Low Doses of Ochratoxin-A Decrease IgY and IgA Production in Broiler Chicks

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
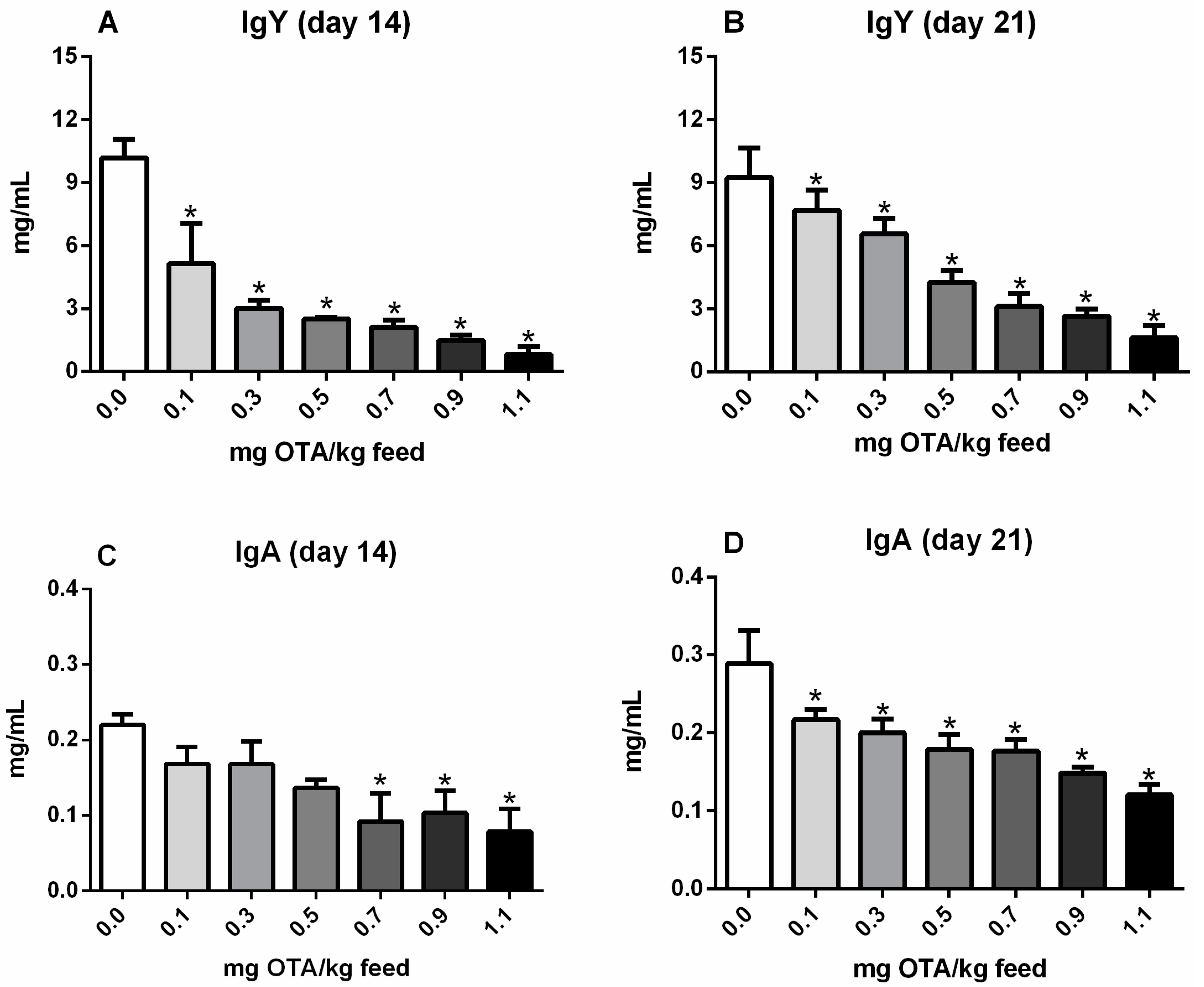
2.1. Serum Levels of Total IgY and IgA in Chicks
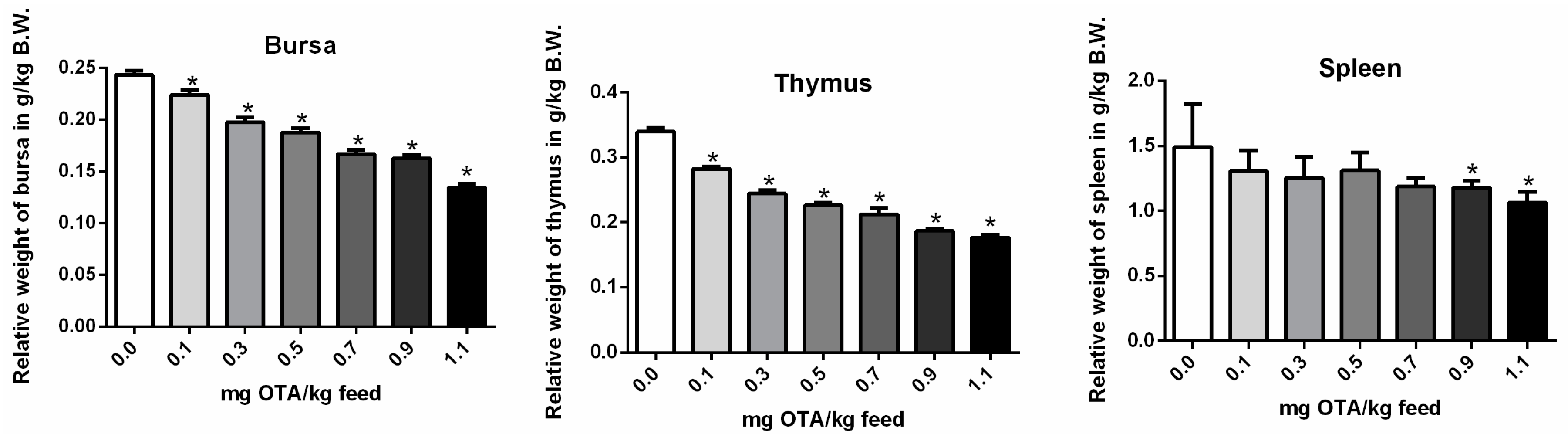
2.2. Relative Weights of the Bursa, Thymus, and Spleen
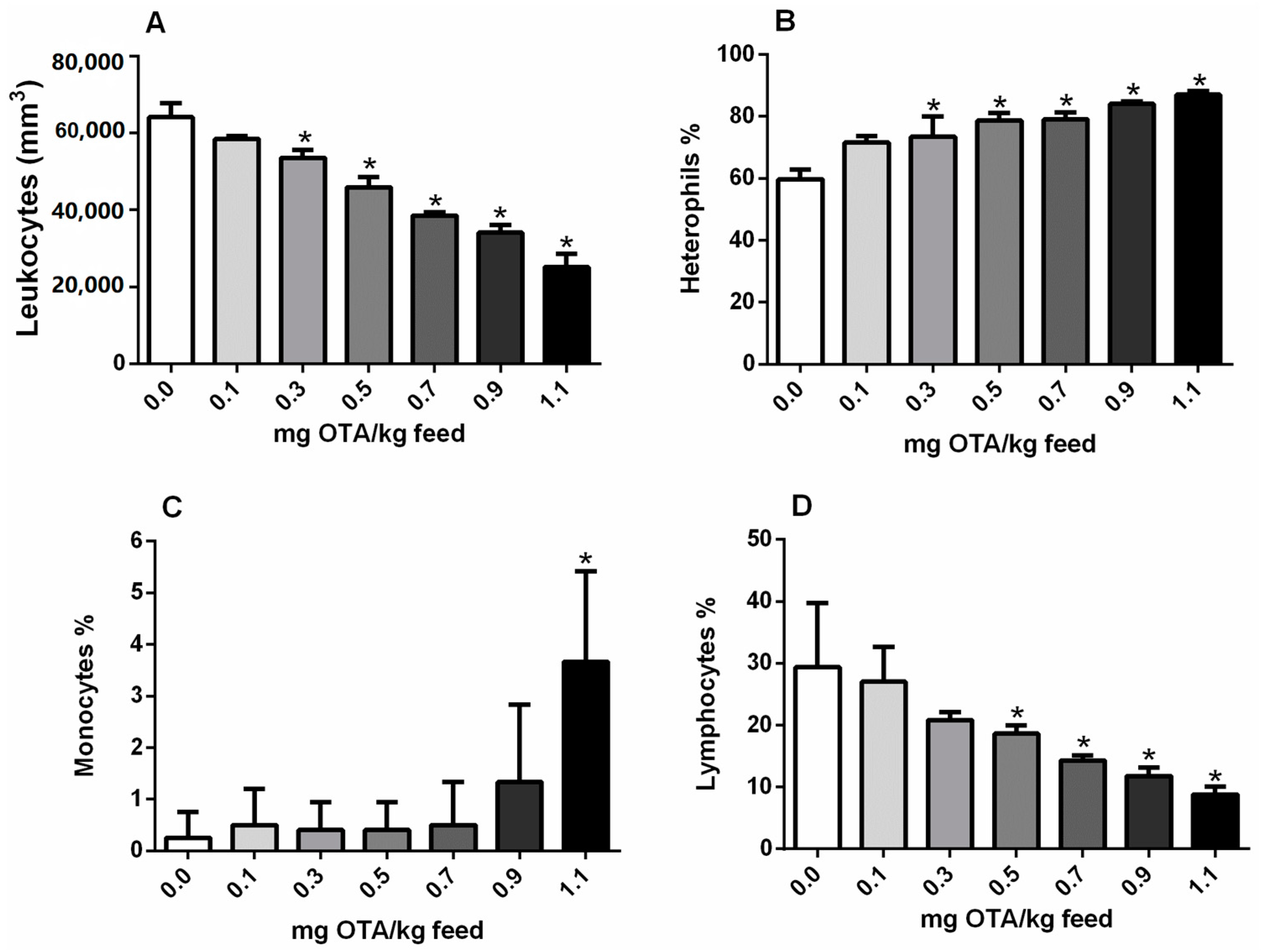
2.3. Circulating Leukocytes and Differential Counts
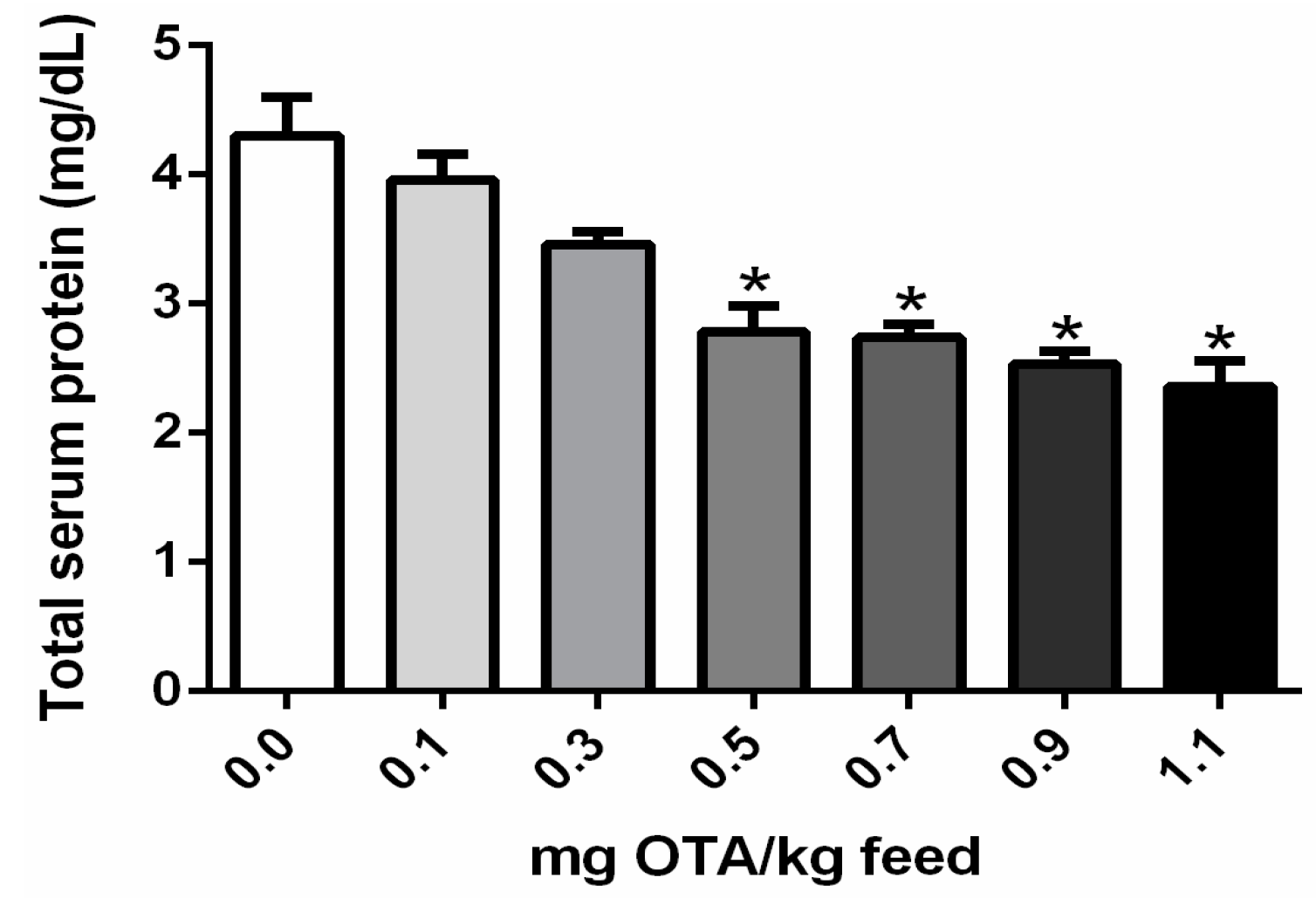
2.4. Total Serum Protein Concentrations in Chicks Exposed to OTA
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Experimental Chicks and Their Management
5.2. Preparation of OTA-Contaminated Feed
5.3. Experimental Design
5.4. Quantification of IgY and IgA
5.5. Relative Weights of the Bursa, Thymus, and Spleen
5.6. Circulating Leukocyte Profiles and Differential Counts
5.7. Total Serum Protein Concentrations in Chicks Exposed to OTA
5.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Trenk, H.L.; Butz, M.E.; Chu, F.S. Production of ochratoxins in different cereal products by Aspergillus ochraceous. Appl. Microbiol. 1971, 21, 1032–1035. [Google Scholar] [PubMed]
- Malir, F.; Ostry, V.; Pfohl-Leszkowicz, A.; Malir, J.; Toman, J. Ochratoxin A: 50 years of research. Toxins 2016, 8, 191. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L. Mycotoxin contamination of the feed supply chain: Implications for animal productivity and feed security. Anim. Feed Sci. Technol. 2012, 173, 134–158. [Google Scholar] [CrossRef]
- Binder, E.M.; Tan, L.M.; Chin, J.L.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Technol. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Magnoli, A.A.; Barberis, C.; Chiacchiera, S.M.; Combina, M.; Dalcero, A. Ochratoxin A production in relation to ecophysiological factors by Aspergillus section Nigri strains isolated from different substrates in Argentina. Sci. Total Environ. 2007, 388, 16–23. [Google Scholar]
- Duarte, S.C.; Lino, C.M.; Pena, A. Ochratoxin A in feed of food-producing animals: An undesirable mycotoxin with health and performance effects. Vet. Microbiol. 2011, 154, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, I.; Naehrer, K. A three-year survey on the worldwide occurrence of mycotoxins in feedstuffs and feed. Toxins 2012, 4, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Streit, E.; Naehrer, K.; Rodrigues, I.; Schatzmayra, G. Mycotoxin occurrence in feed and feed raw materials worldwide: Long-term analysis with special focus on Europe and Asia. J. Sci. Food Agric. 2013, 93, 2892–2899. [Google Scholar] [CrossRef] [PubMed]
- Sherazi, S.T.H.; Shar, Z.H.; Sumbal, G.A.; Eddie, T.; Tan, E.T.; Bhanger, M.I.; Kara, H.; Nizamani, S.M. Occurrence of ochratoxin A in poultry feeds and feed ingredients from Pakistan. Mycotoxin Res. 2015, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhihong, L.; Kunlun, H.; Yunbo, L. Ochratoxin A and ochratoxin-producing fungi on cereal grain in China: A review. Food Add. Cont. Part A 2015, 32, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Boudra, H.; Bars, L.P.; Bars, L.J. Thermostability of Ochratoxin A in wheat under two moisture conditions. Appl. Environ. Microbiol. 1995, 61, 1156–1158. [Google Scholar] [PubMed]
- Reddy, L.; Bhoola, K. Ochratoxins—Food contaminants: Impact on human health. Toxins 2010, 2, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Denli, M.; Perez, J.F. Ochratoxins in feed, a risk for animal and human health: Control strategies. Toxins 2010, 2, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Rigobello, F.F.; e Leonello-Álvares Silva, P.; Yamashita, C.R.T.; Lenhard-Vidal, A.; Ishikawa, A.T.; Kawamura, O.; Hirooka, E.Y.; Itano, E.N. Ochratoxin A levels in plasma from inhabitants of northern Paraná, Brazil. World Mycotoxin J. 2016, 9, 623–632. [Google Scholar] [CrossRef]
- Milićević, D.; Jovanović, M.; Matekalo-Sverak, V.; Radicević, T.; Petrović, M.M.; Lilić, S. A survey of spontaneous occurrence of ochratoxin A residues in chicken tissues and concurrence with histopathological changes in liver and kidneys. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2011, 29, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Ostry, W.; Malir, F.; Toman, J.; Grosse, Y. Mycotoxins as human carcinogens—The IARC Monographs classification. Mycotoxin Res. 2017, 33, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Heussner, A.; Bingle, L.E.H. Comparative ochratoxin toxicity: A review of the available data. Toxins 2015, 7, 4253–4282. [Google Scholar] [CrossRef] [PubMed]
- Sava, V.; Reunova, O.; Velasquez, A.; Harbison, R.; Sanchez-Ramos, J. Acute neurotoxic effects of the fungal metabolite ochratoxin-A. Neurotoxicology 2006, 27, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, V.; Di Giacomo, C.; Acquaviva, R.; Barbagallo, I.; Bognanno, M.; Galvano, F. Toxicity of ochratoxin A and its modulation by antioxidants: A review. Toxins 2013, 5, 1742–1766. [Google Scholar] [CrossRef] [PubMed]
- Anzai, N.; Jutabha, P.; Endou, H. Molecular mechanism of ochratoxin A transport in the kidney. Toxins 2010, 2, 1381–1398. [Google Scholar] [CrossRef] [PubMed]
- Girish, C.K.; Smith, T.K. Impact of feed-borne mycotoxins on avian cell-mediated and humoral immune responses. World Mycotoxin J. 2008, 1, 105–121. [Google Scholar] [CrossRef]
- Anati, M.A.; Petzinger, E. Immunotoxic activity of ochratoxin A. J. Vet. Pharmacol. Ther. 2006, 29, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Elaroussi, M.A.; Mohamed, F.R.; El Barkouky, E.M.; Atta, A.M.; Abdou, A.M.; Hatab, M.H. Experimental ochratoxicosis in broiler chickens. Avian Pathol. 2006, 35, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, F.; Magor, K.E.; Kaspers, B. Structure and evolution of avian immunoglobulins. In Avian Immunology, 1st ed.; Davison, F., Kaspers, B., Schat, K.A., Eds.; Elsevier: San Diego, CA, USA, 2008; pp. 107–127. ISBN 978-0-12-370634-8. [Google Scholar]
- Singh, G.S.; Chauhan, H.V.; Jha, G.J.; Singh, K.K. Immunosuppression due to chronic ochratoxicosis in broiler chicks. J. Comp. Pathol. 1990, 103, 399–410. [Google Scholar] [CrossRef]
- Santin, E.; Paulillo, A.C.; Maiorka, P.C.; Alessi, A.C.; Krabbe, E.L.; Maiorka, A. The effects of ochratoxin/aluminosilicate interaction on the tissues and humoral immune response of broilers. Avian Pathol. 2002, 31, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, J.; Johri, T.; Swain, B.; Ameena, S. Effect of graded levels of aflatoxin, ochratoxin and their combinations on the performance and immune response of broilers. Br. Poult. Sci. 2004, 45, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Indresh, H.C.; Umakantha, B. Effects of ochratoxin and T-2 toxin combination on performance, biochemical and immune. status of commercial broilers. Vet. World 2013, 6, 945–949. [Google Scholar] [CrossRef]
- Dwivedi, P.; Burns, R.B. Effect of ochratoxin A on immunoglobulins in broiler chicks. Res. Vet. Sci. 1984, 36, 117–121. [Google Scholar] [PubMed]
- Pozzo, L.; Salamano, G.; Mellia, E.; Gennero, M.S.; Doglione, L.; Cavallarin, L.; Tarantola, M.; Forneris, G.; Schiavone, A. Feeding a diet contaminated with ochratoxin A for chickens at the maximum level recommended by the EU for poultry feeds (0.1 mg/kg). 1. Effects on growth and slaughter performance, haematological and serum traits. J. Anim. Physiol. Anim. Nutr. 2013, 1, 13–22. [Google Scholar] [CrossRef] [PubMed]
- European Commission: Commission Recommendation of 17 August 2006 on the presence of deoxynivalenol, zearalenone, Ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding, 2006/576/EC. Off. J. Eur. Communities 2006, L 229, 7–9.
- Orr-Burks, N.; Gulley, S.L.; Toro, H.; van Ginkel, F.W. Immunoglobulin A as an early humoral responder after mucosal avian coronavirus vaccination. Avian Dis. 2014, 58, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Stoev, S.; Anguelov, G.; Ivanov, I.; Pavlov, D. Influence of ochratoxin A and an extract of artichoke on the vaccinal immunity and health in broiler chicks. Exp. Toxicol. Pathol. 2000, 52, 43–55. [Google Scholar] [CrossRef]
- Nakamura, K.; Imada, Y.; Abe, F. Effect of cyclophosphamide on infections produced by Escherichia coli of high and low virulence in chickens. Avian Pathol. 1987, 16, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.F.; Huff, W.E.; Hamilton, P.B.A. Leucocytopenia induced in chickens by dietary ochratoxin A. Poult. Sci. 1979, 58, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, S.M.; Warasi, S.M.A.; Vikram Reddy, M. Haematological and biochemical changes in experimental ochratoxicosis in broiler chicken. Indian Vet. J. 1993, 70, 613–617. [Google Scholar]
- Chang, C.F.; Hamilton, P.B. Impairment of phagocytosis by heterophils from chickens during ochratoxicosis. Appl. Environ. Microbiol. 1980, 39, 572–575. [Google Scholar] [PubMed]
- Moura, M.; Machado, C.; Porfirio, L.; Freire, R. Effects of ochratoxin-A on broiler leukocytes. Revista Brasileira de Ciência Avícola 2004, 6, 187–190. [Google Scholar] [CrossRef]
- Bartlomiej, J.; Pliszczak-Król, A.; Graczyk, S.; Kamal, R. Morphological and functional evaluation of chicken blood leukocytes in chronic ochratoxicosis. Int. J. Poult. Sci. 2006, 5, 191–194. [Google Scholar] [CrossRef]
- K˝oszegi, T.; Poór, M. Ochratoxin A: Molecular interactions, mechanisms of toxicity and prevention at the molecular level. Toxins 2016, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Lautert, C.; Ferreiro, L.; Azevedo, M.I.; Botton, S.A.; Santos, J.T.; Gonçalves, P.B.D.; Schlemmer, K.B.; Machado, V.S.; Santurio, J.M. In vitro effect of ochratoxin A and deoxynivalenol on the expression of interleukin 5 and interferon-gamma in broiler chicken lymphocytes. World Mycotoxin J. 2015, 9, 299–304. [Google Scholar] [CrossRef]
- Rostagno, H.S.; Albino, L.F.T.; Donzele, J.L.; Gomes, P.C.; Oliveira, R.F.; Lopes, D.C.; Ferreira, A.S.; Barreto, S.L.T.; Euclides, R.F. Tabelas Brasileiras para aves e suínos: Composição de alimentos e exigências nutricionais. In Brazilian Tables for Poultry and Swine: Composition of Feedstuffs and Nutritional Requirements, 3rd ed.; UFV: Viçosa, Brazil, 2011; pp. 1–252. [Google Scholar]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.A.; Venancio, E.J.; Fernandes, E.V.; Hirooka, E.Y.; Oba, A.; Flaiban, K.K.M.C.; Itano, E.N. Low Doses of Ochratoxin-A Decrease IgY and IgA Production in Broiler Chicks. Toxins 2018, 10, 316. https://doi.org/10.3390/toxins10080316
Khan SA, Venancio EJ, Fernandes EV, Hirooka EY, Oba A, Flaiban KKMC, Itano EN. Low Doses of Ochratoxin-A Decrease IgY and IgA Production in Broiler Chicks. Toxins. 2018; 10(8):316. https://doi.org/10.3390/toxins10080316
Chicago/Turabian StyleKhan, Shahzad A., Emerson J. Venancio, Eduardo V. Fernandes, Elisa Y. Hirooka, Alexandre Oba, Karina K. M. C. Flaiban, and Eiko N. Itano. 2018. "Low Doses of Ochratoxin-A Decrease IgY and IgA Production in Broiler Chicks" Toxins 10, no. 8: 316. https://doi.org/10.3390/toxins10080316
APA StyleKhan, S. A., Venancio, E. J., Fernandes, E. V., Hirooka, E. Y., Oba, A., Flaiban, K. K. M. C., & Itano, E. N. (2018). Low Doses of Ochratoxin-A Decrease IgY and IgA Production in Broiler Chicks. Toxins, 10(8), 316. https://doi.org/10.3390/toxins10080316