An overview of all mycotoxin concentrations determined per incubation time point and per treatment can be found in
Table S1. In the following paragraphs, mycotoxin concentrations are expressed relative to the maximal (free) mycotoxin concentration detected during the total duration (48 h) of the in vitro rumen simulation study. This approach facilitates presentation and interpretation of the results as mycotoxins might adsorb to feed or other particles in rumen fluid. Indeed, bacteria and yeast cell walls could be involved in adsorbing aflatoxins, ZEN, and fumonisins [
5,
6,
7,
8,
9,
10,
11,
41], while, in our former study, ENN B, ROQ-C, and ZEN adsorbed to maize silage and MPA to rumen fluid particles [
42]. Adsorbed mycotoxins are possibly released during the in vitro incubation, e.g., when feed particles are degraded. As such, mycotoxin concentrations could increase during the incubation as only the free mycotoxin fraction was determined.
2.3.1. Mycotoxins Completely Disappearing at Normal pH: DON and NIV
The final models for relative DON or NIV concentration compared to the maximal DON or NIV concentration both count nine fixed effects model parameters with replicate as random intercept and time as random slope. An overview of the parameter estimates for each model can be found in
Table 2.
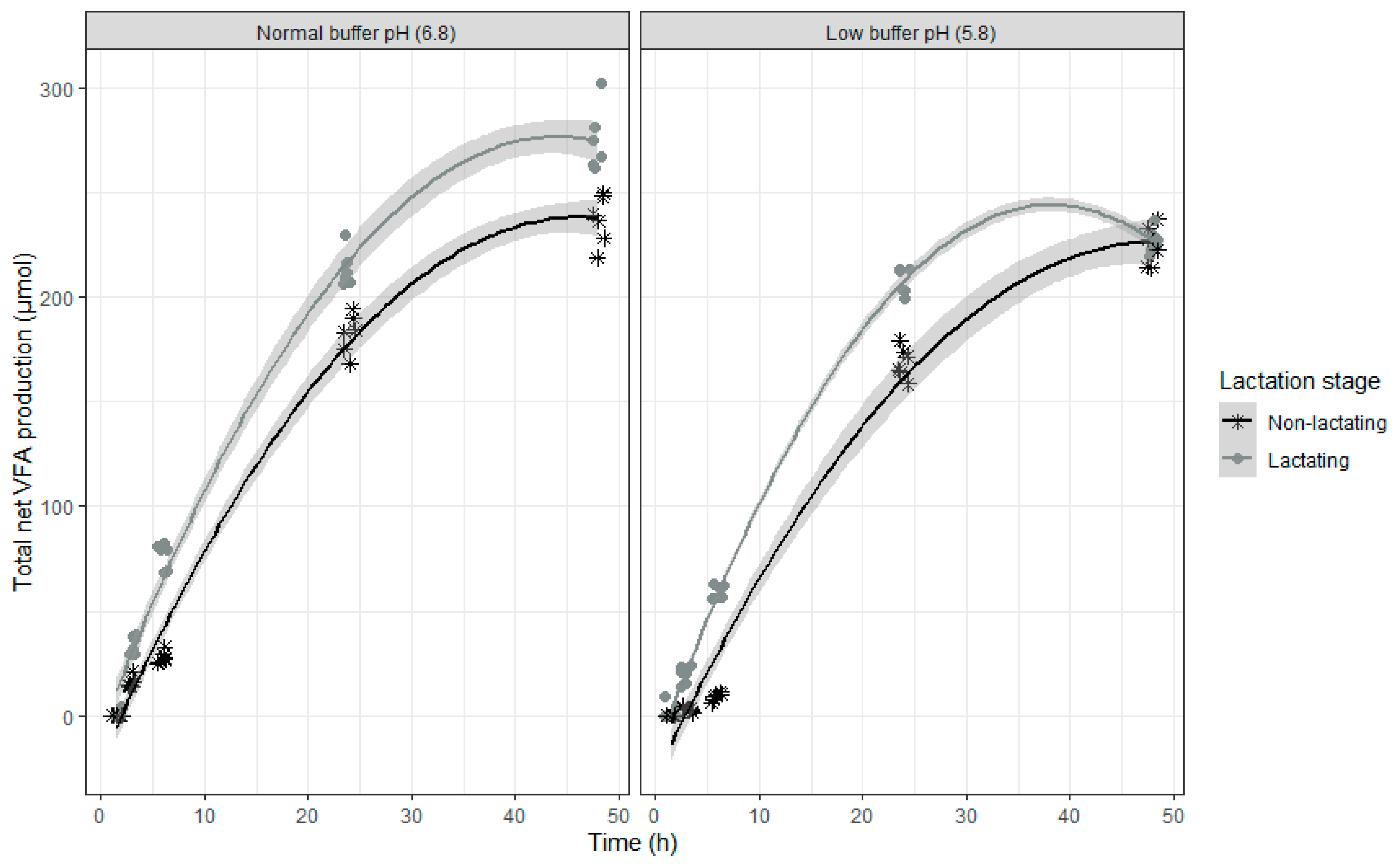
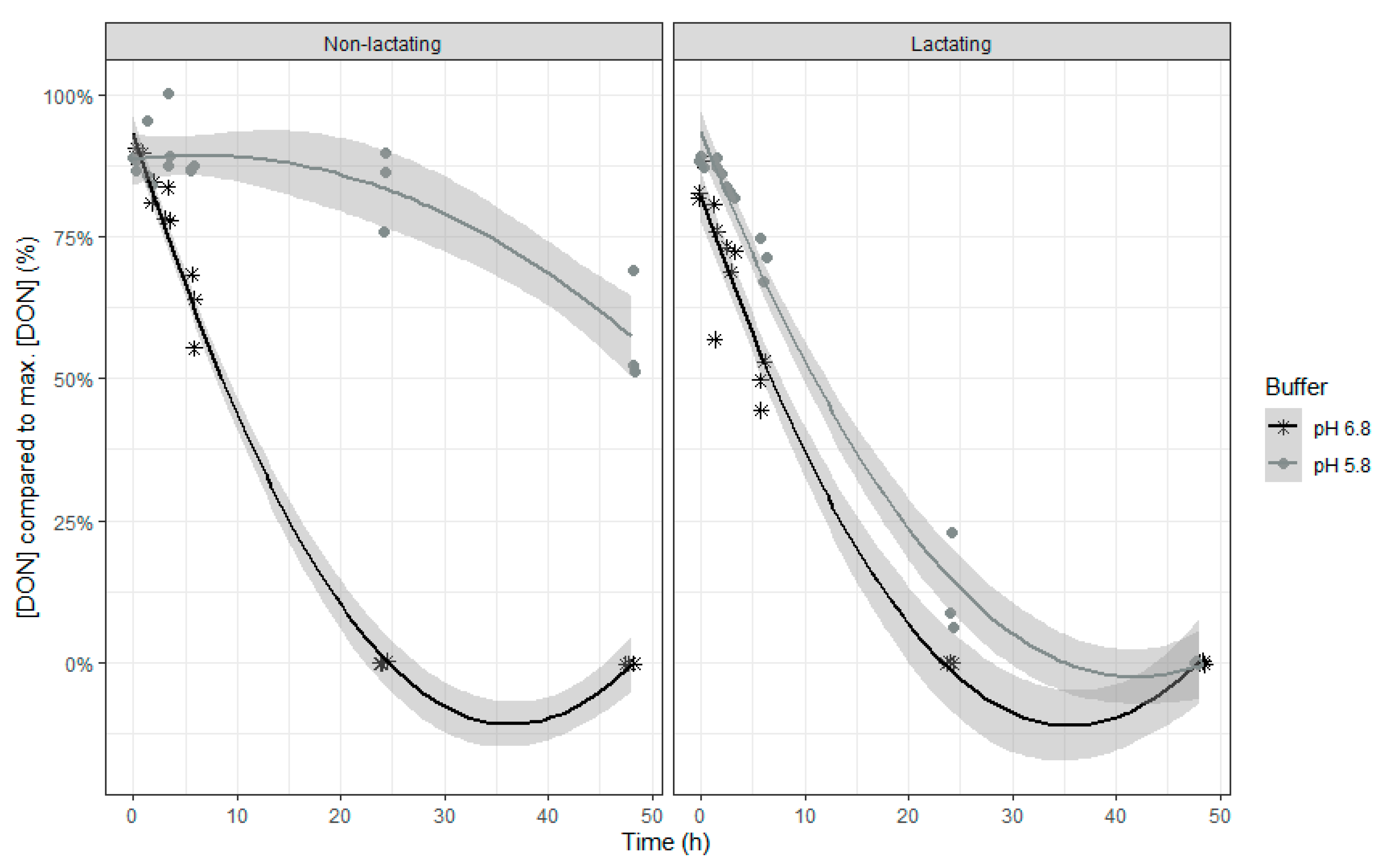
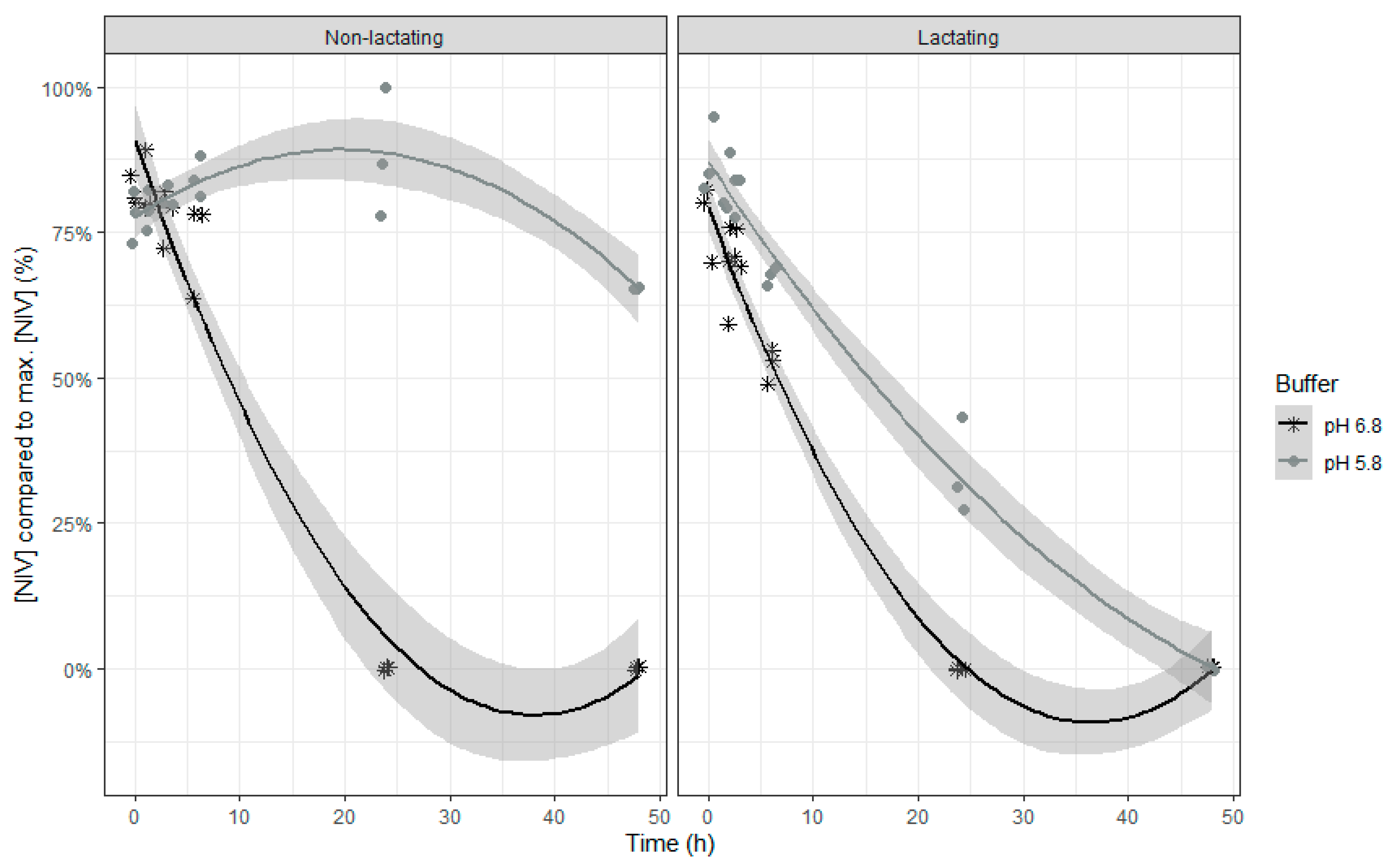
Both models show similar characteristics, which is not surprising as DON and NIV have almost the same chemical structure. When inoculum of lactating cows was used, DON and NIV concentrations generally were lower. For DON, the lower buffer pH led also to an overall higher concentration of this mycotoxin. The relative DON and NIV concentrations decrease over time, with a slower disappearance at the end of the incubation (
Table 2 and
Figure 2 (DON) and
Figure 3 (NIV)). It should be noted that negative values produced by the model are obviously irrelevant and correspond with 0%. A complete disappearance of both mycotoxins at normal pH was seen after 24 h of incubation. However, when the pH of the buffer was low, the DON and NIV disappearance was slower. Low pH particularly reduced DON and NIV disappearance when rumen fluid of non-lactating cows was used, while the negative effect was more moderate when the inoculum of lactating cows was used.
The final model for relative DOM-1 molar concentration compared to the maximal DON molar concentration counts, just like its parent compound DON, nine fixed effects model parameters with replicate as random intercept and time as random slope. An overview of the parameter estimates can be found in
Table 3.
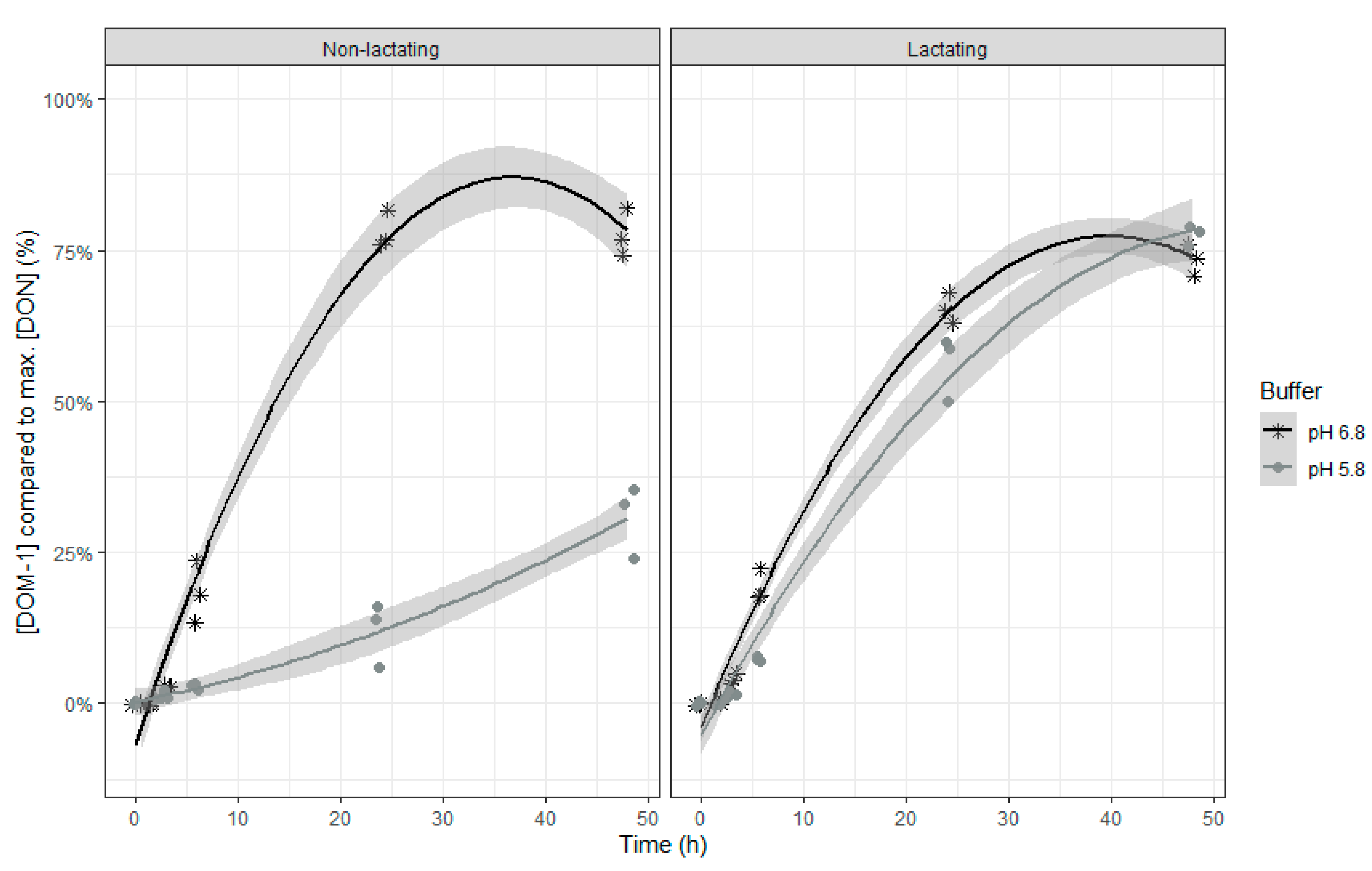
The relative DOM-1 concentration increased over time, with a slower formation at the end of the incubation (
Table 3 and
Figure 4). However, when the pH of the buffer was low, the DOM-1 formation was much slower, except when using rumen fluid of lactating cows.
The DOM-1 accumulation relative to the incubation time corresponds to the inverse of the disappearance over time of its parent compound DON for all four combinations. Except for the combination of low buffer pH and inoculum of non-lactating cows, after 48 h of incubation, DON had completely disappeared with 75% recovered as free DOM-1. After 48 h incubation at the low pH and with inoculum of non-lactating cows, DON disappearance and DOM-1 accumulation represented 40% and 30% of the maximum DON concentration, respectively. This means that in all situations about 25% of the disappeared DON is not recovered as free DOM-1. Although a full recovery was not reached in this study, the recovery of DON + DOM-1 of 75–90% is in line with other in vitro rumen studies. In a study of King et al. (1984), a total recovery of DON + DOM-1 of approximately 89% was reached [
43]. Seeling et al. (2006) also reported a total recovery of DON + DOM-1 of 80–89% in the effluent of a RUSITEC model [
24]. The conversion of DON to its far less toxic metabolite DOM-1 at normal pH is in accordance with literature [
24,
28,
43,
44]. e.g., the Genus
novus (formerly
Eubacterium) species
novus BBSH 797 of the
Coriobacteriaceae family isolated from the rumen of bovines is able to perform this detoxification and has been developed into a commercial detoxifier, Biomin
® BBSH 797 [
36,
44,
45]. EFSA approved the efficacy of this mycotoxin detoxifier in safely biotransforming trichothecenes, such as DON, into non-toxic compounds in the gastrointestinal tract of poultry and pigs, as demonstrated in numerous feeding trials [
46]. As can be seen from the results in rumen fluid, this detoxification can be hampered when the pH of the rumen is low, especially with a rumen microbial community and/or activity similar to that of a non-lactating cow inoculum. In cows with SARA, periods of rumen pH depression are seen, although this is not a constant pH depression as in the performed study. A lower detoxification of DON when pH decreases has already been described by He et al. whereby the contents of the large intestine of chickens could not detoxify DON anymore in vitro when the buffer pH was lower than 5.2 [
31]. In addition, Valgaeren et al. demonstrated a close relationship between the detoxification of DON and the functional ruminal microbiota [
14]. By means of a toxicokinetic study the authors showed that the oral bioavailability of DON is markedly increased in non-ruminating calves (50.7%) compared to ruminating calves (4.1%). Also in our study, lower microbial activity, e.g., by inoculum of non-lactating cows and at low rumen pH, impaired DON detoxification compared to the other treatments. These findings indicate that in these conditions also in ruminants DON might stay partially intact in the rumen.
Subacute ruminal acidosis is accompanied by rumenitis which leads to a higher risk of absorption of toxic compounds, such as lipopolysaccharides, through the rumen wall [
47]. Hence, in the case of SARA, it is possible that non-degraded mycotoxins are also absorbed in the rumen. In addition, the level of feed intake and feed digestibility largely influence the gastro-intestinal retention of ingesta. Hence, high yielding dairy cows that ingest high levels of highly digestible feed are more exposed to DON and other mycotoxins, as the rumen microbiota have less time to transform these toxic compounds due to a shorter retention time in the rumen. As this mycotoxin has cytotoxic effects on bovine intestinal epithelial cells (half of the maximal inhibitory concentration (IC
50) = 1.2–3.6 µM = 0.36 µg/mL–1.1 µg/mL, depending on the performed cytotoxicity test), this mycotoxin can lead to intestinal epithelial barrier disruption [
48]. Symptoms already described in ruminants related to high DON contamination of feed are gastrointestinal problems, soft stool, diarrhea, immunosuppression, decreased feed intake and a general decrease of performance [
49].
We could not confirm the detoxification of NIV into its far less toxic metabolite deepoxynivalenol (de-epoxy-NIV) in the present study as the analytical standard for this metabolite is not commercially available. However, this detoxification step can be assumed as this conversion in rumen fluid has already been described in literature whereby 78–82% of incubated NIV was transformed into de-epoxy-NIV [
25]. Just like DON, the detoxification can be hampered when the pH of the rumen is low (e.g., during periods of rumen pH depression in cows with SARA), especially when the rumen microbiota is less active (as indicated here by the lower VFA production by the inoculum of non-lactating cows). As such, intact NIV also potentially can reach the postruminal parts of the gastrointestinal tract. This mycotoxin shows an even higher cytotoxic effect on bovine intestinal epithelial cells than DON (IC
50 = 0.8–1.0 µM = 0.25 µg/mL–0.31 µg/mL, depending on the performed cytotoxicity test), which may lead to similar toxic effects [
48].
2.3.2. Mycotoxins Partially Disappearing at Normal pH: ENN B, ROQ-C and ZEN
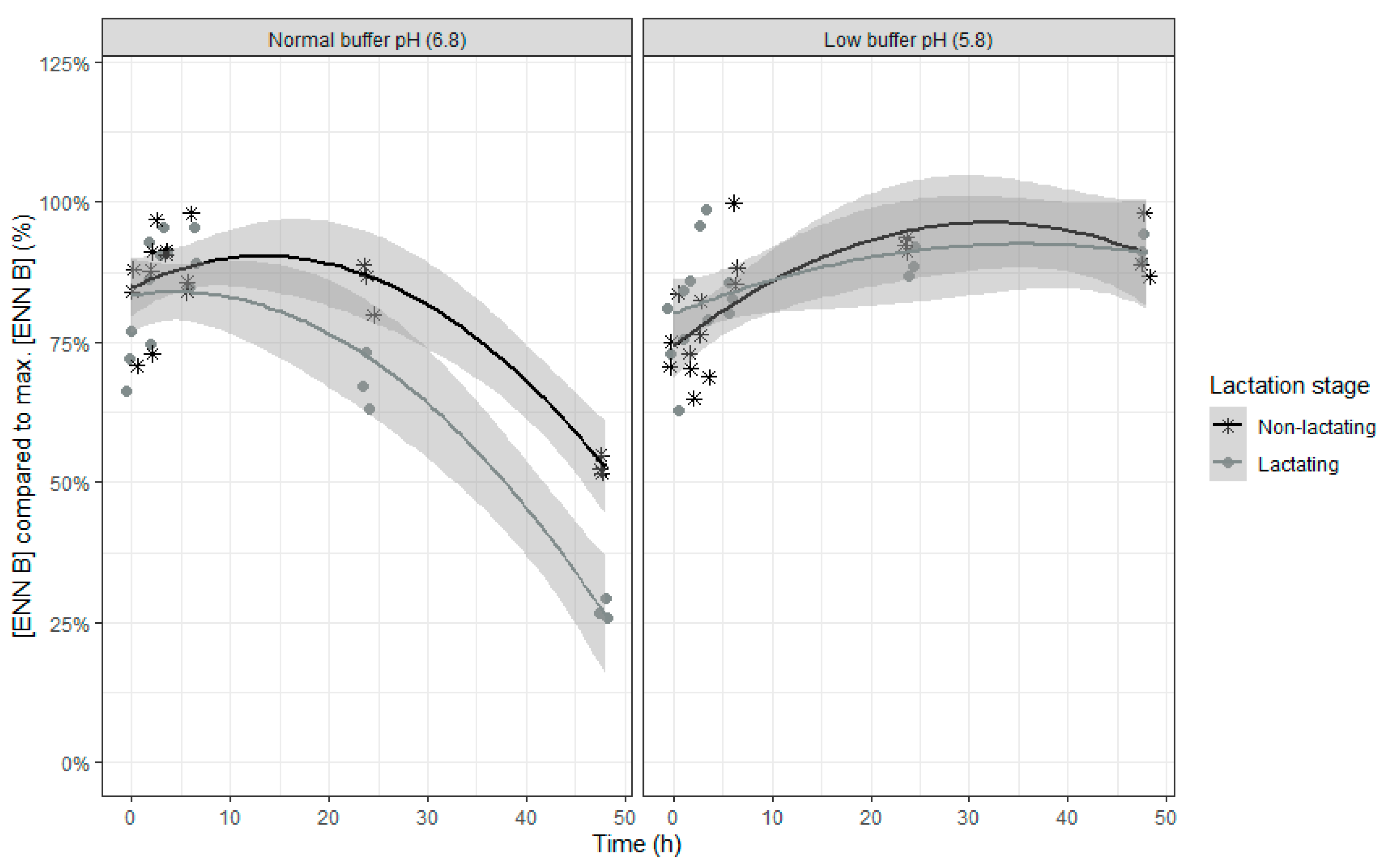
The final model for the ENN B concentration, expressed relative to the maximal ENN B concentration counts, just like DON and NIV, nine fixed effects model parameters with replicate as random intercept and time as random slope. An overview of the parameter estimates can be found in
Table 4.
An increase in the relative ENN B concentrations was seen in the beginning of the incubation, suggesting a release of adsorbed ENN B, followed by a subsequent decrease of the ENN B concentration (
Table 4 and
Figure 5). When the buffer with normal pH was used, the relative concentration of ENN B over time decreased more compared to the use of the low pH buffer.
These findings illustrate that the rumen microbiota at normal pH of 6.8 are able to degrade ENN B to a large extent, up to 72% after 48 h of incubation. However, when the rumen pH is low (as in SARA conditions), degradation of this mycotoxin is hampered. In addition, the type of rumen inoculum further determined the efficacy of the ENN B degradation with inoculum of non-lactating cows showing less degradation of the mycotoxin. As such, the rumen environment, both in terms of pH and microbial activity (as assessed by in vitro VFA production) can lead to a higher portion of intact ENN B reaching the postruminal parts of the gastrointestinal tract intact. This may lead to toxic effects towards intestinal cells due to the ionophoric properties of this mycotoxin [
50]. Cytotoxicity of ENN B already has been demonstrated in a bovine intestinal epithelial cell line with a half maximal inhibitory concentration (IC
50) for lysosomal activity at 4.0 µM (=2.6 µg/mL) and metabolic activity at 6.7 µM (=4.3 µg/mL) [
48].
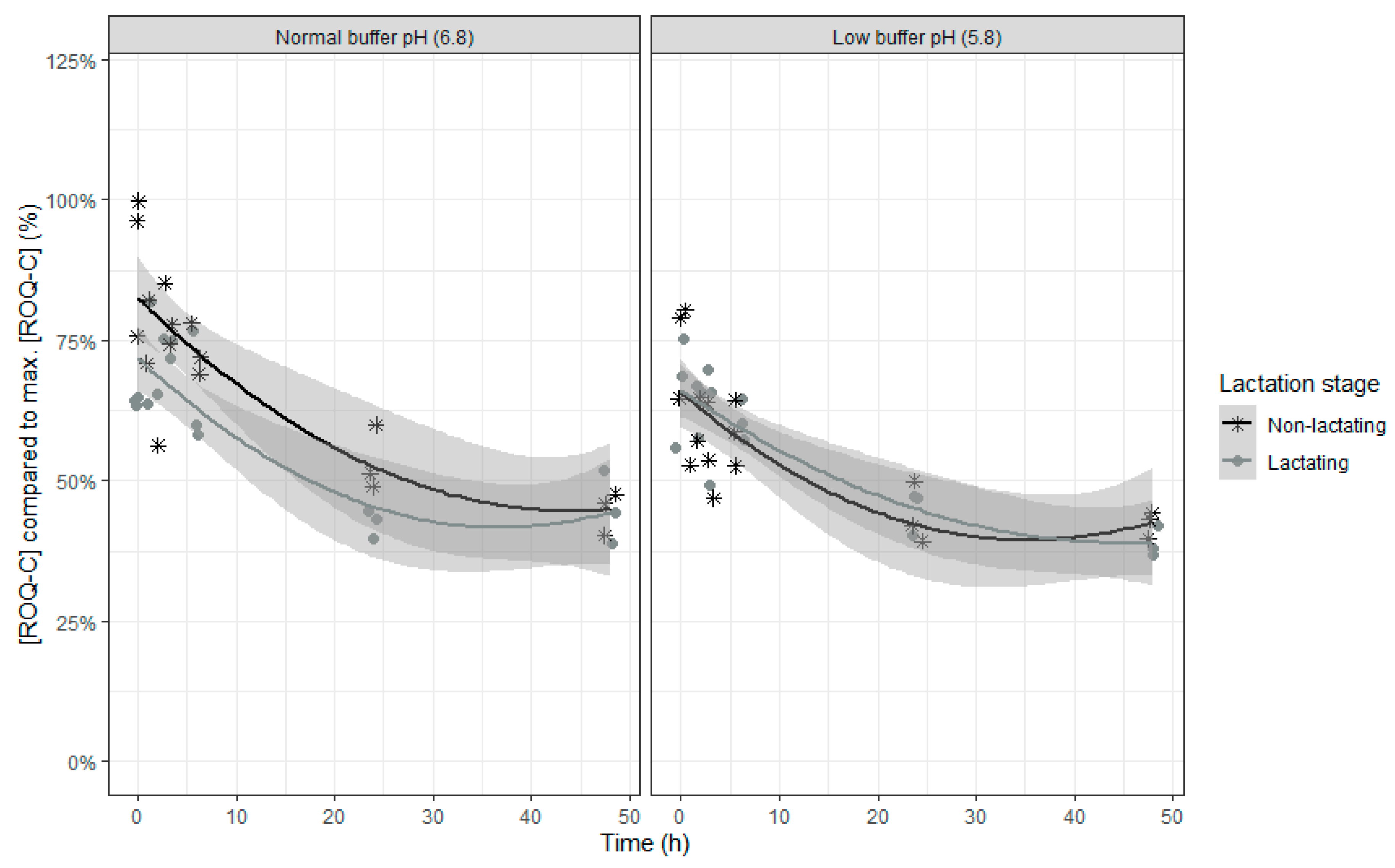
The final model for relative ROQ-C concentration compared to the maximal ROQ-C concentration counts six fixed effects model parameters and replicate as random intercept. An overview of the parameter estimates can be found in
Table 5.
In contrast to ENN B, which also shows a partial disappearance in the rumen in vitro model, no interactions with time were seen, so the model counts three fixed effects model parameters less. The relative ROQ-C concentration decreased over time, with a slower disappearance towards the end of the incubation (
Table 5 and
Figure 6). When the inoculum of lactating cows was used, a higher disappearance of ROQ-C was seen. In addition, when the inoculum of lactating cows was used, a higher difference in disappearance of ROQ-C was seen between the two pH levels compared to when the inoculum of dry cows was used.
The partial degradation of ROQ-C is consistent with literature where a recovery of more than 40% of ROQ-C was reported after 48 h of incubation [
30]. Although the inoculum of lactating cows shows more ROQ-C degradation, when the ruminal pH is low, the degradation capacity decreases. In literature, it has been described that ROQ-C can also be degraded by exposure to daylight [
51]. When ROQ-C was dissolved in ethyl acetate and kept in diffuse daylight a half-life of 50 min was reported [
51]. As the incubation flasks were not protected from daylight, the decrease in free ROQ-C concentration could be partly due to non-microbial degradation. Overall, it can be stated that the mycotoxin ROQ-C can partially reach the postruminal parts of the gastrointestinal tract. As this mycotoxin has antimicrobial properties, this can have a negative effect on the intestinal microbiota [
52,
53]. In addition, cytotoxic properties have also been demonstrated on intestinal epithelial cells. In Caco-2 cells, derived from a human colorectal adenocarcinoma, an IC
20 of 100 µM (=38.9 µg/mL) was determined when exposed for 48 h to ROQ-C [
54]. Although this mycotoxin also has neurotoxic properties when reaching the systemic circulation after absorption, paralytic symptoms are rarely seen [
55].
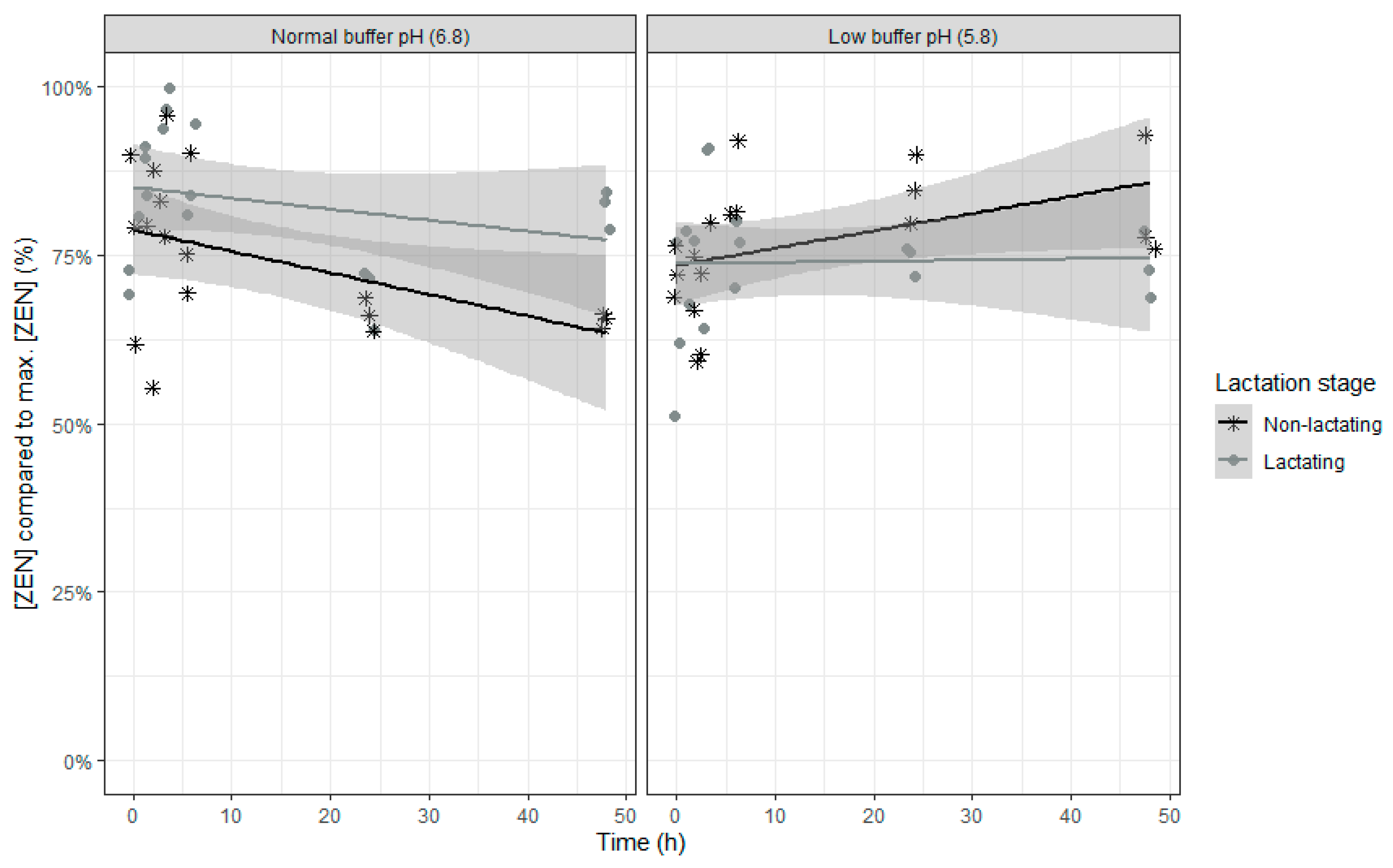
The final model for relative ZEN concentration compared to the maximal ZEN concentration counts four fixed effects model parameters with replicate as random intercept. An overview of the parameter estimates can be found in
Table 6.
For ZEN, even less fixed effects model parameters were included as time didn’t show an effect. As the trend for disappearance of ZEN over time was different at low pH compared to normal pH, the time effect disappeared. When only including the normal pH data in the model, a partial disappearance was seen over time (
p-value time effect: 0.0101). The effect of lactation stage and the interaction of lactation stage with buffer on ZEN disappearance is opposite compared with ROQ-C. The relative ZEN concentration was higher when rumen fluid of lactating cows was used (
Table 6 and
Figure 7). However, in combination with a buffer with low pH, rumen fluid of lactating cows resulted in lower relative ZEN concentrations.
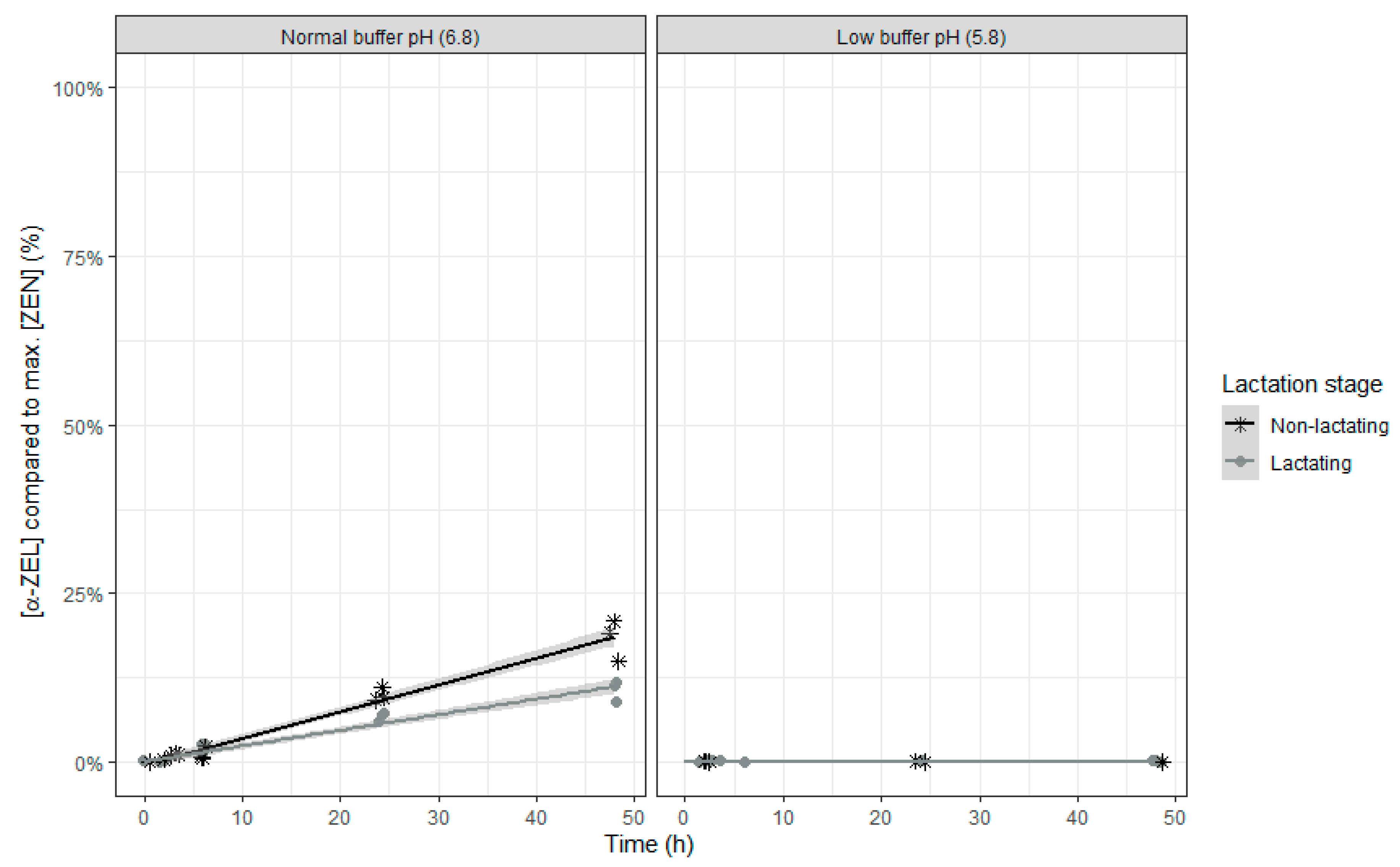
The final models for relative molar α-ZEL or β-ZEL concentration compared to the maximal molar ZEN concentration both count six fixed effects model parameters with replicate as random intercept and time as random slope. An overview of the parameter estimates can be found in
Table 7 and
Table 8.
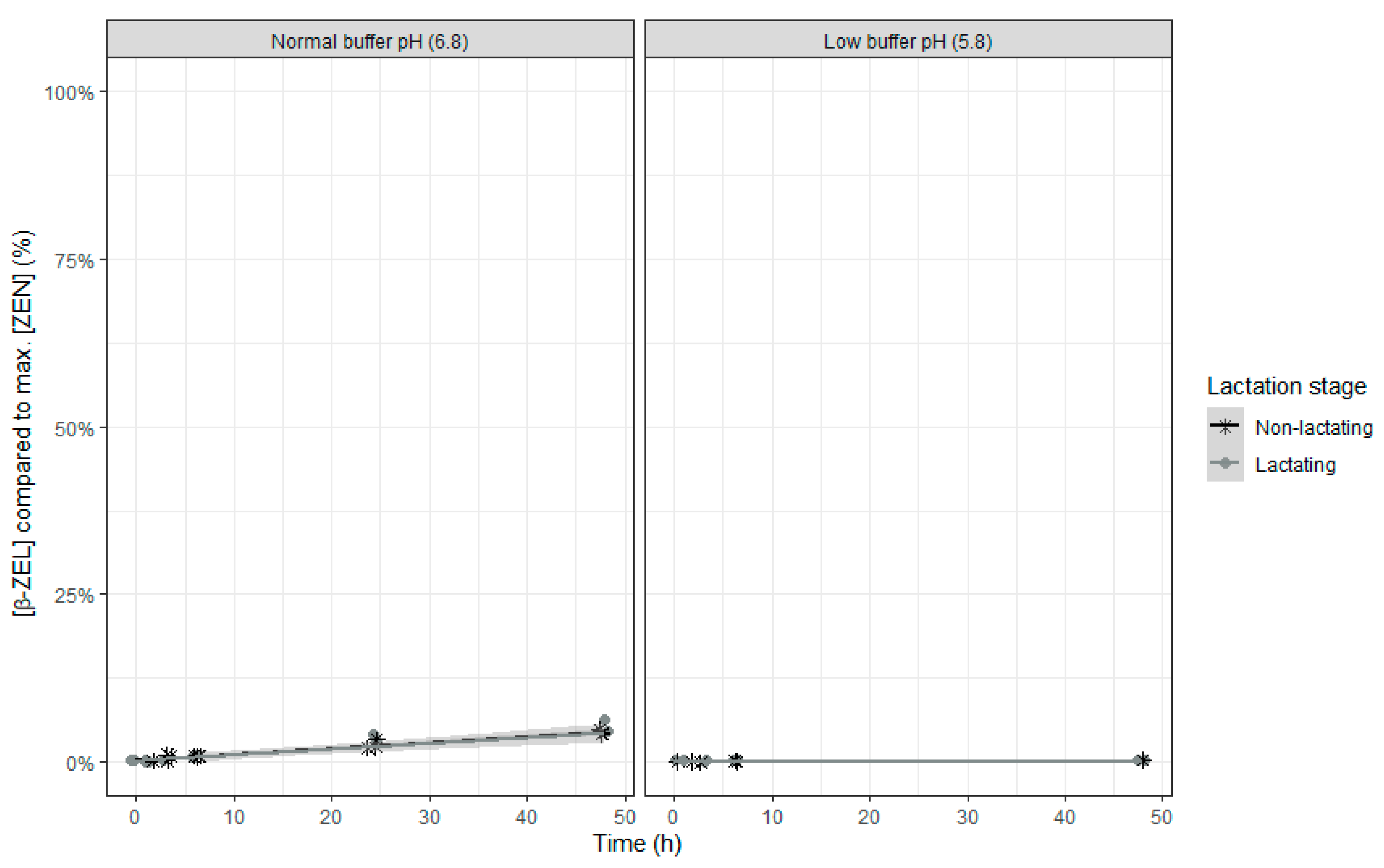
No α-ZEL or β-ZEL metabolites were formed when the rumen fluid-buffer mixture with low pH was used (
Figure 8 and
Figure 9). When the rumen fluid-buffer mixture with normal pH was applied, an increased concentration of α-ZEL and β-ZEL over time was seen (
Table 7 and
Table 8,
Figure 8 and
Figure 9). A higher formation of α-ZEL over time was seen when rumen fluid of non-lactating cows was used. By contrast, no effect of inoculum was detected on formation of the metabolite β ZEL (
Table 8 and
Figure 9).
The metabolites ZAN, α-ZAL and β-ZAL were not detected in this study, which is in accordance with literature [
28,
29].
In this study, transformation of ZEN into its metabolites α-ZEL and β-ZEL occurred only when the pH of the buffer was 6.8, although this transformation was very limited, especially for β-ZEL. Transformation of ZEN to its metabolite α-ZEL is considered an activation step as this metabolite has a higher estrogenic activity compared to ZEN, whereas transformation of ZEN to its metabolite β-ZEL is considered a deactivation step as this metabolite has a lower estrogenic activity compared to ZEN [
56]. In this study, a higher portion was transformed into α-ZEL compared to β-ZEL, which is also reported in literature [
24,
27]. Although α-ZEL is more toxic than its parent compound, its oral bioavailability is lower because of its higher polarity [
57]. Results of previously performed in vitro experiments on ruminal ZEN transformation into its metabolites α-ZEL and β-ZEL are largely variable depending on the combination of the in vitro conditions and experimental setups [
24,
27,
58,
59]. It has been shown that ZEN transformation is more pronounced when the rumen inoculum was taken directly after feeding [
59]. As the rumen inocula in this study were collected before morning feeding, this could be the reason why a relatively high amount of ZEN remained intact. In addition, ZEN and its metabolites α-ZEL and β-ZEL underlie a redox steady-state, which means that the detected ZEN concentration is not only the ruminally non-degraded ZEN, but also ZEN re-formed in the redox cycle [
58]. In vivo studies also confirm that ZEN is partially transformed by the rumen microbiota into its metabolites α-ZEL and β-ZEL, while ZAN, α–ZAL and β-ZAL were not detectable [
28,
33].
As ZEN is not completely metabolized in the rumen, ZEN can enter the intestinal tract intact. Cases of hyperoestrogenism are very rare in ruminants and occur only in calves and young heifers, after ingestion of extremely high contaminated feed or after long-term exposure to ZEN [
60,
61,
62,
63,
64]. As can be seen from this study, long-term exposure to ZEN is possible as ZEN is often found in maize silages and ZEN is not completely degraded in the rumen. The most characteristic symptoms of ZEN intoxication include swelling and hyperemia of the vulva, vagina and udder, enlargement of undeveloped uteri, feminization of young males and reproductive problems [
49].
In conclusion, ENN B, ROQ-C, and ZEN are partially degraded in the rumen at a normal pH, but a low pH and rumen inoculum of non-lactating cows negatively affects the disappearance of ENN B and ROQ-C, while ZEN even doesn’t show any transformation at low pH.
2.3.3. Mycotoxins not Disappearing at Normal pH: MPA
The final model for relative MPA concentration compared to the maximal MPA concentration counts six fixed effects model parameters and replicate as random intercept. An overview of the parameter estimates can be found in
Table 9.
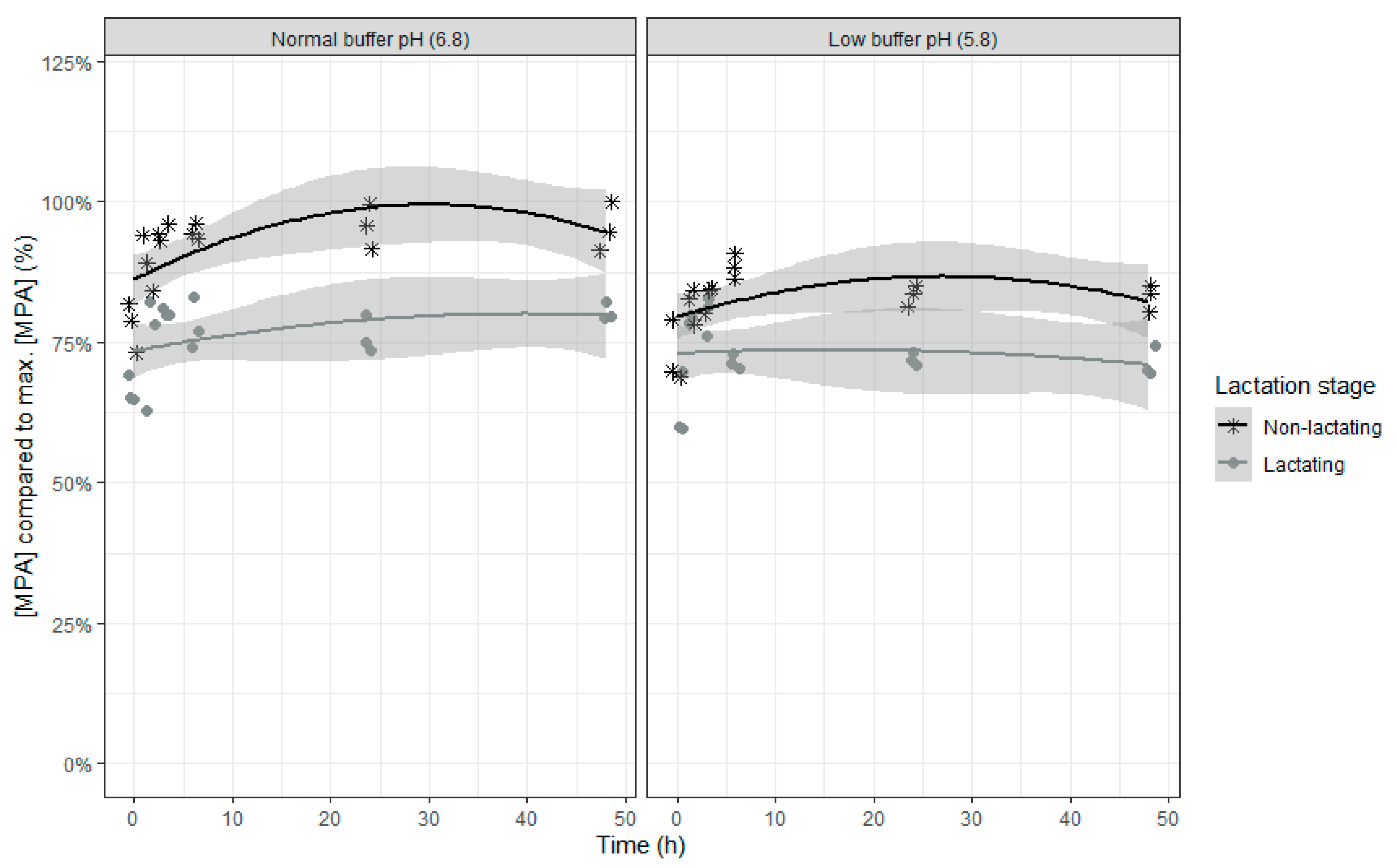
An increase in the relative MPA concentrations was seen in the beginning of the incubation, suggesting a release of bound MPA, followed by a subsequent slight decrease of the MPA concentration (
Table 9 and
Figure 10). As the change in MPA concentration over time is only minimal and differences between the four combinations already exist at the beginning of the incubation, it can be concluded that no biologically relevant microbial degradation occurs during the incubation. When the inoculum of lactating cows or a lower buffer pH was used, a lower concentration of MPA was seen, but the difference between inoculum of lactating and non-lactating cows was greater at normal compared with low pH (interaction effect between lactation stage and buffer).
As no relevant microbial degradation occurs during the incubation, differences in degree of adsorption is hypothesized whereby less free MPA was present when the pH was low. An overall lower MPA concentration was seen for the inoculum of lactating cows compared to the inoculum of non-lactating cows due to a higher microbial activity (demonstrated by the in vitro VFA production), which could explain the lower free MPA concentration. The partial recovery of MPA is consistent with literature where a recovery of more than 70% of MPA was reported after 48 h of incubation [
30]. Although the rumen pH and inoculum may have an influence on the ruminal stability of the mycotoxin, about 75% can reach the postruminal parts of the gastrointestinal tract intact. Cytotoxic properties have already been demonstrated on intestinal epithelial cells. In Caco-2 cells, an IC
20 of 82 µM (=26.3 µg/mL) has been determined when exposed for 48 h to MPA [
54]. Besides an acute toxicity, a chronic toxicity was also seen whereby MPA induced a decrease of the barrier function after 21 days of exposure [
54]. As MPA has immunosuppressive properties after absorption in the gastrointestinal tract, higher incidence of infectious diseases can be expected when ruminants ingest feed that is highly contaminated with this mycotoxin [
53,
65].

 ,
, 











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}