Abstract
Profundiconus is the most divergent cone snail genus and its unique phylogenetic position, sister to the rest of the family Conidae, makes it a key taxon for examining venom evolution and diversity. Venom gland and foot transcriptomes of Profundiconus cf. vaubani and Profundiconus neocaledonicus were de novo assembled, annotated, and analyzed for differential expression. One hundred and thirty-seven venom components were identified from P. cf. vaubani and 82 from P. neocaledonicus, with only four shared by both species. The majority of the transcript diversity was composed of putative peptides, including conotoxins, profunditoxins, turripeptides, insulin, and prohormone-4. However, there were also a significant percentage of other putative venom components such as chymotrypsin and L-rhamnose-binding lectin. The large majority of conotoxins appeared to be from new gene superfamilies, three of which are highly different from previously reported venom peptide toxins. Their low conotoxin diversity and the type of insulin found suggested that these species, for which no ecological information are available, have a worm or molluscan diet associated with a narrow dietary breadth. Our results indicate that Profundiconus venom is highly distinct from that of other cone snails, and therefore important for examining venom evolution in the Conidae family.
Key Contribution:
Reported here is the first characterization of the venom arsenal from Profundiconus, the most divergent cone snail genus. We identified several new Conus venom gene superfamilies, and insulin transcripts that suggest a worm or molluscan diet for the Profundiconus species examined.
1. Introduction
As stated by the “father” of toxicology, “Omnia venenum sunt: nec sine veneno quicquam existit. Dosis sola facit, ut venenum non fit.”: Everything can be a venom, only the dose makes the difference between a poisoning and a non-poisoning substance [1]. This intuition deftly summarized what happened independently several times across the animal kingdom, where gene families, encoding for proteins normally involved in key regulatory processes, were recruited for venom production [2]. Many of such gene families underwent positive selection and rapid expansion during the evolutionary history of venomous animal, showing the importance of defensive and predatory venoms in the prey-predator arms race [3]. Typically, venom is a complex mixture of different components generally referred to as ‘toxins’, which directly interfere with the physiology of the prey acting often as neurotoxins or enzymes impairing haemostasis [2,3]. However, they can be also listed as venom components those enzymes involved in post-translational modifications, folding processes, and in enhancing venom activity by easing toxins spreading [2,3].
Current studies of venomous animals allow for better understanding of convergent recruitment of venom component in distantly related taxa, prey–predator coevolution, and represent a starting point for the identification of new bioactive compounds of pharmaceutical interest. Among taxa that have developed a venomous function, the globally distributed gastropod superfamily Conoidea is a highly diversified group of carnivorous species including the family Conidae J. Fleming, 1822 [4,5]. Based on the latest published molecular phylogeny of the Conidae [6] six genera, comprising ~850 species, can be recognized: Profundiconus Kuroda, 1956, Californiconus J. K. Tucker & Tenorio, 2009, Lilliconus G. Raybaudi Massilia, 1994, Pygmaeconus Puillandre & Tenorio, 2017, Conasprella Thiele, 1929, and Conus Linnaeus, 1758. The Conidae venom arsenal, combined with sophisticated envenomation strategies, has allowed these slow-moving snail species to prey on worms, other molluscs, and fish [7]. Of the six Conidae genera, the complex venom cocktails of the genus Conus Linnaeus, 1758 (Conidae) have been intensively studied and contain mainly conotoxins that target ion channels, neurotransmitter transporters, and receptors of the nervous system of the prey [7]. A conotoxin precursor is typically composed of a conserved signal region, a pro-region that may be present or lacking, a highly variable mature peptide in most cases rich in cysteine residues, and a short terminal region [8]. The conotoxins are classified into gene superfamilies based on the percentage sequence identity (PID) of the signal region, and into cysteine patterns based on the number and distribution of cysteine residues in the mature region [8,9]. Data produced to date show that the conotoxin mixture produced by the venom gland varies not only at the inter- and intra-specific level [10,11], but also within a single individual over time [12], and among different regions of the venom gland [13]. In particular, the distal and the proximal segments of the venom gland, with respect to the venom bulb, produce different kinds of venom, in response to elicitation of a defensive or a predatory behaviour, respectively [13]. Despite the prevalence of conotoxins, other bioactive compounds have been found in conid venom, including insulins, c-type lectins, disulfide isomerase, and astacins [14,15,16,17,18,19,20].
Recent phylogenetic reconstructions, based on exomes and mitogenomes, confidently identified Profundiconus as the sister-lineage to all the other Conidae, dating its divergence from the other genera around 56–62 Mya [21,22]. As suggested by its name, the 28 extant species ascribed to this genus live in, but are not all restricted to, very deep waters, mainly in the Indo-Pacific [23]. The phylogenetic position of Profundiconus, as sister to all other cone-snails, makes it a key taxon for understanding conid venom evolution enabling to infer the venom composition of the ancestor of all present-day cone-snails.
Due to the lack of direct observations, Profundiconus ecology and hunting strategy could only be inferred from their radular morphology, which indicates a likely vermivory [24]. Additionally, the find of a small octopus beak in the stomach of a Profundiconus smirnoides Tenorio, 2015 specimen suggested the ability, of at least this species, to prey on cephalopods [24,25].
This study describes, for the first time, the venom composition of Profundiconus. The venom gland (VG) and foot (F) transcriptomes of Profundiconus cf. vaubani (Röckel & Moolenbeek, 1995) and Profundiconus neocaledonicus Tenorio & Castelin, 2016 were analyzed and novel gene superfamilies, as well as other venom components involved in toxicity, toxin processing, or in enhancing venom activity, are reported here. The novelty of gene superfamilies found, the presence of putative turripeptides, and the high abundance of other venom proteins suggest that Profundiconus venom gland transcriptomes are significantly different from those of other conids.
2. Results
2.1. Analysis of Profundiconus Transcriptomes
The transcriptome sequencing of three venom glands (from Pvau1, Pvau2, and Pneo) and two foot samples (from Pvau2 and Pneo) provided a total of 179,582,083 paired raw reads, (M = 35,916,417; SD = 3,439,048). After assembling and filtering, the dataset was composed of 1,049,747 contigs (M = 209,949; SD = 114,381) (Table S1). VG contigs were translated into 11,070,979 ORFs, including 191,207 with a signal region. After filtering, by similarity with other venom components and by subcellular localization, and removing duplicates, 223 unique putative venom components were retrieved, including those enriched in the VG, all considered new because having ≥1 amino acid (aa) of difference from previously published sequences. The number of venom components varied between specimens, with 51 found in Pvau1, 108 in Pvau2, and 86 in Pneo. Of those, 18 were shared between the two P. cf. vaubani specimens, while 3 between Pvau2 and Pneo, and 1 between Pvau1 and Pneo. At the species level, 137 unique venom components were retrieved from P. cf. vaubani, 82 from P. neocaledonicus, and 4 were shared by both.
OrthoVenn2 was used to compare Pvau2 and Pneo venom gland and foot transcriptomes. The analysis identified a total of 55,827 putative orthologous clusters (Figure S1). Overall, the four transcriptomes appeared to be quite different with a low percentage of clusters shared by all them (18%). The Pvau2 and Pneo foot samples were the most diversified, showing a higher number of total clusters compared with the respective venom glands (Pneo: F 164,539 vs. VG 60,736; Pvau2: F 118,640 vs. VG 79,727). The analysis also detected a higher similarity between similar tissue types than between species. Specifically, the two venom gland transcriptomes, with 29,485 shared clusters, appeared to be overall the most similar. The foot samples showed also a higher number of tissue specific clusters (F 9046 vs. VG 2908). Whereas, the foot and the venom gland of Pvau2 showed a lower overlap (25,606), comparable to the overlap between Pneo F and VG (24,342).
Clusters were grouped by gene ontology (GO) process and molecular function classes. We did not detect a marked difference in the most abundant GO classes present in foot and in venom glands samples. However, when considering the less abundant GO classes, venom glands were more diversified. In particular, in both tissues the four most abundant GO process classes (56–61% of the total) were annotated as involved in “biological”, “cellular”, and “metabolic processes”, and “biological regulation”. In addition, in the venom glands, we found 5% of the clusters involved in “RNA metabolic processes”, while this group was not detected in the foot samples. The situation was similar regarding GO molecular functions classes, with “ion binding”, “nucleic acid banding”, and “hydrolase activity” among the most abundant ones in both tissue (55–61% of the total). In the venom glands, clusters labelled as “transporters” (5%) and “extracellular region” (2%) were also detected.
2.2. Molecular Types and Putative Targets of Profundiconus Venom Components
In all three Profundiconus VG transcriptomes, putative neuropeptides represented the most diversified group (59–80% of the total diversity, total n = 148), followed by the hydrolases (8–28%, n = 45), while oxidoreductases, protease inhibitors, and lectins were present only in small amounts (≤5%, n < 10) (Figure 1 and Figure S2). Unique isolated neuropeptides were identified as: Conotoxins (123), profunditoxins (for definition see Paragraph 5.4) (7), turripeptides (13), cono-insulin (2), cono-cardioactive peptide (1), and prohormone-4 (2) (Figure 2). Seventy-five unique peptides of other putative venom compounds were found, and more than half were exclusively present in Pvau2 (Pvau1 = 10, Pvau2 = 44, and Pneo = 22) (Figure 3). A majority of the other putative venom components were represented by one or two types of peptides, with the exception of glycosyl hydrolases, which was particularly diversified in Pvau2 (nine peptides).
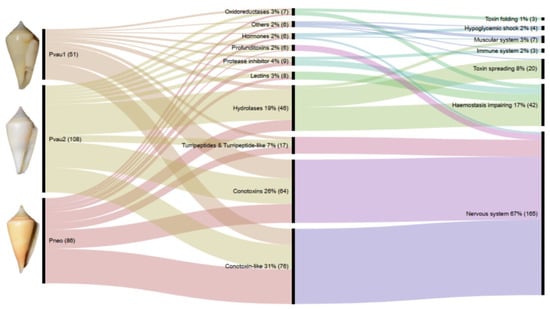
Figure 1.
Comparison of Profundiconus venom peptides and their potential application. Shown is an alluvial diagram made with RAWGraphs [26] composed of three steps (vertical lines) summarizing types of peptides found in the venom gland (centre) of each specimen (left), and their potential physiological target or effects (right). Flows are proportional to number of peptides. Numbers represent percentages and absolute numbers (between parenthesis) of transcripts found in the relative category. On the left, voucher photos of dissected specimens: P. cf. vaubani 1 (MNHN-IM-2013-66002, 20.7 mm length), P. cf. vaubani 2 (MNHN-IM-2013-69344, 22 mm length), and P. neocaledonicus (MNHN-IM-2013-69343, 24.5 mm length); copyright MNHN.
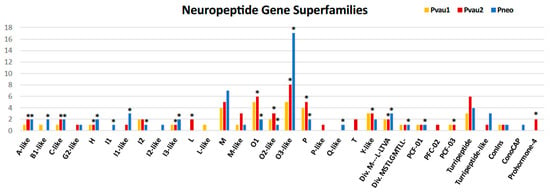
Figure 2.
Number of putative neuropeptides found in the venom glands (VG) of Profundiconus specimens categorized by gene superfamilies. Asterisks indicate specimens and gene superfamilies in which overexpressed peptides were found.
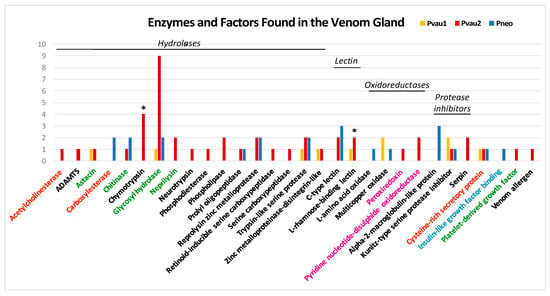
Figure 3.
Number of non-neuropeptide venom components of Profundiconus specimens, categorized by family. Asterisks indicate the specimens and families in which overexpressed peptides were found. Category colours indicate putative compounds targets/effects: orange = muscular system; black = haemostasis impairing; green = easing toxin spreading; fuchsia = toxins folding; light blue = hypoglycaemic shock.
Based on the potential molecular function of each transcript, 58–78% were putative toxins targeting the nervous system, 13–22% could affect haemostasis, while 2–12% could facilitate toxins spreading (Figure 1 and Figure S2). In all three Profundiconus VG specimens, a limited number of transcripts with other putative functions were also found, including toxins inducing a hypoglycaemic shock or impairing the muscular system. Only in Pneo and Pvau2 were toxins targeting the immune system found, and only in Pvau2 were venom components involved in toxin post-translational modifications and folding detected.
The differential expression analysis of the two specimens from which transcripts from both VG and F were available (Pvau2 and Pneo) identified 41 transcripts overexpressed in the VG, all neuropeptides except two. In Pneo, 21 conotoxin precursors and one profunditoxin precursor were overexpressed in the VG, while in Pvau2 15 conotoxin precursors, one profunditoxin precursor, one prohormone-4 (neuropeptide), one chymotrypsin (peptidase S1, hydrolase), and one L-rhamnose-binding lectin appeared overexpressed (Figure 2 and Figure 3 and Figure S3 and Table S2).
2.3. Analysis of Putative Conotoxins and Profunditoxins Transcripts
Of the total 130 unique conotoxin and profunditoxin precursors, 37 were found in Pvau1, 53 in Pvau2, and 55 in Pneo (Figure S4). The three VG specimens did not share identical precursors, since only 15 precursors are shared among the two P. cf. vaubani specimens and one is shared by Pvau1 and Pneo. As a result, 74 unique precursors for P. cf. vaubani, 54 for P. neocaledonicus, and 1 shared by both species were found (Figure S4).
Precursors were classified into 27 gene superfamilies, 17 of which appeared to be new (Table 1). Among the already known superfamilies (28,900 TPM from Pneo and Pvau2), conotoxin precursors were ascribed to the H, I1, I2, L, M, O1, P, and T superfamilies and to the two “divergent” superfamilies M---L-LTVA and MSTLGMTLL-, so called in the Conoserver database because they were first identified in the phylogenetically divergent species Californiconus californicus (Reeve, 1844) [27]. Among the remaining gene superfamilies (15,956 TPM from Pneo and Pvau2), 14 were similar to already described gene superfamilies (A-, B1-, C-, G2-, I1-, I2-, I3-, L-, M-, O2-, O3-, P-, Q-, and Y-like), while three were different from all conotoxins described so far, and therefore named profunditoxins (PFC-01, PFC-02, and PFC-03) (Figure 4). The three VG specimens did not show a great difference in number of identified gene superfamilies (Pvau1 = 18, Pvau2 = 22, and Pneo = 21) (Table 1 and Figure S4). More than half of the 15 gene superfamilies were shared by the three specimens, including the PFC-01 superfamily and the four most diversified ones (O3-like, M, O1, and P). Notably, eight gene superfamilies were unique to one of the studied specimens, namely: L-like in Pvau1; I1, I2-like and Q in Pneo; L, T, P-like and PFC-02 in Pvau2. Eighteen gene superfamilies were present in both species, while only six were found only in P. cf. vaubani (L, L-like, T, P-like, PFC-02, and PFC-03) and three in P. neocaledonicus (I1, I2-like, and Q).

Table 1.
Cysteine frameworks found in conotoxin, profunditoxin, and turripeptide gene superfamilies. 1 indicates cysteine frameworks never described before for that gene superfamily. Numbers represent total transcripts found in each VG for each gene superfamily, and between parentheses those overexpressed in the VG (if any) for the two specimens for which differential expression analysis were conducted (Pvau2 and Pneo).
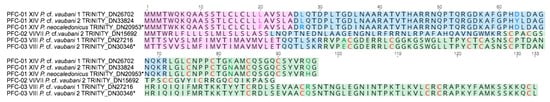
Figure 4.
Putative Profundiconus new gene superfamilies PFC-01, PFC-02, and PFC-03 separately aligned. In the transcripts, signal regions are displayed in fuchsia colour, pro region in light blue, and putative mature peptide in green. Cysteine residues are coloured in red. Labels for each transcript indicate gene superfamily, cysteine framework, species, and sequence or transcript number. The asterisk (*) indicates Profundiconus sequences overexpressed in the venom gland.
Among the profunditoxin gene superfamilies, PCF-01 showed a signal sequence of 25 aa, a pro-region of 46 aa, and a mature region of 25 aa. Three very similar sequences (2–6 divergent aa) were detected, one in each specimen, all sharing the XIV cysteine framework. The sequence identified in Pneo was overexpressed in the VG. The unique sequence of the PCF-02 superfamily was found in Pvau2, and it was composed of a 21 aa signal sequences, a 39 aa pro-region, and a 29 aa mature peptide showing the VI/VII cysteine pattern. Finally, two sequences with the VIII cysteine framework belonging to the PCF-03 superfamily were found, one in each P. cf. vaubani specimen. The sequence found in Pvau2 was overexpressed in the VG.
The 38 overexpressed conotoxin and profunditoxin precursors identified belonged to 19 gene superfamilies, 11 categorized as new, including PFC-01 and PFC-03 (Table 1). In Pvau2, 17 overexpressed precursors were found belonging to 13 gene superfamilies, while in Pneo, we retrieved 22 overexpressed precursors from 16 gene superfamilies. Seven precursors overexpressed in Pvau2 were also found in Pvau1, for which it was not possible to compute contig differential expression. Nine gene superfamilies were overexpressed in both specimens, namely: A-like, C-like, H, I3-like, O1, O2-like, O3-like, P, and M---L-LTVA. Only three were found in Pvau2 (L, Y-like, and PCF-02) and seven were found in Pneo (B1-like, I1, I1-like, I2, Q-like, PCF-01, and MSTLGMTLL-).
Conotoxin diversity values calculated by the Shannon’s Diversity Index were very similar between Pvau1 (H’ = 2.68) and Pneo (H’ = 2.61), while slightly higher in Pvau2 (H’ = 2.89). Instead, the Shannon’s Evenness Index was higher in the two P. cf. vaubani specimens (EPkan1 = 0.81; EPkan2 = 0.78) compared with the P. neocaledonicus one (EPter = 0.62). Combining results of the two P. cf. vaubani specimens, a species diversity index of 2.86 and an evenness of 0.69 were obtained.
A total of 19 different cysteine frameworks (including absence of cysteine) were found from the putative conotoxins, profunditoxins, and turripeptides, some of them present in more than one specimen (Pvau1 = 12; Pvau2 = 13; Pneo = 15). Ten of these frameworks were shared by all specimens: One shared by two, and six found in one specimen only (Table 1). The most abundant in all the three specimens were those with cysteine framework VI/VII (28–39%), followed by peptides without cysteine (15–19%). Thirteen conotoxin gene superfamilies had one or more cysteine frameworks that were not previously described for that superfamily. Five odd cysteine numbers (1 to 9) were also found in putative conotoxins, conotoxin-like, and turripeptides. Odd cysteine numbers have been reported from other cones species [15,27,28] and can be explained as belonging to homodimeric toxins [29] or originated from alternative RNA editing and splicing [30]. However, proteomic and experimental analyses will be needed to confirm that these odd numbered transcripts are toxins.
2.4. Analysis of Putative Turripeptide Trancripts
Nine unique putative turripeptides were identified (Pvau1 = 3, Pvau2 = 4, and Pneo = 3), presenting four cysteine frameworks (0, C, XVI, and IX), three of which shared by Pvau2 and Pneo (Figure 2). Four unique turripeptides, present in Pvau2 and Pneo, were similar to a previously identified turripeptide group characterized by a signal sequence similar to that of the B2 conotoxin gene superfamily, but lacking cysteine residues [31]. Profundiconus peptides belonging to this group lacked cysteine residues too and were similar to turripeptides found in Gemmula speciosa (Reeve, 1842) (D5KXH4), Unedogemmula bisaya (Olivera, 2004) (Ubs_01 and Ubs_15), and Crassispira cerithina (Anton, 1838) (Ccr_06 and Ccr_08). Two other peptides, found in all Profundiconus specimens, belonged to the turrid Pg gene superfamily, first identified in G. speciosa [32].
Four turripeptide-like compounds were also identified (Pvau2 = 1 and Pneo = 3), all with the IX cysteine framework (Figure 2). Two of these sequences, found in Pneo only, were similar to a turripeptide found in Iotyrris olangoensis [33] (OL135-[34]) while the other two, found in Pvau2 and Pneo, were similar to a turrid gene superfamily isolated by Olivera and colleagues [35]. This P-like superfamily is broadly expressed across turrids, but Profundiconus sequences appeared to be particularly similar to the sequence Pal9.2 from Polystira albida (G. Perry, 1811). In two turripeptide-like transcripts (TRINITY_DN29555 from Pvau2 and TRINITY_DN20308 from Pneo) a kazal domain was found, indicating that they may also possess a serine protease inhibitor activity.
2.5. Analysis of Prohormone-4 Transcripts
Two unique sequences of the neuropeptide prohormone-4 (PH-4) from Pvau2 were isolated. One of these sequences (Contig6527) appeared overexpressed in the VG (Figure 5). Prohormone precursors are composed of a signal peptide, a short mature region delimited by a mono or dibasic cleavage site (KR), and a precursor-related peptide. In some cases, there can be multiple cleavage sites producing multiple mature peptides (polyprotein hormone precursor). Profundiconus PH-4 precursors were 539 (Contig6527) and 567 (Contig6529) aa long and structured as follows: i) A signal sequence; ii) a mature sequence, different from the others found so far, followed by a second type of mature peptide, repeated in tandem three times (in the overexpressed Contig6527) or four times (Contig6529), each delimited by predicted cleavage sites; iii) a long post region similar to the post region found in the “prohormone-4-like” Conus group; iv) two cysteines in the tail region of the sequence. This structure suggested that Profundiconus PH-4 is a polyprotein hormone precursor, producing two types of mature peptides, one of which in multiple copies.

Figure 5.
Alignment of Profundiconus Prohormone-4 precursors to known sequences from other taxa. In the transcripts, the signal regions are highlighted in fuchsia, putative mature peptides in green, cleavage sites in blue, Low-Density Lipoprotein (LDL) receptors in orange, and cysteine-free regions in yellow. Amino acids are coloured in greyscale based on Blosum62 similarity score (black 100%). The asterisk (*) indicates the Profundiconus sequence overexpressed in the venom gland. § indicates sequences from Robinson et al. [28] supplementary materials.
PH-4 was first identified in the honeybee and linked to foraging predisposition (preference in collecting nectar or pollen) [36]. It has since been found in the brain of male eastern rock lobster [37] and in the venom gland of Conus victoriae, Conus geographus, Conus tessulatus, Conus varius, and Conus virgo [28], however its role in the envenomation process remains to be determined. Conus PH-4 has a mature sequence similar to the one retrieved in the honeybee and a precursor-related peptide containing an LDL (Low-Density Lipoprotein) receptor A and a cysteine free tail. Noteworthily, Robinson and colleagues (2017) found a second class of ‘‘prohormone-4-like” that is characterized by a longer precursor-related peptide (before the LDL receptor) and a different mature sequence. Of this second group, only one complete sequence was identified from C. tessulatus, but partial sequences were found in Conus marmoreus, Conus victoriae, Conus bullatus, Conus geographus, Conus virgo, and Conus varians [28].
2.6. Analysis of Insulin Transcripts
Two sequences of Profundiconus insulin were isolated: One in the two specimens of P. cf. vaubani, the other in P. neocaledonicus only (Figure 2). In molluscs, this well-known peptide hormone acts as a regulator of the glucose metabolism and as a neurotransmitter, and is derived from a precursor composed of a signal sequences and two chains (A and B) linked by a connecting peptide (C) [38]. In cone snails, two types of insulin have been found so far: A conventional signalling insulin, expressed in the nerve ring and probably involved in mollusc energy metabolism, and a derived insulin, expressed in the venom gland and involved in prey capture [20]. While the former is presumably present in all conids, with a conserved sequence, the latter was only detected in some species and shows higher variability [20]. Moreover, differences have been observed between venom insulin produced by fish-hunter Conus species compared with that of mollusc- and worm-hunter species. As a general trend, fish-hunter Conus venom insulin is similar to insulin found in fish, while mollusc- and worm-hunting Conus insulins are more similar to other mollusc signalling insulins [20,39].
The two Profundiconus predicted insulins were 129–130 aa length and had a sequence identity of 84% for the A chain and 74% for the B chain. The insulin chains of Profundiconus species showed different results when compared with those from other Conus species (Table 2 and Figure S5). Overall, Profundiconus insulin chains showed high similarity and identity with those from worm- or mollusc-hunting cones. While B chains from both Profundiconus species did not show a higher affinity to any one of these two groups, P. neocaledonicus insulin A chain was clearly more similar to molluscivorous Conus (Cylinder) textile Linnaeus, 1758. Alternatively, the A chain of the insulin from P. cf. vaubani showed slightly higher similarity to the A chain of three species of worm-hunting cones: Conus (Lividoconus) floridulus A. Adams & Reeve, 1848, Conus (Virroconus) ebraeus Linnaeus, 1758, and Conus (Strategoconus) planorbis Born, 1778. Notably, Profundiconus insulins also showed one additional cysteine residue on each chain, a feature present in venom insulins of worm- and mollusc-hunters, and also in signalling insulins from conids and other molluscs. These cysteines are supposed to form a third interchain disulfide bond, in addition to the two already existing between the chains A and B [39].
Table 2.
Comparison of Profundiconus A and B insulin chains. Profundiconus species are compared to Conus species grouped by diet (fish-, mollusc-, or worm-hunters), and maximum and minimum values are reported. Identity and similarity percentage inside each diet group are also listed (Intragroup). In bold, the highest identity/similarity values for each combination and, indicated by a number, the Conus species to which they refer: 1-Conus (Cylinder) textile Linnaeus, 1758, 2-Conus (Cylinder) victoriae Reeve, 1843, 3-Conus (Strategoconus) varius Linnaeus, 1758, 4-Conus (Lividoconus) floridulus A. Adams & Reeve, 1848, 5-Conus (Virroconus) ebraeus Linnaeus, 1758, 6-Conus (Strategoconus) planorbis Born, 1778, and 7-Conus (Virgiconus) virgo Linnaeus, 1758).
2.7. Analysis of Lectin Transcripts
Eight Profundiconus transcripts were identified as lectins. Five of the sequences found in Pneo and Pvau2 were identified as C-type lectin, while three were L-rhamnose-binding lectin (RBL) from the two specimens of P. cf. vaubani. RBLs are composed of one to three tandem carbohydrate recognition domains and have been classified in five types according to the number of tandem domains. Each domain is characterized by two conserved motifs (YGR and DPC) responsible for ligand specificity, and four conserved disulfide bounds [40]. One complete type I RBL sequence (three tandem domains) was found in Pvau2 (Contig1167) and overexpressed in the VG (Figure 6). The three domains are similar in sequences to the corresponding domain in Crassostrea gigas (Thunberg, 1793) (35–45% depending on the domain), but structurally homologous to those from the venomous sea urchin Toxopneustes pileolus (Lamarck, 1816) [41]. The two first domains have conserved motifs, while the third has a proline deletion in the DPC motif. Modifications of the first and second aa of this motif were found in anthozoan, bivalves, ascidian, and fish, however, this is the first reported finding of deletions.

Figure 6.
Alignment of Profundiconus L-rhamnose-binding lectin domain and other marine animals. Highlighted in light blue are the two conserved L-rhamnose-binding lectin domain motifs (YGR and DPC), responsible for ligand specificity. The four conserved cysteines are in yellow. Amino acids are coloured in greyscale based on Blosum62 similarity score (black 100%). In transcript sequence names, “DX” indicates the domain number.
RBLs have been mainly studied from fish eggs [42,43] and from globiferous pedicellariae of T. pileolus [41]. They were found also in colonial ascidians [44] and in bivalves, like the penguin wing oyster [45]. RBLs are involved in several processes including mitogenic, chemotactic, cytolytic, and apoptotic activities [41,46,47]. They also showed hemagglutination activity on rabbit erythrocytes, that can be inhibited by L-rhamnose and/or other saccharides [48].
2.8. Analysis of Chymotrypsin Transcripts
Three complete serine protease chymotrypsin-like sequences were found in Pvau2, one of which (TRINITY_DN34379) was overexpressed in the VG (Figure S6). Serine proteases are digestive enzymes characterized by a conserved catalytic triad (H/D/S) involved in the enzymatic activity. However, in some unconventional serine proteases, this triad is modified, like in the sedolisin proteases found in Pseudomonas sp., which have a different catalytic triad (S/D/E) and are active also at low pH conditions, e.g., hot springs or human lysosomes [49,50]. Among serine proteases, chymotrypsins are proteolytic enzymes involved in several intra- and extracellular activities. As venom components, they act on the coagulation cascade, on the fibrinolytic systems, and on platelets causing imbalance of the prey haemostatic system [51]. Profundiconus transcripts displayed sequence similarity with bivalves (30%) and snakes (25%) serine proteases, and structural similarity with chymotrypsins. However, the catalytic triad is the same as in Pseudomonas sp. sedolisin (S/D/E).
3. Discussion
This study represents the first portrait of the venom composition of two Profundiconus species, P. cf. vaubani and P. neocaledonicus. The sister position of Profundiconus to the rest of the family Conidae [21,22] makes it an important piece to the conid venom puzzle for interpreting conotoxin diversity and evolution. Our findings suggest that the Profundiconus venom arsenal is significantly dissimilar to those of other Conus, in particular for the gene superfamilies found, the presence of putative turripeptides, and the high abundance of other venom proteins.
3.1. P. cf. Vaubani Displayed a High Diversity of Nonneuropeptide Venom Components
While all the VG transcriptomes appeared largely similar in terms of putative conotoxin and turripeptide diversity, Pvau2 showed a definitively higher number of other venom components. Because prior conid literature has focused on neuropeptides, few works discussed the presence and diversity of other molecular types of venom components. Therefore, it is difficult to determine if the high percentages of non-neuropeptide sequences observed in P. vaubani can be considered ubiquitous among conids or peculiar to this species.
P. cf. vaubani venom appeared to be composed, in large part, of putative toxins potentially involved in haemostasis impairing, affecting prey muscular and immune system, and a smaller proportion of enzymes responsible for correct toxin folding or facilitating their spreading into prey tissues. For example, RBLs and chymotrypsins, which impact the coagulation cascade, were found exclusively in P. cf. vaubani specimens and enriched in Pvau2 VG, suggesting potential use for impairing prey haemostasis. These two compounds are both found in other animal venoms, and were overexpressed in P. cf. vaubani VG, which suggests they may have an important role in the envenomation process. Of note is the P. cf. vaubani chymotrypsin, which is similar to chymotrypsins active at low pH, and could represent an adaptation to a specific hunting strategy or prey type. Another interesting protein overexpressed in the VG of P. cf. vaubani is PH-4. P. cf. vaubani PH-4 showed a different structure and mature peptide sequence from those found in other conids and in honeybees, and possessed two types of mature peptides, one of which in multiple copies. In honeybees, PH-4 was linked to foraging-related behaviour, however the lack of information concerning Profundiconus ecology and hunting strategy hampers the evaluation of PH-4, RBLs, and chymotrypsins and their potential effects on P. cf. vaubani prey.
3.2. Limited Conotoxin Diversity May Indicate a Narrow Worm or Molluscan Diet
Analysis of our transcriptome assemblies recovered different numbers of putative conotoxin and profunditoxin precursors: 75 for P. cf. vaubani and 55 for P. neocaledonicus. If the three specimens are considered independently, the number of putative conotoxin and profunditoxin precursors are: 37 (Pvau1), 53 (Pvau2), and 55 (Pneo). If only the conid species sequenced in conditions similar to the ones used in the present work (one to three VG samples, sequenced by Illumina HiSeq2000 platform) are taken into account, P. neocaledonicus emerges as the species showing the lowest number of conotoxin and contoxin-like precursors. P. cf. vaubani showed instead a higher number of conotoxin precursors, equal to the one in Conus (Gastridium) geographus Linnaeus, 1758 [52] and C. (Virroconus) ebraeus [53], from which, however, only one specimen was sequenced. Therefore, if we take into consideration only the result of one specimen for each Profundiconus species (Pneo, and the most numerous Pvau2 with 53 conotoxins), P. neocaledonicus and P. cf. vaubani may be included among those conids with a less diversified neuropeptide arsenal.
Conversely, when the numbers of gene superfamilies produced in the same sequencing condition are considered, P. cf. vaubani and P. neocaledonicus show a high number of superfamilies (24 and 21, respectively) comparable to species with higher numbers of conotoxins, like Conus (Virgiconus) virgo Linnaeus, 1758 with 25 gene superfamilies and 113 conotoxins [53] and C. (Gastridium) geographus Linnaeus, 1758 with 21 gene superfamilies and 75 conotoxins [52]. The two Profundiconus species display a similar profile in terms of types and abundance of gene superfamilies. In fact, although with different percentages, the superfamilies O3-like, M, O1, and P appeared to be the most diversified in all the three Profundiconus specimens. The superfamilies M and O1 are among the most common and abundant conid gene superfamilies (e.g., [49,50]), so it is not unexpected to find them well represented in Profundiconus. The O3 superfamily, the most closely related to the newly described O3-like superfamily, is present in several species, but not abundant throughout conids. It was found in small numbers (up to seven different conotoxins) in 13 conid species with different feeding habits such as fish-, mollusc-, and worm-hunting [53,54]. Interestingly, this gene superfamily, along with the J and T ones, were overexpressed in the distal part of the VG of C. (Gastridium) geographus, accounting for ~50% of the total conotoxin reads found in this segment that is supposed to be the one producing predation venom, but for only ~5% in the proximal segments of the VG, the one producing defence-evoked venom [13,55]. The P superfamily was found in several fish-, mollusc-, and worm-hunting Conus species but always with low diversity (up to 14 different conotoxins), with the exception of Conus (Turriconus) praecellens A. Adams, 1855, in which this superfamily was the most abundant and diversified [52]. Remarkably, conotoxins belonging to the T gene superfamily, which are frequently common and diversified in the other conid species, are lacking in the venom gland transcriptome of the Profundiconus species analyzed here.
The overexpressed conotoxin and profunditoxin fraction, as calculated by differential expression analysis on TMP values, was quite similar in composition between P. cf. vaubani and P. neocaledonicus. In fact, 40% of the gene superfamilies found were overexpressed in both species, and overall, 50–70% of them included at least one transcript overexpressed in the VG. These results suggest that the venom cocktails of P. cf. vaubani and P. neocaledonicus may be not identical but for a large part similar, at least for what concern the conotoxin composition.
Comparing Profundiconus Shannon’s diversity and evenness indexes with those of other conid species sequenced in similar conditions, like C. tribblei (H’ = 3.30, E = 0.90) and C. lenevati (H’ = 3.30, E = 0.89) [56], confirmed what was already suggested by conotoxins abundance. P. cf. vaubani and P. neocaledonicus showed slightly lower conotoxin diversities compared with other species and a less homogeneous distribution of transcripts among gene superfamilies. However, the pipeline we used to identify putative toxins in Profundiconus tissues was largely based on similarity with conotoxins, potentially leaving highly divergent profunditoxins undetected, and leading to an underestimation of the real toxin diversity in Profundiconus. Pipelines that do not rely mainly on similarity with previously described toxins but more on, e.g., their structural properties, are required to detect the eventual new toxins of this divergent genus that may be different from what found in Conus up to now. This is particularly true when it is taken into account that the range of conotoxin precursors and gene superfamilies reported, the number of specimens used (1–20), and the types of sequencing platforms (five types) vary greatly among the conid VG transcriptomes produced to date: From 30 conotoxin precursors and 6 gene superfamilies in Conus (Textilia) bullatus Linnaeus, 1758 [14], to 401 conotoxin precursors in Conus (Harmoniconus) sponsalis Hwass in Bruguière, 1792 [53] and 55 gene superfamilies in Conus (Chelyconus) ermineus Born, 1778 [54] (Table S3). Two studies reported even higher numbers of conotoxin precursors found in a single species: 662 in Conus (Rhizoconus) miles Linnaeus, 1758 [57] and 3305 in Conus (Darioconus) episcopatus da Motta, 1982 [58] (Table S3). These studies, reporting exceptionally high putative conotoxins numbers, may be the only ones that have been able to detect minor conotoxin variants already found in proteomic studies, but may also be a result of sequencing data processing [52,53]. This lack of homogeneity, along with the use of different bioinformatic pipelines and threshold criteria, the physiological intraspecific variation of the venom composition [11], and the lack of corroborating proteomic data for much of the reported findings, demands caution in comparing numbers resulting from different projects.
In conid literature, venom complexity has always been related to prey preference. However, recent studies [53,59] pointed out that conotoxin diversity is correlated to diet breadth more than prey type, revealing the tendency of species with more generalized diets to have more complex venoms and more predation-evoked venom genes. Almost nothing is known about Profundiconus ecology, but the limited data available suggest a worm-hunting diet with at least one species able to prey on fast moving molluscs [24,25]. The venom insulins found in Profundiconus are similar to those of worm- and mollusc-hunting Conus species and have an additional cysteine residue characteristic these species groups. As a result, based on transcriptomic analyses, P. cf. vaubani and P. neocaledonicus diets may include only a limited diversity of worm and/or mollusc preys.
3.3. Characterization of the First Putative Profunditoxin Gene Superfamilies Indicate They Are Divergent
Three completely new gene superfamilies were predicted, two of which are overexpressed in the VG, and therefore potentially represent main Profundiconus venom components. The high level of sequence divergence between the three gene superfamilies suggests three unrelated functions. Moreover, the presence of almost identical sequences of PFC-01 in both species may suggest a broader spread of this gene superfamily across Profundiconus species. Considering the phylogenetic position of Profundiconus as an independent lineage sister to the rest of the Conidae, these toxin superfamilies may be classified as “divergent” [60,61]. However, because of the high divergence of Profundiconus from the other cone snails, we expected to find more profunditoxins superfamilies, especially when considering that new Conus gene superfamilies continue to be routinely discovered, as recently reported for C. tribblei and C. lenevati [56,62].
3.4. Turripeptides Retained in Profundiconus Venom
In Profundiconus, turripeptide-related transcripts were among the most diversified neuropeptide classes. Turripeptides and conotoxins are thought to have a common evolutionary origin as they show similar precursor organization of signal, pro mature, and post regions [35]. However, when the first turrid venom peptides were discovered, little overlap was found with conotoxins, and the large majority of them belonged to new gene superfamilies, not yet found in conids [31,32,34,35]. Later, the discovery of peptides similar to conotoxins or turripeptides in distant taxa, likely as the results of convergent evolution, underlined the broader benefits obtained by recruiting them as venom or secretion components. For example, turripeptide-like toxins were found among the feeding-related proteins of the vampire snail Colubraria reticulata (Blainville, 1829) [63], in the hunting venom of bloodworms [64], and in the defensive one of fireworms [65], while conotoxin-like peptides were found in the mussel Mytilus galloprovincialis Lamarck, 1819 [66]. If the present distribution of turripeptides among taxa is considered in the context of the conoidean phylogeny, they might be present in the Conoidea common ancestor, and perhaps even earlier. Eventually, in the Conidae, some turripeptides (what we currently call conotoxins, found in both turrids and conids) started to rapidly diversify, becoming the prevalent component of conid venom, up to the complete loss of turripeptides at least in the genus Conus. The early divergence of Profundiconus from the rest of Conidae may explain why some turripeptide gene superfamilies are still retained in this genus. However, this hypothesis needs to be corroborated by characterizing the VG components of more Profundiconus species, and by a more extensive comparison of turripeptides from other groups belonging to the conid radiation, such as Conasprella, Lilliconus, and Pygmaeoconus, for which limited, or no data, are available.
4. Conclusions
This is the first study to explore the toxin diversity in the venom gland of the genus Profundiconus, the earliest offshoot of cone snails. In the Profundiconus species examined, P. cf. vaubani and P. neocaledonicus, putative conotoxin numbers were not particularly high, however the large majority of them belonged to potentially new gene superfamilies, with three gene superfamilies highly divergent from conotoxin superfamilies described to date. The presence of some rare conid gene superfamilies and the absence of those largely widespread in other cone snails, along with the occurrence of putative turripeptides and a relevant percentage of non-neuropeptides components, constitute uncharacteristic features of Profundiconus venom composition with respect to the other cones investigated to date. Importantly, the observation of prohormone-4 and lectins provide a first overview about Profundiconus venom being potentially involved in prey behavioural alteration and haemostasis impairing. Additionally, the low conotoxin diversity and the presence of a signalling-like venom insulin in all specimens offer clues about the poorly known trophic ecology of these species, suggesting that P. cf. vaubani and P. neocaledonicus may prey on a limited number of species, possibly worms or other molluscs. Finally, our results indicate that, even if several cone snail venom gland transcriptomes have been studied, we are still far from reaching the plateau of conotoxins diversity. The venom composition of Profundiconus calls for further investigation of venom gland transcriptomes in neglected Conidae lineages, and for a more detailed analysis of those from previously studied species, in order to fully understand the venom arsenal of this family and its ecological and evolutionary importance.
5. Materials and Methods
5.1. Sample Collection and Identification
Three specimens of the genus Profundiconus were collected during the expedition KANACONO (doi 10.17600/16003900; expeditions.mnhn.fr) in New Caledonia, South of the Isle of Pines: MNHN-IM-2013-66002 (st. DW4666, 22°53′ S, 167°16′ E, 530–545 m), MNHN-IM-2013-69343 and MNHN-IM-2013-69344 (st. DW4722, 22°54′ S, 167°16′ E, 496–505 m). The specimen MNHN-IM-2013-66343 was morphologically identified as Profundiconus neocaledonicus (Pneo) Tenorio & Castelin, 2016. This identification was then confirmed by sequencing the Barcode fragment of the COI gene (results not shown). The specimens MNHN-IM-2013-66002 and MNHN-IM-2013-66344 most probably correspond to a white form of Profundiconus vaubani (Röckel & Moolenbeek, 1995), and their COI sequences also match COI sequences of P. vaubani from New Caledonia (Tenorio & Castelin, 2016). We will then refer to them as P. cf. vaubani (Pvau). In this work, the three specimens are referred as following: Pvau1 for MNHN-IM-2013-66002, Pvau2 for MNHN-IM-2013-69344, and Pneo for MNHN-IM-2013-69343. COI sequences were deposited in GenBank (accession numbers MN540394-6).
5.2. RNA Extraction, Sequencing, and De Novo Assembly
For Pvau2 and Pneo, a piece of foot was isolated, and for all the three specimens, the venom gland was dissected after crushing the shells (the shell remains were kept as vouchers). RNA was extracted using the trizol method. Bioanalyzer traces were used to assess total RNA quality and determine suitability for sequencing. The cDNA libraries were prepared and sequenced at the New York Genome Center. Libraries were prepared using the automated polyA RNAseq library prep protocol and sequenced with Illumina HiSeq 4000 with 150-bp paired-end reads. Trimmomatic v.0.36 [67] was used to remove adapter contamination and filter low-quality reads (ILLUMINACLIP option enabled, seed mismatch threshold = 2, palindrome clip threshold = 40, simple clip threshold of 15; SLIDING WINDOW option enabled, window size = 4, quality threshold = 20; MINLEN = 36; LEADING = 3; TRAILING = 3). Reads were merged using FLASH v.1.2.8 [68] with a min overlap parameter of 5, a max overlap parameter of 100, and a mismatch ratio of 0.05. Trinity v.2.4 was then used to assemble transcripts [69]. CAP3 [70] was finally used with default parameters and cd-hit v.4.6 (percent identity = 99%; [71]) to reduce redundancy in the assemblies. Transcriptome data are available at GenBank (BioProject PRJNA574281).
5.3. Transcriptome Annotation and Differential Expression Analysis
To identify Profundiconus venom components, a slightly modified standard pipeline was used [65,72]. Briefly, VG contigs were searched for all possible open reading frames (ORFs) with a minimum length of 10 aa using a homemade script (Script 1), and signal sequences were identified using SignalP. Two similarity searches (e-value 10−5) were then performed in parallel on filtered VG transcripts. The first was made using BLASTp v.2.6 (National Center for Biotechnology Information, Bethesda, MD, USA) [73] against a customized toxin database, built using proteins from ConoServer [60,74], UniProt Animal Toxin Annotation Project [75], and teretoxin sequences from Gorson et al. [72]. For the second, transcripts were searched against hidden Markov models (HMM) profiles built on toxins from the protein family Pfam database [76], using HMMER v.3.2 [77]. Pfam database was chosen as a complement to the homemade database because, being built through a different search strategy that takes into consideration the full protein domain similarity (e.g., also less-conserved regions and insertions), it includes in the toxin category sequences belonging to a wider range of organisms. This approach is particularly important for finding putative venom components in a genus, like Profundiconus, never studied before.
Transcript per million (TPM) values were calculated using Kallisto [78] for all the VG and F contigs of the two specimens for which both tissues were sequenced (Pvau2 and Pneo) (bootstrap = 100). Differential expression analyses with technical replicates were then performed between the contigs of the VG and the F of the same specimen, using the R package NOISeq v.2.22 (NOISeq-sim function, normalization = rpkm, length correction = 1 [79]). Those contigs overexpressed in the VG were added to the dataset, if not already present, after ORFs and signal sequences searches. All selected transcripts were searched using BLASTp against UniProt full database and NCBI nr database. Results were manually inspected, and transcripts with a higher hit to a non-toxin protein family were discarded. DeepLoc v.1 (protein encoding = Profiles [80]) was used to predict the subcellular localization of the remaining contigs, and only the extracellular ones were retained. Finally, identical contigs (100%) were identified with cd-hit v.4.6 [71], and duplicates within a single transcriptome were discarded. Protein sequences were deposited in UniProt.
OrthoVenn2 web platform [81] was used to compare the compounds expressed in the VG and F of Pvau2 and Pneo by grouping the ORF obtained into putative orthologues clusters (e-value 10−5) and annotating them using Gene Ontology biological classes.
5.4. Venom Component Identification and Diversity
Particular attention was given to proteins similar to previously identified venom components and to those overexpressed in the venom glands. When possible, for each transcript, the hypothetical molecular type, the protein family, and a potential molecular target were identified through sequence (BLASTp and HMMER) and structural (SMART [82] and SWISS-MODEL [83]) comparison with already-described proteins. All these assignations have to be considered preliminary, until confirmed by proteomics and bioactivity tests; consequently, all the functional annotations we propose have also to be considered putative. VG transcripts were divided into three putative functional groups: Components interfering with the physiology of the prey (toxins), those in charge of toxin post-translation modifications and folding, and peptides facilitating toxin spread in prey’s body. The toxin group included bioactive compounds putatively assigned to several molecular types: Neuropeptides, hydrolases, lectins, oxidoreductases, and protease inhibitors. Among them, the venom neuropeptide category included predicted peptides hypothetically affecting prey’ nervous system (conotoxins and turripeptides), muscular system (cardioactive peptides), immune system (allergens), or causing hypoglycaemic shock (insulins). The rest of the toxins were instead targeting prey’ hemostasis system (e.g., kunitz-type serine protease inhibitors, multicopper oxidases, C-type lectins, and trypsin-like serine proteases). The other two venom component categories respectively included oxidoreductases involved in toxin post-translation modifications and folding (e.g., pyridine nucleotide-disulfide oxidoreductase), and hydrolases easing toxins spreading (e.g., chitinase, astacins, and neprilysins). Profundiconus venom components were considered new when including at least one different aa compared to known ones.
Those Profundiconus transcripts similar to conotoxins and turripeptides, principal components of other conoidean venoms, were further investigated for identifying gene superfamilies and cysteine patterns. Conotoxin preliminary gene superfamily designations were based on results obtained from BLASTp and HMMER. The full sequences were analyzed with BLASTp, while the signal sequences only were searched against profiles of conotoxin precursor signal sequences retrieved from ConoServer. Gene superfamily hypothetical assignation, or both results in case of discordance between the two methods used, were tested in the following step. The average percentage sequence identity (=similarity score) of the signal region was calculated between each transcript and all conotoxins ascribed to the same preliminary gene superfamily (pPID). For each gene superfamily, the average percentage sequence identity of the signal sequences (sPID) and its standard deviation (sSD) were also calculated. A transcript was considered belonging to a known gene superfamily when pPID > sPID - sSD. For some small gene superfamilies, exceptionally high sPID values were retrieved. However, these values can be an underestimation of the real gene superfamilies diversity, mainly due to the fact that only a few sequences are available and for a limited number of Conus species. For this reason, in those gene superfamilies with a sPID > 76% (the average sPID value of the biggest conotoxin superfamily [9]), this score was instead used as a threshold. All transcripts showing similarity to conotoxins but with a pPID score under the respective threshold were assigned to the gene superfamily “X-like”, where X represented the most similar gene superfamily. Sequences showing an exceptional low pPID value (<40% as used in ConoServer) were instead categorized in completely new gene superfamilies (“PFC-XX”, for ProFundiConus-XX), and in this work named “profunditoxins” (defined as natural peptide toxins produced by Profundiconus, by analogy with conotoxins, turritoxins, and teretoxins [84,85]). Conotoxins and profunditoxins diversity was measured by Shannon’s diversity index (H’) and homogeneity among gene superfamilies were calculated by Shannon’s evenness index (E) (as in [62]) using Past3 [86].
For turripeptides, a unique and broadly used gene superfamily classification scheme is not available. Consequently, we calculated the percentage of sequence identity for the signal region between each transcript resulting similar to a turripeptide with BLASTp or HMMER and known turripeptides (tPID). Those transcripts with tPID > 76% were labelled as “turripeptide”, and the rest as “turripeptide-like”. A homemade script (Script 2) was used to identify cysteine pattern of conotoxins, conotoxin-like, profunditoxins, turripeptides, and turripeptide-like mature region. For the other protein families found in Profundiconus, homologous sequences from other molluscs and venomous animals were retrieved from NCBI and UniProt and the domain region was aligned using MAFFT v.7 (E-INS-i algorithm [87,88]). For insulin, Profundiconus chain A and B were aligned with those from 17 Conus species grouped by diet (fish, mollusc, and worm) (Figure S5). Intra and inter groups identity and similarity (Blosum62) percentages were then calculated.
Supplementary Materials
The following are available online at https://www.mdpi.com/2072-6651/11/11/623/s1, Figure S1: Graphical summary of clusters obtained for each sample, Figure S2: Pie-charts of transcripts putative molecular types and effect/target for each specimen, Figure S3: Plots of differential expression results between venom gland and foot of Pneo and Pvau2, Figure S4: Venn diagram of conotoxins and profunditoxins found in the VG of Profundiconus specimens, Figure S5: Alignment of insulin chain A and B, Figure S6: Serine protease domain alignment of sequence from Profundiconus, bivalves and snakes, Table S1: Sequencing, assembling and pipeline numbers for each tissue sample analyzed, Table S2: List of putative venom component found in the VG of Pvau1, Pvau2, and Pneo with molecular identification and amino acidic sequences, Table S3: Comparison of conotoxins, profunditoxins and gene superfamilies found in P. neocaledonicus and P. cf. vaubani with those of other conids, Script 1: Python script for identifying ORF in transcript sequences, Script 2: Python script for identifying cysteine patterns in amino acid sequences.
Author Contributions
Conceptualization, G.F., M.V.M. and N.P.; software, L.M.; formal analysis, A.E.F., G.F., J.G., Y.I.K., and P.Z.; investigation, G.F.; resources, M.H. and N.P.; writing—original draft preparation, G.F.; writing—review and editing, all authors.; project administration, N.P.; funding acquisition, M.H., A.E.F. and N.P.
Funding
This research was funded by Service de Systématique Moléculaire [UMS 2700 CNRS – MNHN], by the project CONOTAX, funded by the French Agence Nationale de la Recherche – France [ANR-13-JSV7-0013-01 to NP], and by the LABEX BCDiv [SU, MNHN, UPMC, CNRS, IRD, ANR, EPHE, Investissements d’avenir, Paris Diderot] and by research grants by the Camille & Henry Dreyfus Foundation and NIH-NIMHD grant 8-G-12-MD007599 to MH. GF support was provided by the fellowship “Borsa di Perfezionamento all’Estero 2016” from Sapienza University of Rome. JG support was partially provided by The CUNY Graduate Center Graduate Fellowship program. A.E.F. was funded by the research grant from the Russian Science Foundation, N 19-74-10020. M.V.M. was founded by the EU Horizon 2020 research and innovation programme under grant agreement No 748902.
Acknowledgments
The MHNH material was collected during the KANACONO expedition in New Caledonia (convention MNHN-Province Sud, APA_NCPS_2016_012; PI N. Puillandre and S. Samadi). This expedition operated under the regulations then in force in the country in question and satisfies the conditions set by the Nagoya Protocol for access to genetic resources. We thank Barbara Buge, and Julien Brisset for their help in curating the vouchers and databasing the information related to the expedition and the vouchers. We are grateful to Julio Pedraza and Jawad Abdelkrim for the help with the computer cluster of the Service de Systématique Moléculaire, MNHN. We thank Manuel Tenorio for the morphological identification of the specimens.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Paracelsus. Opera Omnia Medico-Chemico-Chirurgica, Tribus Voluminibus Comprehensa. Editio Novissima et Emendatissima ad Germanica & Latina Exemplaria Accuratissime Collata; Sumptibus Joan. Antonii & Samuelis De Tournes: Geneva, Switzerland, 1658. [Google Scholar]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The toxicogenomic multiverse: Convergent recruitment of proteins into animal venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef]
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex cocktails: The evolutionary novelty of venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Puillandre, N.; Kantor, Y.I.; Sysoev, A.; Couloux, A.; Meyer, C.; Rawlings, T.; Todd, J.A.; Bouchet, P. The dragon tamed? A molecular phylogeny of the Conoidea (Gastropoda). J. Molluscan Stud. 2011, 77, 259–272. [Google Scholar] [CrossRef]
- Abdelkrim, J.; Aznar-Cormano, L.; Buge, B.; Buge, B.; Kantor, Y.; Zaharias, P.; Puillandre, N. Delimiting species of marine gastropods (Turridae, Conoidea) using RAD sequencing in an integrative taxonomy framework. Mol. Ecol. 2018, 27, 4591–4611. [Google Scholar] [CrossRef]
- Phuong, M.A.; Alfaro, M.E.; Mahardika, G.N.; Marwoto, R.M.; Prabowo, R.E.; von Rintelen, T.; Vogt, P.W.H.; Hendricks, J.R.; Puillandre, N. Lack of signal for the impact of conotoxin gene diversity on speciation rates in cone snails. Syst. Biol. 2019, syz016. [Google Scholar] [CrossRef]
- Olivera, B.M.; Seger, J.; Horvath, M.P.; Fedosov, A.E. Prey-Capture Strategies of Fish-Hunting Cone Snails: Behavior, Neurobiology and Evolution. Brain Behav. Evol. 2015, 86, 58–74. [Google Scholar] [CrossRef]
- Puillandre, N.; Koua, D.; Favreau, P.; Olivera, B.M.; Stocklin, R. Molecular Phylogeny, Classification and Evolution of Conopeptides. J. Mol. Evol. 2012, 74, 297–309. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.; Craik, D.J. Conopeptide characterization and classifications: An analysis using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Rivera-ortiz, J.A.; Cano, H.; Marí, F. Intraspecies variability and conopeptide profiling of the injected venom of Conus ermineus. Peptides 2011, 32, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.M.; Dutertre, S.; Lewis, R.J.; Marí, F. Intraspecific variations in Conus purpurascens injected venom using LC/MALDI-TOF-MS and LC-ESI-TripleTOF-MS. Anal. Bioanal. Chem. 2015, 407, 6105–6116. [Google Scholar] [CrossRef] [PubMed]
- Prator, C.A.; Murayama, K.M.; Schulz, J.R. Venom Variation during Prey Capture by the Cone Snail, Conus textile. PLoS ONE 2014, 9, e98991. [Google Scholar] [CrossRef] [PubMed]
- Dutertre, S.; Jin, A.-H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Characterization of the Conus bullatus genome and its venom-duct transcriptome. BMC Genomics 2011, 12, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Terrat, Y.; Biass, D.; Dutertre, S.; Favreau, P.; Remm, M.; Stöcklin, R.; Piquemal, D.; Ducancel, F. High-resolution picture of a venom gland transcriptome: Case study with the marine snail Conus consors. Toxicon 2012, 59, 34–46. [Google Scholar] [CrossRef] [PubMed]
- Violette, A.; Leonardi, A.; Piquemal, D.; Terrat, Y.; Biass, D. Recruitment of Glycosyl Hydrolase Proteins in a Cone Snail Venomous Arsenal: Further Insights into Biomolecular Features of Conus Venoms. Mar. Drugs 2012, 10, 258–280. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Hu, H.; Gorasia, D.G.; Bandyopadhyay, P.K.; Veith, P.D.; Young, N.D.; Reynolds, E.C.; Yandell, M.; Olivera, B.M.; Purcell, A.W. Combined Proteomic and Transcriptomic Interrogation of the Venom Gland of Conus geographus Uncovers Novel Components and Functional Compartmentalization. Mol. Cell. Proteomics 2014, 13, 938–953. [Google Scholar] [CrossRef]
- Figueroa-Montiel, A.; Ramos, M.A.; Mares, R.E.; Duenas, S.; Pimienta, G.; Ortiz, E.; Possani, L.D.; LIcea-Navarro, A.F. In Silico Identification of Protein Disulfide Isomerase Gene Families in the De Novo Assembled Transcriptomes of Four Different Species of the Genus Conus. PLoS ONE 2016, 11, e0148390. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Li, Q.; Jackson, R.L.; Song, A.S.; Boomsma, W. Rapid expansion of the protein disulfide isomerase gene family facilitates the folding of venom peptides. Proc. Natl. Acad. Sci. USA 2016, 113, 3227–3232. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Lu, A.; Li, Q.; Fedosov, A.E.; Biggs, J.; Corneli, P.S.; Seger, J.; Yandell, M.; Olivera, B.M. Venom Insulins of Cone Snails Diversify Rapidly and Track Prey Taxa. Mol. Biol. Evol. 2016, 33, 2924–2934. [Google Scholar] [CrossRef]
- Uribe, J.E.; Puillandre, N.; Zardoya, R. Beyond Conus: Phylogenetic relationships of Conidae based on complete mitochondrial genomes. Mol. Phylogenet. Evol. 2017, 107, 142–151. [Google Scholar] [CrossRef]
- Abdelkrim, J.; Aznar-cormano, L.; Fedosov, A.E.; Kantor, Y.I.; Lozouet, P.; Phuong, M.A.; Zaharias, P.; Puillandre, N. Exon-Capture-Based Phylogeny and Diversification of the Venomous Gastropods (Neogastropoda, Conoidea). Mol. Biol. Evol. 2018, 35, 2355–2374. [Google Scholar] [CrossRef]
- MolluscaBase Profundiconus Kuroda, 1956. Accessed Through: World Register of Marine Species. 2019. Available online: http://www.marinespecies.org/aphia.php?p=taxdetails&id=428962 on (accessed on 4 September 2019).
- Tenorio, M.J.; Castelin, M. Genus Profundiconus Kuroda, 1956 (Gastropoda, Conoidea): Morphological and molecular studies, with the description of five new species from the Solomon Islands and New Caledonia. Eur. J. Taxon. 2016, 173, 1–45. [Google Scholar] [CrossRef]
- Marshall, B.A. New records of Conidae (Mollusca: Gastropoda) from the New Zealand region. New Zeal. J. Zool. 2012, 8, 493–501. [Google Scholar] [CrossRef]
- Mauri, M.; Elli, T.; Caviglia, G.; Uboldi, G.; Azzi, M. RAWGraphs: A Visualisation Platform to Create Open Outputs open output. In Proceedings of the 12th Biannual Conference on Italian SIGCHI Chapter ACM, New York, NY, USA, 18–20 September 2017; pp. 28:1–28:5. [Google Scholar]
- Elliger, C.A.; Richmond, T.A.; Lebaric, Z.N.; Pierce, N.T.; Sweedler, J.V.; Gilly, W.F. Diversity of conotoxin types from Conus californicus reflects a diversity of prey types and a novel evolutionary history. Toxicon 2011, 57, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Li, Q.; Bandyopadhyay, P.K.; Gajewiak, J.; Yandell, M.; Papenfuss, A.T.; Purcell, A.W.; Norton, R.S.; Safavi-hemami, H. General and Comparative Endocrinology Hormone-like peptides in the venoms of marine cone snails. Gen. Comp. Endocrinol. 2017, 244, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Quinton, L.; Gilles, N.; De Pauw, E. TxXIIIA, an atypical homodimeric conotoxin found in the Conus textile venom. J. Proteomics 2009, 72, 219–226. [Google Scholar] [CrossRef]
- Degueldre, M.; Verdenaud, M.; Legarda, G.; Minambres, R.; Zuniga, S.; Leblanc, M.; Gilles, N.; Ducancel, F.; De Pauw, E.; Quinton, L. Diversity in sequences, post-translational modifications and expected pharmacological activities of toxins from four Conus species revealed by the combination of cutting-edge proteomics, transcriptomics and bioinformatics. Toxicon 2017, 130, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, D.T.T.; Saloma, C.P. A bioinformatics survey for conotoxin-like sequences in three turrid snail venom duct transcriptomes. Toxicon 2014, 92, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Heralde, F.M.; Imperial, J.; Bandyopadhyay, P.K.; Olivera, B.M.; Concepcion, G.P.; Santos, A.D. A rapidly diverging superfamily of peptide toxins in venomous Gemmula species. Toxicon 2008, 51, 890–897. [Google Scholar] [CrossRef]
- Olivera, B.M. Conus Venom Peptides: Reflections from the Biology of Clades and Species. Annu. Rev. Ecol. Syst. 2002, 33, 25–47. [Google Scholar] [CrossRef]
- Watkins, M.; Hillyard, D.R.; Olivera, B.M. Genes Expressed in a Turrid Venom Duct: Divergence and Similarity to Conotoxins. J. Mol. Evol. 2006, 62, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Watkins, M.; Bandyopadhyay, P.; Imperial, J.S.; de la Cotera, E.P.H.; Aguilar, M.B.; Lopez Vera, E.; Concepcion, G.P.; Lluisma, A. Adaptive radiation of venomous marine snail lineages and the accelerated evolution of venom peptide genes. Ann. N. Y. Acad. Sci. 2012, 1267, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Brockmann, A.; Annangudi, S.P.; Richmond, T.A.; Ament, S.A.; Xie, F.; Southey, B.R.; Rodriguez-zas, S.R.; Robinson, G.E.; Sweedler, J. V Quantitative peptidomics reveal brain peptide signatures of behavior. Proc. Natl. Acad. Sci. USA 2009, 106, 2383–2388. [Google Scholar] [CrossRef] [PubMed]
- Ventura, T.; Cummins, S.F.; Fitzgibbon, Q.; Battaglene, S.; Elizur, A. Analysis of the Central Nervous System Transcriptome of the Eastern Rock Lobster Sagmariasus verreauxi Reveals Its Putative Neuropeptidome. PLoS ONE 2014, 9, e97323. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.; van Kesteren, R.; Li, K.; van Minnen, J.; Spijker, S.; van Heerikhuizen, H.; Geraerts, W. Toward Understanding the Role of Insulin in the Brain: Lessons from Insulin-Related Signaling Systems in the Invertebrate Brain. Prog. Neurobiol. 1998, 54, 35–54. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1–6. [Google Scholar] [CrossRef]
- Ogawa, T.; Watanabe, M.; Naganuma, T.; Muramoto, K. Diversified Carbohydrate-Binding Lectins from Marine Resources. J. Amino Acids 2011, 2011, 1–20. [Google Scholar] [CrossRef]
- Nakagawa, H.; Tanigawa, T.; Tomita, K.; Tomihara, Y.; Araki, Y.; Tachikawa, E. Recent Studies on the Pathological Effects of Purified Sea Urchin Toxins. J. Toxicol. 2003, 22, 633–649. [Google Scholar] [CrossRef]
- Tateno, H.; Saneyoshi, A.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Saneyoshi, M. Isolation and Characterization of Rhamnose-binding Lectins from Eggs of Steelhead Trout (Oncorhynchus mykiss) Homologous to Low Density Lipoprotein Receptor Superfamily. J. Biol. Chem. 1998, 273, 19190–19197. [Google Scholar] [CrossRef]
- Hosono, M.; Ishikawa, K.; Mineki, R.; Murayama, K.; Numata, C.; Ogawa, Y.; Takayanagi, Y.; Nitta, K. Tandem repeat structure of rhamnose-binding lectin from catfish (Silurus asotus) eggs. Biochim. Biophys. Acta 1999, 1472, 668–675. [Google Scholar] [CrossRef]
- Gasparini, F.; Franchi, N.; Spolaore, B.; Loriano, B. Novel rhamnose-binding lectins from the colonial ascidian Botryllus schlosseri. Dev. Comp. Immunol. 2008, 32, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, T.; Ogawa, T.; Hirabayashi, J.; Kasai, K.; Muramoto, K. Isolation, characterization and molecular evolution of a novel pearl shell lectin from a marine bivalve, Pteria penguin. Mol. Divers. 2006, 10, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Edo, K.; Sakai, H.; Nakagawa, H.; Hashimoto, T.; Shinohara, M.; Ohura, K. Immunomodulatory activity of a pedicellarial venom lectin from the toxopneustid sea urchin, Toxopneustes pileolus. Toxin Rev. 2012, 31, 54–60. [Google Scholar] [CrossRef]
- Takei, M.; Nakagawa, H. A sea urchin lectin, SUL-1, from the Toxopneustid sea urchin induces DC maturation from human monocyte and drives Th1 polarization in vitro. Toxicol. Appl. Pharmacol. 2006, 213, 27–36. [Google Scholar] [CrossRef]
- Hatakeyama, T.; Ichise, A.; Unno, H.; Goda, S.; Oda, T.; Tateno, H.; Hirabayashi, J.; Sakai, H.; Nakagawa, H. Carbohydrate recognition by the rhamnose-binding lectin SUL-I with a novel three-domain structure isolated from the venom of globiferous pedicellariae of the flower sea urchin Toxopneustes pileolus. Protein Sci. 2017, 26, 1574–1583. [Google Scholar] [CrossRef]
- Ekici, O.D.; Paetzel, M.; Dalbey, R.E. Unconventional serine proteases: Variations on the catalytic Ser/His/Asp triad configuration. Protein Sci. 2008, 17, 2023–2037. [Google Scholar] [CrossRef]
- Wlodawer, A.; Li, M.; Dauter, Z.; Gustchina, A.; Uchida, K.; Oyama, H.; Dunn, B.M.; Oda, K. Carboxyl proteinase from Pseudomonas defines a novel family of subtilisin-like enzymes. Nat. Struct. Biol. 2001, 8, 442–446. [Google Scholar] [CrossRef]
- Siigur, E.; Tõnismägi, K.; Trummal, K.; Samel, M.; Vija, H.; Aaspõllu, A.; Rönnholm, G.; Subbi, J.; Kalkkinen, N.; Siigur, J. A new tyrosine-specific chymotrypsin-like and angiotensin-degrading serine proteinase from Vipera lebetina snake venom. Biochimie 2011, 93, 321–330. [Google Scholar] [CrossRef]
- Li, Q.; Barghi, N.; Lu, A.; Fedosov, A.E.; Bandyopadhyay, P.K.; Lluisma, A.O.; Concepcion, G.P.; Yandell, M.; Olivera, B.M.; Safavi-hemami, H. Divergence of the Venom Exogene Repertoire in Two Sister Species of Turriconus. Genome Biol. Evol. 2017, 9, 2211–2225. [Google Scholar] [CrossRef]
- Phuong, M.A.; Mahardika, G.N.; Alfaro, M.E. Dietary breadth is positively correlated with venom complexity in cone snails. BMC Genom. 2016, 17, 401. [Google Scholar] [CrossRef]
- Abalde, S.; Tenorio, M.J.; Afonso, C.M.L.; Zardoya, R. Conotoxin Diversity in Chelyconus ermineus (Born, 1778) and the Convergent Origin of Piscivory in the Atlantic and Indo-Pacific Cones. Genome Biol. Evol. 2018, 10, 2643–2662. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Elucidation of the molecular envenomation strategy of the cone snail Conus geographus through transcriptome sequencing of its venom duct. BMC Genom. 2012, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. Comparison of the Venom Peptides and Their Expression in Closely Related Conus Species: Insights into Adaptive Post-speciation Evolution of Conus Exogenomes. Genome Biol. Evol. 2015, 7, 1797–1814. [Google Scholar] [CrossRef] [PubMed]
- Jin, A.; Kaas, Q.; Lavergne, V.; Kubala, P.; Lewis, R.J.; Alewood, P.F. Transcriptomic Messiness in the Venom Duct of Conus miles Contributes to Conotoxin Diversity. Mol. Cell. Proteomics 2013, 12, 3824–3833. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2015, 112, E3782–E3791. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Duda, T.F., Jr. Age-related association of venom gene expression and diet of predatory gastropods. BMC Evol. Biol. 2016, 16, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.; Halai, R.; Wang, C.K.L.; Craik, D.J. ConoServer, a database for conopeptide sequences and structures. Bioinform. Appl. Note 2008, 24, 445–446. [Google Scholar] [CrossRef]
- Biggs, J.S.; Watkins, M.; Puillandre, N.; Ownby, J.; Lopez-vera, E.; Christensen, S.; Juarez, K.; Bernaldez, J.; Licea-navarro, A.; Showers, P.; et al. Molecular Phylogenetics and Evolution Evolution of Conus peptide toxins: Analysis of Conus californicus Reeve, 1844. Mol. Phylogenet. Evol. 2010, 56, 1–12. [Google Scholar] [CrossRef]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. High Conopeptide Diversity in Conus tribblei Revealed Through Analysis of Venom Duct Transcriptome Using Two High-Throughput Sequencing Platforms. Mar. Biotechnol. 2015, 17, 81–98. [Google Scholar] [CrossRef]
- Modica, M.V.; Lombardo, F.; Franchini, P.; Oliverio, M. The venomous cocktail of the vampire snail Colubraria reticulata (Mollusca, Gastropoda). BMC Genom. 2015, 16, 441. [Google Scholar] [CrossRef]
- Von Reumont, B.M.; Campbell, L.I.; Richter, S.; Hering, L.; Sykes, D.; Hetmank, J.; Jenner, R.A.; Bleidorn, C. A Polychaete’s Powerful Punch: Venom Gland Transcriptomics of Glycera Reveals a Complex Cocktail of Toxin Homologs. Genome Biol. Evol. 2014, 6, 2406–2423. [Google Scholar] [CrossRef]
- Verdes, A.; Simpson, D.; Holford, M. Are Fireworms Venomous? Evidence for the Convergent Evolution of Toxin Homologs in Three Species of Fireworms (Annelida, Amphinomidae). Genome Biol. Evol. 2017, 10, 249–268. [Google Scholar] [CrossRef]
- Gerdol, M.; Puillandre, N.; De Moro, G.; Guarnaccia, C.; Lucafo, M.; Benincasa, M.; Zlatev, V.; Manfrin, C.; Torboli, V.; Giulianini, P.G.; et al. Identification and Characterization of a Novel Family of Cysteine-Rich Peptides (MgCRP-I) from Mytilus galloprovincialis. Genome Biol. Evol. 2015, 7, 2203–2219. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Huang, X.; Madan, A. CAP3: A DNA Sequence Assembly Program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinforma. Appl. Note 2006, 22, 1658–1659. [Google Scholar] [CrossRef]
- Gorson, J.; Ramrattan, G.; Verdes, A.; Wright, M.; Kantor, Y.I.; Srinivasan, R.; Musunuri, R.; Packer, D.; Albano, G.; Qiu, W.; et al. Molecular Diversity and Gene Evolution of the Venom Arsenal of Terebridae Predatory Marine Snails. Genome Biol. Evol. 2015, 7, 1761–1778. [Google Scholar] [CrossRef]
- Altschup, S.F.; Gish, W.; Pennsylvania, T.; Park, U.; Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kaas, Q.; Yu, R.; Jin, A.; Dutertre, S.; Craik, D.J. ConoServer: Updated content, knowledge, and discovery tools in the conopeptide database. Nucl. Acids Res. 2012, 40, 325–330. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucl. Acids Res. 2019, 47, 506–515. [Google Scholar] [CrossRef]
- Sonnhammer, E.L.L.; Eddy, S.R.; Durbin, R. Pfam: A Comprehensive Database of Protein Domain Families Based on Seed Alignments. Proteins Struct. Funct. Genet. 1997, 28, 405–420. [Google Scholar] [CrossRef]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucl. Acids Res. 2013, 41, e121. [Google Scholar] [CrossRef]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Tarazona, S.; Garcia-Alcade, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential expression in RNA-seq: A matter of depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef]
- Armenteros, J.A.; Sønderby, C.K.; Sønderby, S.K.; Nielsen, H.; Winther, O. DeepLoc: Prediction of protein subcellular localization using deep learning. Bioinformatics 2017, 33, 3387–3395. [Google Scholar] [CrossRef]
- Xu, L.; Dong, Z.; Fang, L.; Luo, Y.; Wei, Z.; Guo, H.; Zhang, G.; Gu, Y.Q.; Coleman-Derr, D.; Xia, Q.; et al. OrthoVenn2: A web server for whole-genome comparison and annotation of orthologous clusters across multiple species. Nucl. Acids Res. 2019, 47, W52–W58. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation. Nucl. Acids Res. 2018, 46, 493–496. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; De Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucl. Acids Res. 2018, 46, 296–303. [Google Scholar] [CrossRef]
- Lopez-Vera, E.; Heimer de la Cotera, E.P.; Maillo, M.; Riesgo-Escovar, J.R.; Olivera, B.M.; Aguilar, M.B. A novel structural class of toxins: The methionine-rich peptides from the venoms of turrid marine snails (Mollusca, Conoidea). Toxicon 2004, 43, 365–374. [Google Scholar] [CrossRef]
- Puillandre, N.; Holford, M. The Terebridae and teretoxins: Combining phylogeny and anatomy for concerted discovery of bioactive compounds. BMC Chem. Biol. 2010, 10, 7. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability Article Fast Track. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).