Toxin Production in Soybean (Glycine max L.) Plants with Charcoal Rot Disease and by Macrophomina phaseolina, the Fungus that Causes the Disease


Abstract
:1. Introduction
2. Results and Discussion
2.1. Toxin Production in Culture by M. Phaseolina Isolates from Plants with Charcoal Rot Disease
2.2. Analysis of Toxin Levels in Tissue Samples from Soybean Plants Naturally Infected with Charcoal Rot Disease
3. Conclusions
4. Materials and Methods
4.1. Soybean Plant and Greenhouse Conditions
4.2. M. phaseolina Culture Sources
4.3. Preparation of Cell-Free Culture Extracts
4.4. Toxicity of Cell-Free Filtrates of M. phaseolina Culture Media in Soybean Leaf Disc Cultures
4.5. Toxin Standards for LC/MS Analyses
4.6. Preparation of Plant Tissue and M. phaseolina Culture Medium Extracts for LC/MS Analyses
4.7. LC/MS Analysis
4.8. Statistical Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dhingra, O.D.; Sinclair, J.B. Location of Macrophomina phaseoli on soybean plants related to culture characteristics and virulence. Phytopathology 1973, 63, 934–936. [Google Scholar] [CrossRef]
- Holliday, P.; Punithalingam, E. Macrophomina Phaseolina; No. 275; Descriptions of Pathogenic Fungi and Bacteria; CMI: Kew, Surrey, UK, 1970. [Google Scholar]
- Ghosh, T.; Biswas, M.K.; Guin, C.; Roy, P. A review on characterization, therapeutic approaches and pathogenesis of Macrophomina phaseolina. Plant Cell Biotechnol. Mol. Biol. 2018, 19, 72–84. [Google Scholar]
- Mengistu, A.; Ray, J.D.; Smith, J.R.; Paris, R.L. Charcoal rot disease assessment of soybean genotypes using a colony-forming unit index. Crop Sci. 2007, 47, 2453–2461. [Google Scholar] [CrossRef]
- Wyllie, T.D. Macrophomina phaseolina—Charcoal rot. In World Soybean Research: Proceedings of the World Soybean Research Conference; Hill, L.D., Ed.; Interstate Printers and Publishers Inc.: Danville, IL, USA, 1976; pp. 482–484. [Google Scholar]
- Bellaloui, N.; Mengistu, A.; Paris, R.L. Soybean seed composition in cultivars differing in resistance to charcoal rot (Macrophomina phaseolina). J. Agric. Sci. 2008, 146, 667–675. [Google Scholar] [CrossRef]
- Bellaloui, N.; Mengistu, A.; Zobiole, L.H.S.; Shier, W.T. Resistance to toxin-mediated fungal infection: Role of lignins, isoflavones, other seed phenolics, sugars and boron in the mechanism of resistance to charcoal rot disease in soybean. Toxin Rev. 2012, 31, 16–26. [Google Scholar] [CrossRef]
- Bowen, C.R.; Schapaugh, W.T., Jr. Relationship among charcoal rot infection, yield, and stability estimates in soybean blends. Crop Sci. 1989, 29, 42–46. [Google Scholar] [CrossRef]
- Gupta, G.K.; Sharma, S.K.; Ramteke, R. Biology, epidemiology and management of the pathogenic fungus Macrophomina phaseolina (Tassi) Goid with special reference to charcoal rot of soybean (Glycine max (L.) Merrill). J. Phytopathol. 2012, 160, 167–180. [Google Scholar] [CrossRef]
- Wrather, J.A. Soybean disease loss estimates for the southern United States, 1974 to 1994. Plant Dis. 1995, 79, 1076–1079. [Google Scholar]
- Wrather, J.A.; Koenning, S.R. Estimates of disease effects on soybean yields in the United States 2003–2005. J. Nematol. 2006, 38, 173–180. [Google Scholar]
- Illinois Soybean Association. Ideas that Elevate; Research report; Illinois Soybean Association: Bloomington, IL, USA, 2010; pp. 2–16. [Google Scholar]
- Smith, G.S.; Wyllie, T.D. Charcoal rot. In Compendium of Soybean Diseases, 4th ed.; Hartman, G.L., Sinclair, J.B., Rupe, J.C., Eds.; APS Press, The American Phytopathological Society: St. Paul, MN, USA, 1999; pp. 29–31. [Google Scholar]
- Gangopadhyay, S.; Wyllie, T.D.; Luedders, V.D. Charcoal rot disease of soybean transmitted by seeds. Plant Dis. Rep. 1970, 54, 1088–1091. [Google Scholar]
- Kaiser, W.J.; Horner, G.M. Root rot of irrigated lentils in Iran. Can. J. Bot. 1980, 58, 2549–2556. [Google Scholar] [CrossRef]
- Abawi, G.S.; Pastor-Corrales, M.A. Root Rots of Beans in Latin America and Africa; Diagnosis, Research Methodologies and Management Strategies; International Center for Tropical Agriculture (CIAT): Palmira, Cali, Columbia, 1990; p. 114. [Google Scholar]
- Francl, L.J.; Wyllie, T.D.; Rosenbrock, S.M. Influence of crop rotation on population density of Macrophomina phaseolina in soil infested with Heterodora glycines. Plant Dis. 1988, 72, 760–764. [Google Scholar] [CrossRef]
- Ghaffar, A.; Zentmyer, G.A.; Erwin, D.C. Effect of organic amendments on severity of Macrophomina phaseolina root rot of cotton. Phytopathology 1969, 59, 1267–1269. [Google Scholar]
- Mueller, J.D.; Short, B.J.; Sinclair, J.B. Effect of cropping history, cultivar, and sampling date on the internal fungi of soybean roots. Plant Dis. 1985, 69, 520–523. [Google Scholar] [CrossRef]
- Rothrock, C.S.; Kirkpatrick, T.L. The influence of winter legume cover crops on soilborne plant pathogens and cotton seedling diseases. Plant Dis. 1995, 79, 167–171. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Mahmood, I. Biological control of Meloidogyne incognita race 3 and Macrophomina phaseolina by Paecilomyces lilacinus and Bacillus subtilis alone and in combination in chickpea. Fund. Appl. Nematol. 1993, 16, 215–218. [Google Scholar]
- Todd, T.C. Soybean planting date and maturity effects on Hetrodera glycines and Macrophomina phaseolina on southeastern Kansas. Suppl. J. Nematol. 1993, 25, 731–737. [Google Scholar]
- Wyllie, T.D.; Scott, D.H. (Eds.) Charcoal rot of soybean-current status. In Soybean Diseases of the North Central Region; American Phytopathological Society: St. Paul, MN, USA, 1988; pp. 106–113. [Google Scholar]
- Kendig, S.R.; Rupe, J.C.; Scott, H.D. Effect of irrigation and soil water stress on densities of Macrophomina phaseolina in soil and roots of two soybean cultivars. Plant Dis. 2000, 84, 895–900. [Google Scholar] [CrossRef]
- Mengistu, A.; Arelli, P.A.; Bond, J.P.; Shannon, G.J.; Wrather, A.; Rupe, J.B.; Chen, P.; Little, C.R.; Canaday, C.H.; Newman, M.A.; et al. Evaluation of soybean genotypes for resistance to charcoal rot. Plant Health Prog. 2011. [Google Scholar] [CrossRef]
- Smith, G.S.; Carvil, O.N. Field screening of commercial and experimental soybean cultivars for their reaction to Macrophomina phaseolina. Plant Dis. 1997, 81, 363–368. [Google Scholar] [CrossRef]
- Mengistu, A.; Ray, J.D.; Smith, J.R.; Arelli, P.R.; Bellaloui, N.; Chen, P.; Shannon, G.; Boykin, D. Effect of charcoal rot on selected putative drought tolerant soybean genotypes and yield. Crop Prot. 2018, 105, 90–101. [Google Scholar] [CrossRef]
- Paris, R.L.; Mengistu, A.; Tyler, J.M.; Smith, J.R. Registration of soybean germplasm line DT97-4290 with moderate resistance to charcoal rot. Crop Sci. 2006, 46, 2324–2325. [Google Scholar] [CrossRef]
- Smith, J.R.; Ray, J.D.; Mengistu, A. Genotypic differences in yield loss of irrigated soybean attributable to charcoal rot. J. Crop Improv. 2018, 32, 781–800. [Google Scholar] [CrossRef]
- Dhingra, O.D.; Sinclair, J.B. Variation among isolates of Macrophomina phaseoli (Rhizoctonia bataticola) from the same soybean plant. Phytopathology 1972, 62, S1108. [Google Scholar]
- Jana, T.K.; Singh, N.K.; Koundal, K.R.; Sharma, T.R. Genetic differentiation of Charcoal rot pathogen, Macrophomina phaseolina, into specific groups using URP-PCR. Can. J. Microbiol. 2005, 51, 159–164. [Google Scholar] [CrossRef]
- Mayék-Pérez, N.; López-Castañeda, C.; González-Chavira, M.; Garcia-Espinosa, R.; Acosta-Gallegos, J.; de la Vega, O.M.; Simpson, J. Variability of Mexican isolates of Macrophomina phaseolina based on pathogenesis and AFLP genotype. Physiol. Mol. Plant Pathol. 2001, 59, 257–264. [Google Scholar] [CrossRef]
- Reyes-Franco, M.C.; Hernández-Delgado, S.; Beas-Fernández, R.; Medina-Fernández, M.; Simpson, J.; Mayek-Pérez, N. Pathogenic and genetic variability within Macrophomina phaseolina from Mexico and other countries. J. Phytopathol. 2006, 154, 447–453. [Google Scholar] [CrossRef]
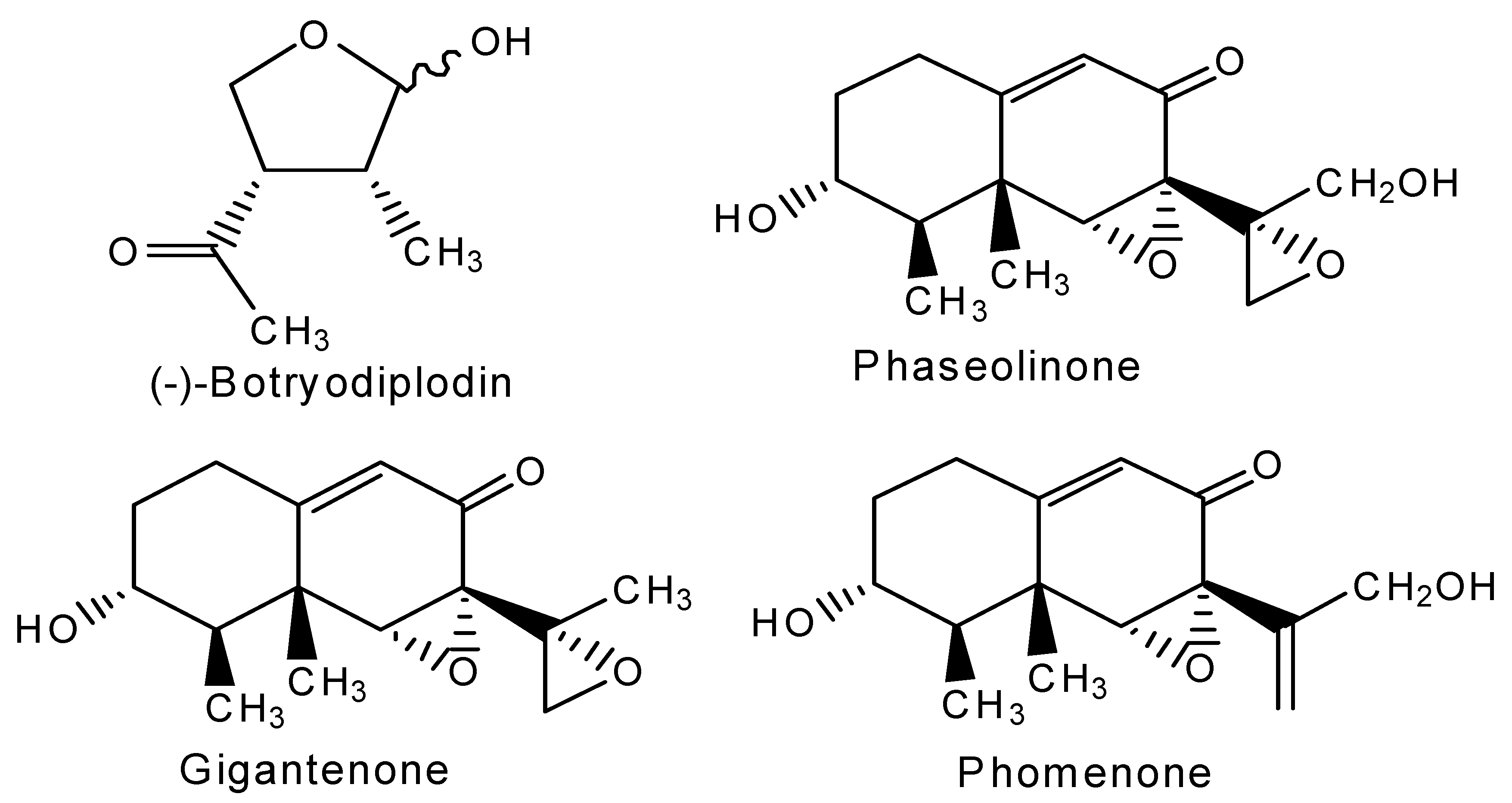
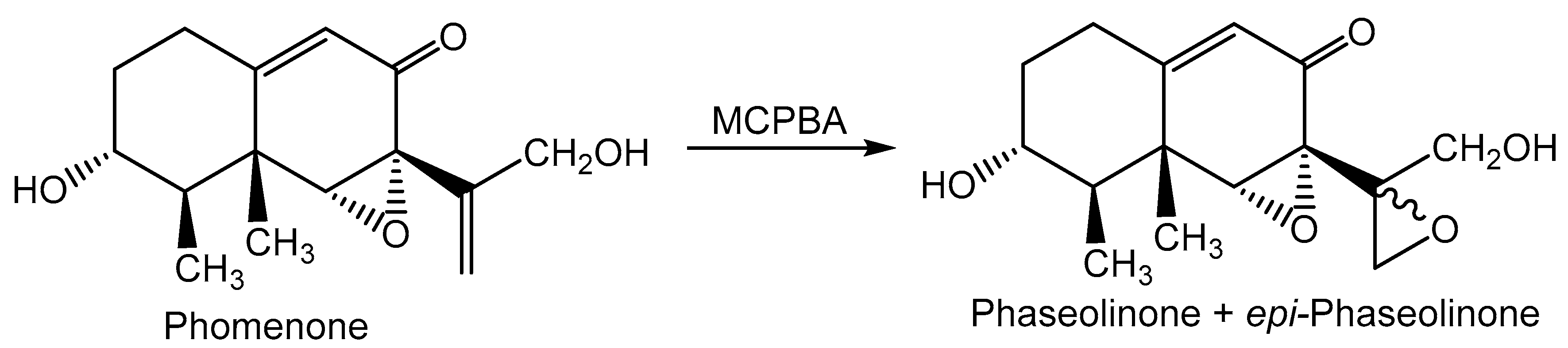
- Dhar, T.K.; Siddiqui, K.A.I.; Ali, E. Structure of phaseolinone, a novel phytotoxin from Macrophomina phaseolina. Tetrahedron Lett. 1982, 23, 5459–5462. [Google Scholar]
- Ramezani, M.; Shier, W.T.; Abbas, H.K.; Tonos, J.L.; Baird, R.E.; Sciumbato, G.L. Soybean charcoal rot disease fungus Macrophomina phaseolina in Mississippi produces the phytotoxin, (-)-botryodiplodin, but no detectable phaseolinone. J. Nat. Prod. 2007, 70, 128–129. [Google Scholar] [CrossRef]
- Shier, W.T.; Abbas, H.K.; Baird, R.E.; Ramezani, M.; Sciumbato, G.L. (-)-Botryodiplodin, a unique ribose-analog toxin. Toxin Rev. 2007, 26, 343–386. [Google Scholar] [CrossRef]
- Siddiqui, K.A.I.; Gupta, A.K.; Paul, A.K.; Banerjee, A.K. Purification and properties of a heat-resistant exotoxin produced by Macrophomina phaseolina (Tassi) Goid in culture. Experientia 1979, 35, 1222–1223. [Google Scholar] [CrossRef]
- Wrather, J.A.; Anderson, T.R.; Arsyad, D.M.; Tan, Y.; Ploper, L.D.; Porta-Puglia, A.; Ram, H.H.; Yorinori, J.T. Soybean disease loss estimates for the top ten soybean-producing countries in 1998. Can J. Plant Pathol. 2001, 23, 115–121. [Google Scholar] [CrossRef]
- Wrather, J.A.; Stienstra, W.C.; Koenning, S.R. Soybean disease loss estimates for the United States from 1996 to 1998. Can J. Plant Pathol. 2001, 23, 122–131. [Google Scholar] [CrossRef]
- Wrather, J.A.; Koenning, S.R.; Anderson, T.R. Effect of diseases on soybean yields in the United States and Ontario (1999–2002). Plant Health Prog. 2003. [Google Scholar] [CrossRef]
- Dunlap, J.R.; Bruton, B.D. Pigment biosynthesis by Macrophomina phaseolina: The glycine–specific requirement. Trans. Br. Mycol. Soc. 1986, 86, 111–115. [Google Scholar] [CrossRef]
- Shier, W.T.; Abbas, H.K.; Kotowicz, J.K.; Khambhati, V. Induction of simultaneous (-)-botryodiplodin release, microschlerotia formation and enhanced hyphal branching in Macrophomina phaseolina. Mycotoxin Res. in preparation.
- Shultz, J.L.; Ray, J.D.; Smith, J.R. Mapping two genes in the purine metabolism pathway of soybean. DNA Seq. 2008, 19, 264–269. [Google Scholar] [CrossRef]
- Abbas, H.K.; Bellaloui, N.; Butler, A.M.; Nelson, J.L.; Abou-Karam, M.; Shier, W.T. Botryodiplodin, a toxin produced by the charcoal rot disease fungus, Macrophomena phaseolina, induces phytotoxic responses in soybean (Glycine max L.) that are of types that could be used to facilitate root infection. Toxins 2019. submitted. [Google Scholar]



{kind=link}
{kind=link}
| Toxicity a in Leaf Disc Cultures of Two Soybean Genotypes | ||||||||
|---|---|---|---|---|---|---|---|---|
| DS97-84-1 | DT97-4290 | |||||||
| Isolate | Collection Site | Plant Host | 50% Strength | 100% Strength | 50% Strength | 100% Strength | Color b in One-Week Cultures | (-)-Botryodiplodin c Concentration (µg/mL) |
| Mp001A | MS | Soybean | ++ | ++ | + | + | beige | Trace |
| Mp001B | MS | Soybean | + | + | + | + | l tan | 0.19 |
| Mp004 | MS | Soybean | ++++ | ++++ | +++ | ++++ | d tan | 0.18 |
| Mp006A | MS | Soybean | ++ | ++ | + | ++ | l yellow | Trace |
| Mp006B | MS | Soybean | ++ | +++ | + | + | l tan | 0.14 |
| Mp007 | MS | Soybean | ++ | +++ | ++ | +++ | beige | 0.29 |
| Mp008A | MS | Soybean | + | + | + | + | l yellow | 0.15 |
| Mp008B | MS | Soybean | ++ | +++ | ++ | +++ | beige | 0.14 |
| Mp009A | MS | Soybean | ++ | ++ | + | + | l tan | Trace |
| Mp010B | MS | Soybean | ++ | ++ | + | + | l tan | Trace |
| Mp011A | MS | Soybean | ++ | ++ | +++ | +++ | l tan | Trace |
| Mp144 | KY | Soybean | +++ | +++ | ++ | +++ | beige | 0.18 |
| Mp146 | KY | Soybean | +++ | +++ | ++ | ++ | beige | 0.2 |
| Mp176 | AR | Soybean | + | + | + | + | l yellow | 0.08 |
| Mp178 | AR | Soybean | +++++ | +++++ | +++++ | +++++ | beige | 1.64 |
| Mp203 | LA | Soybean | +++ | +++ | +++ | +++ | tan | Trace |
| Mp204 | LA | Soybean | ++ | ++ | + | + | l tan | Trace |
| Mp214 | SD | Soybean | +++ | ++++ | +++ | +++ | l yellow | Trace |
| Mp220 | TN | Soybean | ++ | ++ | + | + | l tan | 0.17 |
| Mp223 | TX | Soybean | +++ | +++ | ++++ | ++++ | tan | 0 |
| Mp242 | ND | Soybean | + | + | + | + | l tan | 0.16 |
| Mp272 | MN | Soybean | ++++ | ++++ | ++++ | ++++ | l tan | Trace |
| Mp279 | OK | Soybean | +++++ | +++++ | ++++ | +++++ | d grey | 4.03 |
| Mp302 | KS | Soybean | +++ | +++ | ++ | ++ | l tan | 0.98 |
| Mp305 | KS | Soybean | ++++ | ++++ | +++ | +++ | l tan | Trace |
| Mp251 | NE | Dry bean | ++++ | +++++ | ++++ | +++++ | tan | 4.5 |
| Mp228 | NC | Fraser fir | + | + | + | + | l yellow | 0 |
| Mp264 | MI | Fir | +++++ | +++++ | +++++ | +++++ | tan | 6.11 |
| Mp275 | MN | Redwood | +++++ | +++++ | ++++ | +++++ | tan | Trace |
| Mp183 | FL | Strawberry | +++ | ++++ | +++ | ++++ | tan | 0.74 |
| Mp315 | AZ | Watermelon | ++++ | +++++ | ++++ | +++++ | beige | Trace |
| Mp249 | GA | Unknown | +++++ | +++++ | ++++ | ++++ | tan | 2.04 |
| Mp216 | Unknown | Unknown | ++++ | ++++ | +++ | +++ | l grey | 0.31 |
| Mp234 | Unknown | Unknown | ++++ | ++++ | ++++ | ++++ | tan | 0 |
| Mp235 | Unknown | Unknown | + | + | + | + | l tan | Trace |
| Mp238 | Unknown | Unknown | + | + | + | + | l tan | 0 |
| Mp239 | Unknown | Unknown | + | + | + | + | l tan | Trace |
| Sample Name | Charcoal Rot Disease | Soybean Tissue Type | (-)-Botryodiplodin (µg/g) a | Phomenone (µg/g) a | Gigantenone (µg/g) a | Phaseolinone (µg/g) a |
|---|---|---|---|---|---|---|
| RTS 999 302 | Yes | Roots | 0.786 | 0 | 0 | 0 |
| USG 7582 306 | Yes | Roots | 0.13 | 0 | 0 | 0 |
| DK B58-51 326 | Yes | Roots | 0.23 | 0 | 0 | 0 |
| AG 5903 324 | Yes | Roots | 0.046 | 0 | 0 | 0 |
| AG 5701 329 | Yes | Roots | 0.139 | 0 | 0 | 0 |
| PGX 5703 313 | Yes | Roots | 0.334 | 0 | 0 | 0 |
| PGY 5822 319 | Yes | Roots | 0.332 | 0 | 0 | 0 |
| ESXVT-46 328 | Yes | Roots | 0.134 | 0 | 0 | 0 |
| P 95 B96 301 | Yes | Roots | 0.006 | 0 | 0 | 0 |
| P GX 5714 311 | Yes | Roots | 0.141 | 0 | 0 | 0 |
| Garst 5812 331 | Yes | Roots | 0.209 | 0 | 0 | 0 |
| DK 5767 32 | Yes | Roots | 0.061 | 0 | 0 | 0 |
| All samples tested | Yes | Seeds | 0 | 0 | 0 | 0 |
| All samples tested | Yes | Pulp | 0 | 0 | 0 | 0 |
| All samples tested | Yes | Branches | 0 | 0 | 0 | 0 |
| All samples tested | Yes | Twigs | 0 | 0 | 0 | 0 |
| All samples tested | Yes | Leaves | 0 | 0 | 0 | 0 |
| Undiseased control b | No | Roots | 0 | 0 | 0 | 0 |
| Field and Location * | (-)-Botryodiplodin (µg/g) | Phomenone (µg/g) | Gigantenone (µg/g) | Phaseolinone (µg/g) |
|---|---|---|---|---|
| 1 KY | 0.870 | 0 | 0 | 0 |
| 417 MS | trace | 0 | 0 | 0 |
| 1 KY | 0.567 | 0 | 0 | 0 |
| 314 MS | 3.139 | 0 | 0 | 0 |
| 3 KY | 0.114 | 0 | 0 | 0 |
| 4 KY | 0.115 | 0 | 0 | 0 |
| 2 KY | 0.938 | 0 | 0 | 0 |
| 209 MS | 0.757 | 0 | 0 | 0 |
| 4 KY | 0.946 | 0 | 0 | 0 |
| 312 MS | 0.703 | 0 | 0 | 0 |
| 210 P12 MS | trace | 0 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, H.K.; Bellaloui, N.; Accinelli, C.; Smith, J.R.; Shier, W.T. Toxin Production in Soybean (Glycine max L.) Plants with Charcoal Rot Disease and by Macrophomina phaseolina, the Fungus that Causes the Disease. Toxins 2019, 11, 645. https://doi.org/10.3390/toxins11110645
Abbas HK, Bellaloui N, Accinelli C, Smith JR, Shier WT. Toxin Production in Soybean (Glycine max L.) Plants with Charcoal Rot Disease and by Macrophomina phaseolina, the Fungus that Causes the Disease. Toxins. 2019; 11(11):645. https://doi.org/10.3390/toxins11110645
Chicago/Turabian StyleAbbas, Hamed K., Nacer Bellaloui, Cesare Accinelli, James R. Smith, and W. Thomas Shier. 2019. "Toxin Production in Soybean (Glycine max L.) Plants with Charcoal Rot Disease and by Macrophomina phaseolina, the Fungus that Causes the Disease" Toxins 11, no. 11: 645. https://doi.org/10.3390/toxins11110645
APA StyleAbbas, H. K., Bellaloui, N., Accinelli, C., Smith, J. R., & Shier, W. T. (2019). Toxin Production in Soybean (Glycine max L.) Plants with Charcoal Rot Disease and by Macrophomina phaseolina, the Fungus that Causes the Disease. Toxins, 11(11), 645. https://doi.org/10.3390/toxins11110645