Brachydanio rerio): Results of a comparative laboratory study. Ecotoxicol. Environ. Saf. 1991, 21, 157–164. [Google Scholar] [CrossRef]
- Zhu, B.; Liu, T.; Hu, X.; Wang, G. Developmental toxicity of 3,4-dichloroaniline on rare minnow (Gobiocypris rarus) embryos and larvae. Chemosphere 2013, 90, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- DIN Norm 38415-6 T6. German Standard Methods for the Examination of Water, Waste Water and Sludge—Subanimal Testing (Group T)—Part 6: Toxicity to Fish; Determination of the Non-Acute-Poisonous Effect of Waste Water to Fish Eggs by Dilution Limits (T 6); Deutsches Institut für Normung e.V.: Berlin, Germany, 2001. [Google Scholar]
- Gauron, C.; Rampon, C.; Bouzaffour, M.; Ipendey, E.; Teillon, J.; Volovitch, M.; Vriz, S. Sustained production of ROS triggers compensatory proliferation and is required for regeneration to proceed. Sci. Rep. 2013, 3, 2084. [Google Scholar] [CrossRef] [PubMed]
- Pietsch, C.; Noser, J.; Wettstein, F.E.; Burkhardt-Holm, P. Unraveling the mechanisms involved in zearalenone-mediated toxicity in permanent fish cell cultures. Toxicon 2014, 88, 44–61. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Point | EC50 (mg/L) | LC50 (mg/L) | Ratio LC50 to EC50 |
|---|---|---|---|
| 24 h | 2.65 | 24.22 | 9.14 |
| 48 h | 0.73 | 2.57 | 3.52 |
| 72 h | 0.56 | 3.32 | 5.93 |
| 96 h | 0.29 | 0.36 | 1.24 |
| Treatment | 72 h | 96 h |
|---|---|---|
| contr | 83.3% (100%) | 83.3% (100%) |
| etoh | 95.8% (95.8%) | 100% (100%) |
| 0.16 mg/L OTA | 95.8% (95.8%) | 100% (100%) |
| 0.31 mg/L OTA | 79.2% (82.6%) | 37.5% (81.8%) |
| 0.63 mg/L OTA | 16.7% (21.1%) | 4.2% (33.3%) |
| 1.25 mg/L OTA | 0% (0%) | 0% (0%) |
| 2.50 mg/L OTA | 0% (0%) | 0% (0%) |
| 5.00 mg/L OTA | 0% (0%) | 0% (0%) |
| Time Point | EC50 (mg/L) | LC50 (mg/L) | Ratio LC50 to EC50 |
|---|---|---|---|
| 24 h | 3.47 | 9.06 | 2.61 |
| 48 h | 2.09 | 9.06 | 4.59 |
| 72 h | 2.22 | 6.08 | 2.74 |
| 96 h | 1.59 | 2.88 | 1.81 |
| Time Point | Endpoint |
|---|---|
| 24 h | eye visible, somites, tail movements |
| 48 h | pigmentation, blood circulation, heartbeats, oedema, embryo movements |
| 72 h | pigmentation, blood circulation, heartbeats, oedema, embryo movements, hatching |
| 96 h | pigmentation, blood circulation, heartbeats, oedema, embryo movements, hatching |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tschirren, L.; Siebenmann, S.; Pietsch, C. Toxicity of Ochratoxin to Early Life Stages of Zebrafish (Danio rerio). Toxins 2018, 10, 264. https://doi.org/10.3390/toxins10070264
Tschirren L, Siebenmann S, Pietsch C. Toxicity of Ochratoxin to Early Life Stages of Zebrafish (Danio rerio). Toxins. 2018; 10(7):264. https://doi.org/10.3390/toxins10070264
Chicago/Turabian StyleTschirren, Linda, Seraina Siebenmann, and Constanze Pietsch. 2018. "Toxicity of Ochratoxin to Early Life Stages of Zebrafish (Danio rerio)" Toxins 10, no. 7: 264. https://doi.org/10.3390/toxins10070264
APA StyleTschirren, L., Siebenmann, S., & Pietsch, C. (2018). Toxicity of Ochratoxin to Early Life Stages of Zebrafish (Danio rerio). Toxins, 10(7), 264. https://doi.org/10.3390/toxins10070264