Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review

 ,
,  and
and 
Abstract
:1. Introduction
2. Characterized PST-Transforming Enzymes
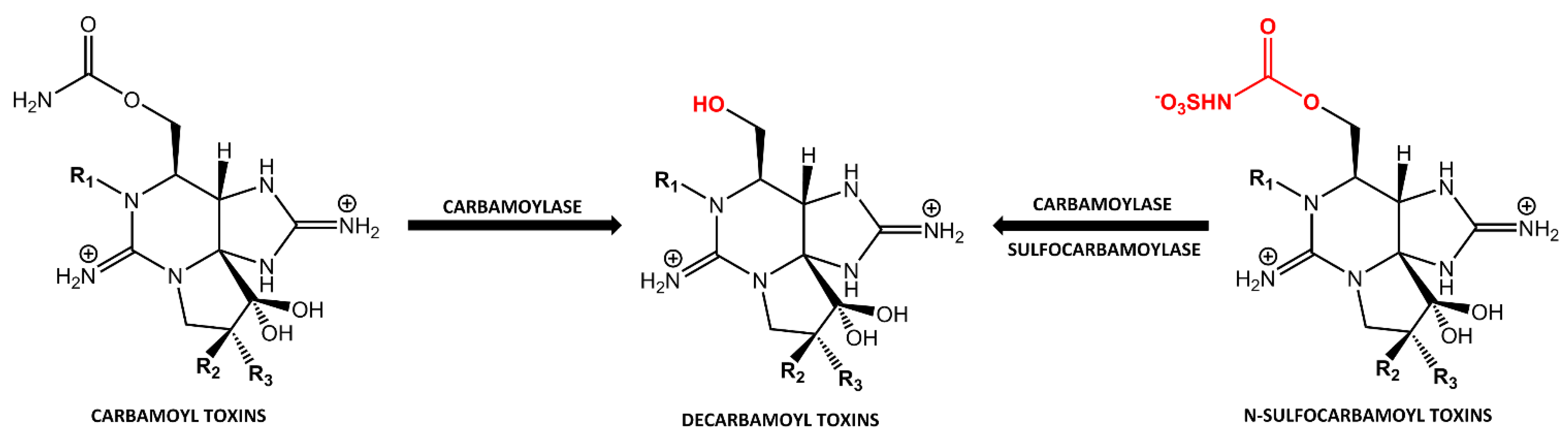
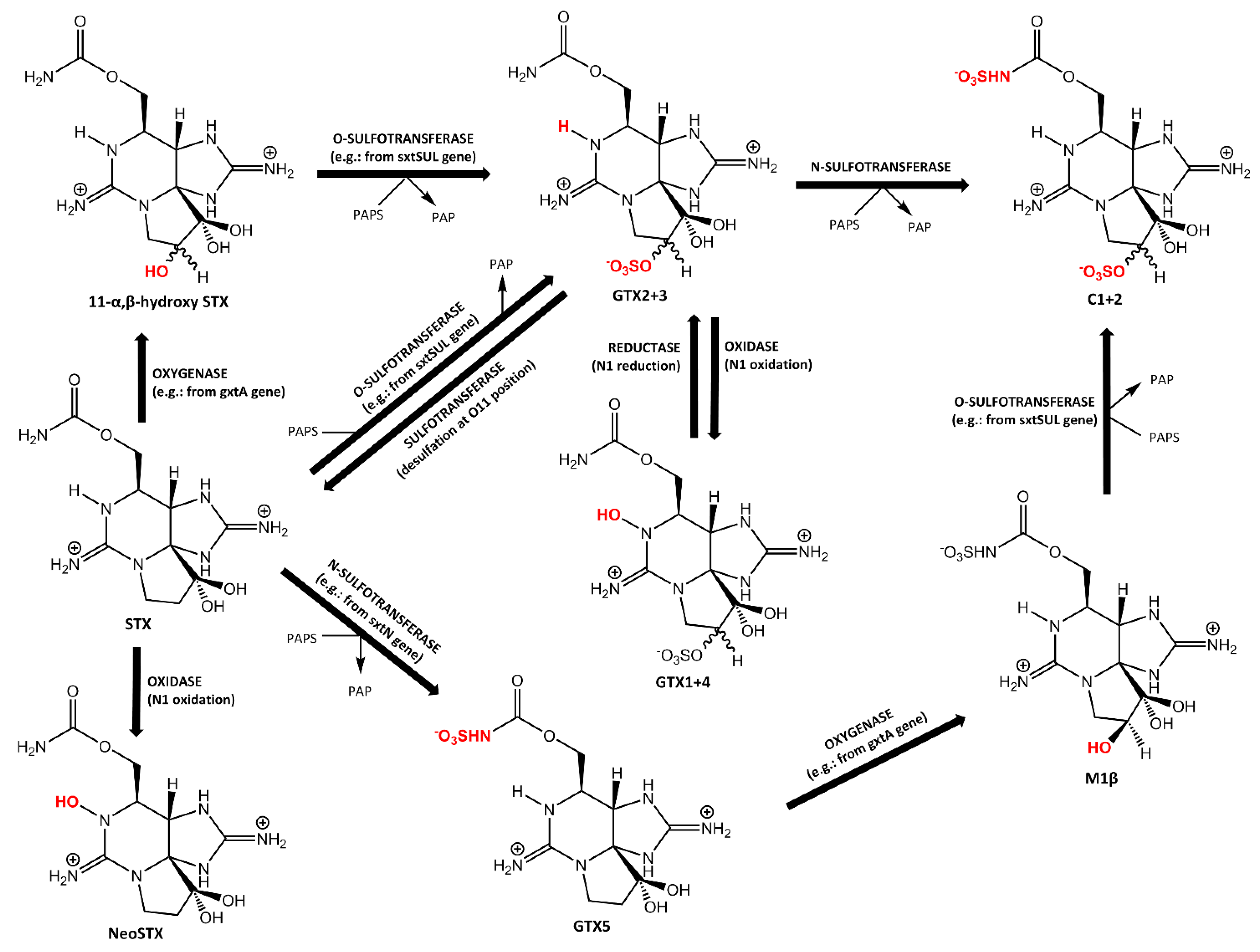
2.1. Carbamoylase and Sulfocarbamoylase
2.2. Sulfotransferases
2.2.1. Sulfotransferases Involved in PST Biosynthesis
2.2.2. Sulfotransferases Involved in PST Metabolism
2.3. Rieske Oxygenase
3. Other PST-Transforming Enzymes
3.1. Reduction at N1 Position
3.2. Oxidation at the N1 Position
3.3. Enzymes Involved in PST Detoxification
4. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Halstead, B.W.; Schantz, E.J.; World Health Organization. Paralytic Shellfish Poisoning. WHO Offset Publ. 1984, 79, 1–60. [Google Scholar]
- Schantz, E.J.; Mold, J.D.; Stranger, D.W.; Shavel, J.; Riel, F.J.; Bowden, J.P.; Lynch, J.M.; Wyler, R.S.; Riegel, B.; Sommer, H. Paralytic Shellfish Poison VI. A Procedure for the Isolation and Purification of the Poison from Toxic Clam and Mussel Tissues. J. Am. Chem. Soc. 1957, 79, 5230–5235. [Google Scholar] [CrossRef]
- Oshima, Y. Chemical and Enzymatic Transformation of Paralytic Shellfish Toxins in Marine Organisms. Harmful Mar. Algal Blooms 1995, 475–480. [Google Scholar]
- Alexander, J.; Benford, D.; Cockburn, A.; Cravedi, J.; Dogliotti, E.; Domenico, A.D.; Fernández-cruz, M.L.; Fink-gremmels, J.; Fürst, P.; Galli, C.; et al. Marine Biotoxins in Shellfish—Saxitoxin Group. EFSA J. 2009, 7, 1–76. [Google Scholar] [CrossRef]
- Negri, A.; Stirling, D.; Quilliam, M.; Blackburn, S.; Bolch, C.; Burton, I.; Eaglesham, G.; Thomas, K.; Walter, J.; Willis, R. Three Novel Hydroxybenzoate Saxitoxin Analogues Isolated from the Dinoflagellate Gymnodinium catenatum. Chem. Res. Toxicol. 2003, 16, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Onodera, H.; Satake, M.; Oshima, Y.; Yasumoto, T.; Carmichael, W.W. New Saxitoxin Analogues from the Freshwater Filamentous Cyanobacterium Lyngbya Wollei. Nat. Toxins 1997, 5, 146–151. [Google Scholar] [CrossRef]
- Vale, P. Complex Profiles of Hydrophobic Paralytic Shellfish Poisoning Compounds in Gymnodinium catenatum Identified by Liquid Chromatography with Fluorescence Detection and Mass Spectrometry. J. Chromatogr. A 2008, 1195, 85–93. [Google Scholar] [CrossRef]
- Minowa, T.; Cho, Y.; Oshima, Y.; Konoki, K.; Yotsu-Yamashita, M. Identification of a Novel Saxitoxin Analogue, 12β-Deoxygonyautoxin 3, in the Cyanobacterium, Anabaena Circinalis (TA04). Toxins 2019, 11, 539. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, J.M.; Rogart, R.B. The Binding of Saxitoxin and Tetrodotoxin to Excitable Tissue. Rev. Physiol. Biochem. Pharmacol. 1977, 79, 1–50. [Google Scholar] [CrossRef]
- Kao, C.Y.; Nishiyama, A. Actions of Saxitoxin on Peripheral Neuromuscular Systems. J. Physiol. 1965, 180, 50–66. [Google Scholar]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Cristofori-Armstrong, B.; Smith, J.J.; Chin, Y.K.Y.; Lei, J.; Zhou, Q.; et al. Structural Basis for the Modulation of Voltage-Gated Sodium Channels by Animal Toxins. Science 2018, 362, 16–18. [Google Scholar] [CrossRef] [Green Version]
- Llewellyn, L.E. Saxitoxin, a Toxic Marine Natural Product That Targets a Multitude of Receptors. Nat. Prod. Rep. 2006, 23, 200. [Google Scholar] [CrossRef]
- Vale, C.; Alfonso, A.; Vieytes, M.R.; Romarís, X.M.; Arévalo, F.; Botana, A.M.; Botana, L.M. In Vitro and in Vivo Evaluation of Paralytic Shellfish Poisoning Toxin Potency and the Influence of the PH of Extraction. Anal. Chem. 2008, 80, 1770–1776. [Google Scholar] [CrossRef]
- Oshima, Y. Postcolumn Derivatization Liquid Chromatographic Method for Paralytic Shellfish Toxins. J. AOAC Int. 1995, 78, 528–532. [Google Scholar] [CrossRef]
- Llewellyn, L.; Negri, A.; Quilliam, M. High Affinity for the Rat Brain Sodium Channel of Newly Discovered Hydroxybenzoate Saxitoxin Analogues from the Dinoflagellate Gymnodinium catenatum. Toxicon 2004, 43, 101–104. [Google Scholar] [CrossRef]
- Weigele, J.B.; Barchi, R.L. Analysis of Saxitoxin Binding in Isolated Rat Synaptosomes Using a Rapid Filtration Assay. FEBS Lett. 1978, 91, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W.; Evans, W.R.; Yin, Q.Q.; Bell, P.; Moczydlowski, E. Evidence for Paralytic Shellfish Poisons in the Freshwater Cyanobacterium Lyngbya Wollei (Farlow Ex Gomont) Comb. Nov. Appl. Environ. Microbiol. 1997, 63, 3104–3110. [Google Scholar] [CrossRef] [Green Version]
- Lagos, N.; Onodera, H.; Zagatto, P.A.; Andrinolo, D.; Azevedo, S.M.F.Q.; Oshima, Y. The First Evidence of Paralytic Shellfish Toxins in the Freshwater Cyanobacterium Cylindrospermopsis Raciborskii, Isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar] [CrossRef]
- Lafebvre, K.A.; Bill, B.D.; Erickson, A.; Baugh, K.A.; O’Rourke, L.; Costa, P.R.; Nance, S.; Trainer, V.L. Characterization of Intracellular and Extracellular Saxitoxin Levels in Both Field and Cultured Alexandrium Spp. Samples from Sequim Bay, Washington. Mar. Drugs 2008, 6, 103–116. [Google Scholar] [CrossRef]
- Oshima, Y.; Blackburn, S.I.; Hallegraeff, G.M. Comparative Study on Paralytic Shellfish Toxin of the Dinoflagellate Gymnodinium catenatum from Three Different Countries. Mar. Biol. 1993, 116, 471–476. [Google Scholar] [CrossRef]
- Usup, G.; Ahmad, A.; Matsuoka, K.; Lim, P.T.; Leaw, C.P. Biology, Ecology and Bloom Dynamics of the Toxic Marine Dinoflagellate Pyrodinium Bahamense. Harmful Algae 2012, 14, 301–312. [Google Scholar] [CrossRef]
- AOAC Official Method 2005.06. Paralytic Shellfish Poisoning Toxins in Shellfish: Prechromatographic Oxidation and Liquid Chromatography with Fluorescence Detection. J. AOAC Int. 2006. [Google Scholar]
- Botelho, M.J.; Vale, C.; Ferreira, J.G. Profiles of Paralytic Shellfish Toxins in Bivalves of Low and Elevated Toxicities Following Exposure to Gymnodinium catenatum Blooms in Portuguese Estuarine and Coastal Waters. Chemosphere 2015, 138, 1028–1036. [Google Scholar] [CrossRef]
- Fast, M.D.; Cembella, A.D.; Ross, N.W. In Vitro Transformation of Paralytic Shellfish Toxins in the Clams Mya Arenaria and Protothaca Staminea. Harmful Algae 2006, 5, 79–90. [Google Scholar] [CrossRef]
- Sullivan, J.J.; Iwaoka, W.T.; Liston, J. Enzymatic Transformation of PSP Toxins in the Littleneck Clam (Protothaca staminea). Biochem. Biophys. Res. Commun. 1983, 114, 465–472. [Google Scholar] [CrossRef]
- Cembella, A.D.; Shumway, S.E.; Larocque, R. Sequestering and Putative Biotransformation of Paralytic Shellfish Toxins by the Sea Scallop Placopecten Magellanicus: Seasonal and Spatial Scales in Natural Populations. J. Exp. Mar. Biol. Ecol. 1994, 180, 1–22. [Google Scholar] [CrossRef]
- Donovan, C.J.; Ku, J.C.; Quilliam, M.A.; Gill, T.A. Bacterial Degradation of Paralytic Shellfish Toxins. Toxicon 2008, 52, 91–100. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Shumway, S.E. Paralytic Shellfish Toxins in Bivalve Molluscs: Occurrence, Transfer Kinetics, and Biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Bricelj, V.; Lee, J.; Cembella, A.; Anderson, D.; Oshima, Y. Uptake Kinetics of Paralytic Shellfish Toxins from the Dinoflagellate Alexandrium Fundyense in the Mussel Mytilus Edulis. Mar. Ecol. Prog. Ser. 1995, 63, 475–480. [Google Scholar] [CrossRef]
- Anderson, D.M.; Sullivan, J.J.; Reguera, B. Paralytic Shellfish Poisoning in Northwest Spain: The Toxicity of the Dinoflagellate Gymnodinium catenatum. Toxicon 1989, 27, 665–674. [Google Scholar] [CrossRef]
- Lukowski, A.L.; Ellinwood, D.C.; Hinze, M.E.; Deluca, R.J.; Du Bois, J.; Hall, S.; Narayan, A.R.H. C-H Hydroxylation in Paralytic Shellfish Toxin Biosynthesis. J. Am. Chem. Soc. 2018, 140, 11863–11869. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Lewis, A.M.; O’Neil, A.; Hatfield, R.G. Transformation of Paralytic Shellfish Poisoning Toxins in UK Surf Clams (Spisula solida) for Targeted Production of Reference Materials. Toxicon 2013, 65, 41–58. [Google Scholar] [CrossRef] [PubMed]
- McCallum, M.E.; Balskus, E.P. Enzymes That Detoxify Marine Toxins. Nature 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, K.; Vilarin, N.; Botana, L.M.; Elliott, C.T. A European Perspective on Progress in Moving Away from the Mouse Bioassay for Marine-Toxin Analysis. Trends Anal. Chem. 2011, 30, 239–253. [Google Scholar] [CrossRef]
- Humpage, A.R.; Magalhaes, V.F.; Froscio, S.M. Comparison of Analytical Tools and Biological Assays for Detection of Paralytic Shellfish Poisoning Toxins. Anal. Bioanal. Chem. 2010, 397, 1655–1671. [Google Scholar] [CrossRef]
- Shimizu, Y.; Fallon, W.E.; Wekell, J.C.; Gerber, D.; Gauglitz, E.J. Analysis of Toxic Mussels (Mytilus Sp.) from the Alaskan Inside Passage. J. Agric. Food Chem. 1978, 26, 878–881. [Google Scholar] [CrossRef]
- Shimizu, Y.; Yoshioka, M. Transformation of Paralytic Shellfish Toxins as Demonstrated in Scallop Homogenates. Science 1981, 212, 547–549. [Google Scholar] [CrossRef]
- Buzy, A.; Thibault, P.; Laycock, M.V. Development of a Capillary Electrophoresis Method for the Characterization of Enzymatic Products Arising from the Carbamoylase Digestion of Paralytic Shellfish Poisoning Toxins. J. Chromatogr. A 1994, 688, 301–316. [Google Scholar] [CrossRef]
- Lin, H.P.; Cho, Y.; Yashiro, H.; Yamada, T.; Oshima, Y. Purification and Characterization of Paralytic Shellfish Toxin Transforming Enzyme from Mactra Chinensis. Toxicon 2004, 44, 657–668. [Google Scholar] [CrossRef]
- Cho, Y.; Ogawa, N.; Takahashi, M.; Lin, H.; Oshima, Y. Purification and Characterization of Paralytic Shellfish Toxin-Transforming Enzyme, Sulfocarbamoylase I, from the Japanese Bivalve Peronidia Venulosa. Biochim. Biophys. Acta Proteins Proteom. 2008, 1784, 1277–1285. [Google Scholar] [CrossRef]
- Artigas, M.L.; Vale, P.J.V.; Gomes, S.S.; Botelho, M.J.; Rodrigues, S.M.; Amorim, A. Profiles of Paralytic Shellfish Poisoning Toxins in Shellfish from Portugal Explained by Carbamoylase Activity. J. Chromatogr. A 2007, 1160, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Jaime, E.; Gerdts, G.; Luckas, B. In Vitro Transformation of PSP Toxins by Different Shellfish Tissues. Harmful Algae 2007, 6, 308–316. [Google Scholar] [CrossRef]
- Medina-Elizalde, J.; García-Mendoza, E.; Turner, A.D.; Sánchez-Bravo, Y.A.; Murillo-Martínez, R. Transformation and Depuration of Paralytic Shellfish Toxins in the Geoduck Clam Panopea Globosa From the Northern Gulf of California. Front. Mar. Sci. 2018, 5. [Google Scholar] [CrossRef]
- Botelho, M.J.; Vale, C.; Grilo, R.V.; Ferreira, J.G. Uptake and Release of Paralytic Shellfish Toxins by the Clam Ruditapes Decussatus Exposed to Gymnodinium catenatum and Subsequent Depuration. Mar. Environ. Res. 2012, 77, 23–29. [Google Scholar] [CrossRef]
- Turner, A.D.; Stubbs, B.; Coates, L.; Dhanji-Rapkova, M.; Hatfield, R.G.; Lewis, A.M.; Rowland-Pilgrim, S.; O’Neil, A.; Stubbs, P.; Ross, S.; et al. Variability of Paralytic Shellfish Toxin Occurrence and Profiles in Bivalve Molluscs from Great Britain from Official Control Monitoring as Determined by Pre-Column Oxidation Liquid Chromatography and Implications for Applying Immunochemical Tests. Harmful Algae 2014, 31, 87–99. [Google Scholar] [CrossRef]
- Cembella, A.D.; Shumway, S.E.; Lewis, N.I.I. Anatomical Distribution and Spatiotemporal Variation in Paralytic Shellfish Toxin Composition in Two Bivalve Species from the Gulf of Maine. J. Shellfish Res. 1993, 12, 389–403. [Google Scholar]
- Contreras, A.M.; Marsden, I.D.; Munro, M.H.G. Effects of Short-Term Exposure to Paralytic Shellfish Toxins on Clearance Rates and Toxin Uptake in Five Species of New Zealand Bivalve. Mar. Freshw. Res. 2012, 63, 166. [Google Scholar] [CrossRef]
- Mackenzie, L.; White, D.; Adamson, J. Temporal Variation and Tissue Localization of Paralytic Shellfish Toxins In the New Zealand Tuatua (Surfclam), Paphies Subtriangulata. J. Shellfish Res. 1996, 15, 735–740. [Google Scholar]
- Samsur, M.; Takatani, T.; Yamaguchi, Y.; Sagara, T.; NOGUCHI, T.; ARAKAWA, O. Accumulation and Elimination Profiles of Paralytic Shellfish Poison in the Short-Necked Clam Tapes Japonica Fed with the Toxic Dinoflagellate Gymnodinium catenatum. Shokuhin Eiseigaku Zasshi. 2007, 48, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Samsur, M.; Yamaguchi, Y.; Sagara, T.; Takatani, T.; Arakawa, O.; Noguchi, T. Accumulation and Depuration Profiles of PSP Toxins in the Short-Necked Clam Tapes Japonica Fed with the Toxic Dinoflagellate Alexandrium Catenella. Toxicon 2006, 48, 323–330. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Lee, J.H.; Cembella, A.D. Influence of Dinoflagellate Cell Toxicity on Uptake and Loss of Paralytic Shellfish Toxins in the Northern Quahog Mercenaria Mercenaria. Mar. Ecol. Prog. Ser. 1991, 74, 33–46. [Google Scholar] [CrossRef]
- Costa, S.T.; Vale, C.; Raimundo, J.; Matias, D.; Botelho, M.J. Changes of Paralytic Shellfish Toxins in Gills and Digestive Glands of the Cockle Cerastoderma Edule under Post-Bloom Natural Conditions. Chemosphere 2016, 149, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.-C.; Hsieh, D.P.H.; Lam, P.K.S.; Wang, W.-X. Field Depuration and Biotransformation of Paralytic Shellfish Toxins in Scallop Chlamys Nobilis and Green-Lipped Mussel Perna Viridis. Mar. Biol. 2003, 143, 927–934. [Google Scholar] [CrossRef]
- Vale, P. Fate of Benzoate Paralytic Shellfish Poisoning Toxins from Gymnodinium catenatum in Shellfish and Fish Detected by Pre-Column Oxidation and Liquid Chromatography with Fluorescence Detection. J. Chromatogr. A 2008, 1190, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Grant, F.; Ferguson, C.M.J.; Gallacher, S. Biotransformations of Paralytic Shellfish Toxins by Bacteria Isolated from Bivalve Molluscs. Appl. Environ. Microbiol. 2001, 67, 2345–2353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, C.; Bravo, M.D.C.; Lagos, M.; Lagos, N. Paralytic Shellfish Poisoning: Post-Mortem Analysis of Tissue and Body Fluid Samples from Human Victims in the Patagonia Fjords. Toxicon 2004, 43, 149–158. [Google Scholar] [CrossRef]
- Yoshida, T.; Sako, Y.; Uchida, A.; Ishida, Y.; Arakawa, O.; Noguchi, T. Purification and Properties of Paralytic Shellfish Poisoning Toxins Sulfotransferase from Toxic Dinoflagellate Gymnodinium catenatum. Harmful Toxic Algal Blooms 1996, 499–502. [Google Scholar]
- Sako, Y.; Yoshida, T.; Uchida, A.; Arakawa, O.; Noguchi, T.; Ishida, Y. Purification and Characterization of Sulfotransferase Specific to N-21 of Saxitoxin and Gonyautoxin 2 + 3 from the Toxic Dinoflagellate Gymnodinium catenatum (Dinophyceae). J. Phycol. 2001, 37, 1044–1051. [Google Scholar] [CrossRef]
- Yoshida, T.; Sako, Y.; Kakutani, T.; Fujii, A.; Uchida, A.; Ishida, Y.; Arakawa, O.; Noguchi, T. Comparative Study of Two Sulfotransferases for Sulfation to N-21 of Gymnodinium catenatum and Alexandrium Catenella Toxins. In Harmful Algae, Proceedings of the VIII International Conference on Harmful Algae, 25–29 June 1997, Vigo, Spain; Reguera, B., Blanco, J., Fernandez, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Vigo, Spain, 1998; pp. 366–369. [Google Scholar]
- Wang, D.; Zhang, S.; Hong, H. A Sulfotransferase Specific to N-21 of Gonyautoxin 2/3 from Crude Enzyme Extraction of Toxic Dinoflagellate Alexandrium Tamarense CI01. Chin. J. Oceanol. Limnol. 2007, 25, 227–234. [Google Scholar] [CrossRef]
- Yoshida, T.; Sako, Y.; Uchida, A.; Kakutani, T.; Arakawa, O.; Noguchi, T.; Ishida, Y. Purification and Characterization of Sulfotransferase Specific to O-22 of 11-Hydroxy Saxitoxin from the Toxic Dinoflagellate Gymnodinium catenatum (Dinophyceae). Fish. Sci. 2002, 68, 634–642. [Google Scholar] [CrossRef]
- Wang, D.Z.; Gao, Y.; Lin, L.; Hong, H.S. Comparative Proteomic Analysis Reveals Proteins Putatively Involved in Toxin Biosynthesis in the Marine Dinoflagellate Alexandrium Catenella. Mar. Drugs 2013, 11, 213–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soto-Liebe, K.; Murillo, A.A.; Krock, B.; Stucken, K.; Fuentes-Valdés, J.J.; Trefault, N.; Cembella, A.; Vásquez, M. Reassessment of the Toxin Profile of Cylindrospermopsis Raciborskii T3 and Function of Putative Sulfotransferases in Synthesis of Sulfated and Sulfonated PSP Toxins. Toxicon 2010, 56, 1350–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihali, T.K.; Carmichael, W.W.; Neilan, B.A. A Putative Gene Cluster from a Lyngbya Wollei Bloom That Encodes Paralytic Shellfish Toxin Biosynthesis. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Kellmann, R.; Mihali, T.K.; Young, J.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic Intermediate Analysis and Functional Homology Reveal a Saxitoxin Gene Cluster in Cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [Green Version]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the Paralytic Shellfish Toxin Biosynthesis Gene Clusters in Anabaena Circinalis AWQC131C and Aphanizomenon Sp. NH-5. BMC Biochem. 2009, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Moustafa, A.; Loram, J.E.; Hackett, J.D.; Anderson, D.M.; Plumley, F.G.; Bhattacharya, D. Origin of Saxitoxin Biosynthetic Genes in Cyanobacteria. PLoS ONE 2009, 4. [Google Scholar] [CrossRef] [Green Version]
- Cullen, A.; D’Agostino, P.M.; Mazmouz, R.; Pickford, R.; Wood, S.; Neilan, B.A. Insertions within the Saxitoxin Biosynthetic Gene Cluster Result in Differential Toxin Profiles. ACS Chem. Biol. 2018, 13, 3107–3114. [Google Scholar] [CrossRef]
- Lukowski, A.L.; Denomme, N.; Hinze, M.E.; Hall, S.; Isom, L.L.; Narayan, A.R.H. Biocatalytic Detoxification of Paralytic Shellfish Toxins. ACS Chem. Biol. 2019, 14, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Dell’Aversano, C.; Walter, J.A.; Burton, I.W.; Stirling, D.J.; Fattorusso, E.; Quilliam, M.A. Isolation and Structure Elucidation of New and Unusual Saxitoxin Analogues from Mussels. J. Nat. Prod. 2008, 71, 1518–1523. [Google Scholar] [CrossRef]
- Li, Y.; Sun, X.; Hu, X.; Xun, X.; Zhang, J.; Guo, X.; Jiao, W.; Zhang, L.; Liu, W.; Wang, J.; et al. Scallop Genome Reveals Molecular Adaptations to Semi-Sessile Life and Neurotoxins. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef]
- Ding, L.; Qiu, J.; Li, A. Proposed Biotransformation Pathways for New Metabolites of Paralytic Shellfish Toxins Based on Field and Experimental Mussel Samples. J. Agric. Food Chem. 2017, 65, 5494–5502. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Meng, F.; Ding, L.; Che, Y.; McCarron, P.; Beach, D.G.; Li, A. Dynamics of Paralytic Shellfish Toxins and Their Metabolites during Timecourse Exposure of Scallops Chlamys Farreri and Mussels Mytilus Galloprovincialis to Alexandrium Pacificum. Aquat. Toxicol. 2018, 200, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Ma, J.; Cao, J.; Wang, Q.; Yu, R.; Thomas, K.; Quilliam, M.A. Analysis of Paralytic Shellfish Toxins and Their Metabolites in Shellfish from the North Yellow Sea of China. Food Addit. Contam. Part A 2012, 29, 1455–1464. [Google Scholar] [CrossRef]
- Kotaki, Y. Screening of Bacteria Which Convert Crab A. Floridus. Nippon Suisan Gakkaishi 1989, 55, 1293. [Google Scholar] [CrossRef] [Green Version]
- Kotaki, Y.; Oshima, Y.; Yasumoto, T. Bacterial Transformation of Paralytic Shellfish Toxins in Coral Reef Crabs and a Marine Snail. Bull. Jpn. Soc. Sci. Fish. 1985, 51, 1009–1013. [Google Scholar] [CrossRef]
- Kvitek, R.; Beitler, M. Relative Insensitivity of Butter Clam Neurons to Saxitoxin; a Pre-Adaptation for Sequestering Paralytic Shellfish Poisoning Toxins as a Chemical Defense. Mar. Ecol. Prog. Ser. 1991, 69, 47–54. [Google Scholar] [CrossRef]
- Manfrin, C.; De Moro, G.; Torboli, V.; Venier, P.; Pallavicini, A.; Gerdol, M. Physiological and Molecular Responses of Bivalves to Toxic Dinoflagellates. Invertebr. Surviv. J. 2012, 9, 184–199. [Google Scholar]
- Barry, S.M.; Challis, G.L. Mechanism and Catalytic Diversity of Rieske Non-Heme Iron-Dependent Oxygenases. ACS Catal. 2013, 3, 2362–2370. [Google Scholar] [CrossRef] [Green Version]
- Murakami, R.; Yamamoto, K.; Noguchi, T. Difference in PSP Composition among Various Parts of Surf Clam. Food Hyg. Saf. Sci. 1999, 40, 55–61_1. [Google Scholar] [CrossRef]
- Chen, C.Y.; Chou, H.N. Accumulation and Depuration of Paralytic Shellfish Poisoning Toxins by Purple Clam Hiatula Rostrata Lightfoot. Toxicon 2001, 39, 1029–1034. [Google Scholar] [CrossRef]
- Tian, H.; Gao, C.; Wang, Z.; Sun, P.; Fan, S.; Zhu, M. Comparative Study on in Vitro Transformation of Paralytic Shellfish Poisoning (PSP) Toxins in Different Shellfish Tissues. Acta Oceanol. Sin. 2010, 29, 120–126. [Google Scholar] [CrossRef]
- Smith, E.A.; Mackintosh, F.H.; Grant, F.; Gallacher, S. Sodium Channel Blocking (SCB) Activity and Transformation of Paralytic Shellfish Toxins (PST) by Dinoflagellate-Associated Bacteria. Aquat. Microb. Ecol. 2002, 29, 1–9. [Google Scholar] [CrossRef]
- Lagos, N. Paralytic Shellfish Poisoning Phycotoxins: Occurrence in South America. Comments Toxicol. 2003, 9, 175–193. [Google Scholar] [CrossRef]
- García, C.; Barriga, A.; Díaz, J.C.; Lagos, M.; Lagos, N. Route of Metabolization and Detoxication of Paralytic Shellfish Toxins in Humans. Toxicon 2010, 55, 135–144. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Rodriguez-Navarro, A.; Díaz, J.C.; Torres, R.; Lagos, N. Evidence of in Vitro Glucuronidation and Enzymatic Transformation of Paralytic Shellfish Toxins by Healthy Human Liver Microsomes Fraction. Toxicon 2009, 53, 206–213. [Google Scholar] [CrossRef]
- Sies, H. Glutathione and Its Role in Cellular Functions. Free Radic. Biol. Med. 1999, 27, 916–921. [Google Scholar] [CrossRef]
- Gubbins, M.J.; Eddy, F.B.; Gallacher, S.; Stagg, R.M. Paralytic Shellfish Poisoning Toxins Induce Xenobiotic Metabolising Enzymes in Atlantic Salmon (Salmo Salar). Mar. Environ. Res. 2000, 50, 479–483. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, P.; Guilherme, S.; Barata, M.; Nicolau, L.; Santos, M.A.; Pacheco, M.; Pousão-Ferreira, P. Biotransformation Modulation and Genotoxicity in White Seabream upon Exposure to Paralytic Shellfish Toxins Produced by Gymnodinium catenatum. Aquat. Toxicol. 2012, 106–107, 42–47. [Google Scholar] [CrossRef]
- Costa, P.R.; Pereira, P.; Guilherme, S.; Barata, M.; Santos, M.A.; Pacheco, M.; Pousão-Ferreira, P. Hydroxybenzoate Paralytic Shellfish Toxins Induce Transient GST Activity Depletion and Chromosomal Damage in White Seabream (Diplodus sargus). Mar. Environ. Res. 2012, 79, 63–69. [Google Scholar] [CrossRef]
- Miners, J.O.; Mackenzie, P.I. Drug Glucuronidation in Humans. Pharmacol. Ther. 1991, 51, 347–369. [Google Scholar] [CrossRef]
- Turner, A.D.; Lewis, A.M.; Hatfield, R.G.; Galloway, A.W.; Higman, W.A. Transformation of Paralytic Shellfish Poisoning Toxins in Crassostrea Gigas and Pecten Maximus Reference Materials. Toxicon 2012, 60, 1117–1134. [Google Scholar] [CrossRef] [PubMed]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. Int. Ed. 2014, 53, 5760–5784. [Google Scholar] [CrossRef] [PubMed]
- Lattes, K.; Venegas, P.; Lagos, N.; Lagos, M.; Pedraza, L.; Rodriguez-Navarro, A.J.; García, C. Local Infiltration of Gonyautoxin Is Safe and Effective in Treatment of Chronic Tension-Type Headache. Neurol. Res. 2009, 31, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Garrido, R.; Lagos, N.; Lagos, M.; Rodríguez-Navarro, A.J.; Garcia, C.; Truan, D.; Henriquez, A. Treatment of Chronic Anal Fissure by Gonyautoxin. Color. Dis. 2007, 9, 619–624. [Google Scholar] [CrossRef]
- Hinzpeter, J.; Barrientos, C.; Zamorano, Á.; Martinez, Á.; Palet, M.; Wulf, R.; Barahona, M.; Sepúlveda, J.M.; Guerra, M.; Bustamante, T.; et al. Gonyautoxins: First Evidence in Pain Management in Total Knee Arthroplasty. Toxicon 2016, 119, 180–185. [Google Scholar] [CrossRef]
- Valenzuela, C.; Torres, C.; Muñoz, V.; Simbaina, J.C.; Sánchez, A.; Bustamante, T.; Sepúlveda, J.M.; Piron, R.; Del Campo, M.; Lagos, N. Evaluation of Neosaxitoxin as a Local Anesthetic during Piglet Castration: A Potential Alternative for Lidocaine. Toxicon 2019, 164, 26–30. [Google Scholar] [CrossRef]
- Carter, R.M.; Poli, M.A.; Pesavento, M.; Sibley, D.E.T.; Lubrano, G.J.; Guilbault, G.G. Immunoelectrochemical Biosensors for Detection of Saxitoxin and Brevetoxin. Immunomethods 1993, 3, 128–133. [Google Scholar] [CrossRef]
- Fonfría, E.S.; Vilariño, N.; Campbell, K.; Elliott, C.; Haughey, S.A.; Ben-Gigirey, B.; Vieites, J.M.; Kawatsu, K.; Botana, L.M. Paralytic Shellfish Poisoning Detection by Surface Plasmon Resonance-Based Biosensors in Shellfish Matrixes. Anal. Chem. 2007, 79, 6303–6311. [Google Scholar] [CrossRef]
- Vélez, P.; Sierralta, J.; Alcayaga, C.; Fonseca, M.; Loyola, H.; Johns, D.C.; Tomaselli, G.F.; Marbán, E.; Suaréz-Isla, B.A. A Functional Assay for Paralytic Shellfish Toxins That Uses Recombinant Sodium Channels. Toxicon 2001, 39, 929–935. [Google Scholar] [CrossRef]
- Bergantin, J.H.; Sevilla, F. Quartz Crystal Microbalance Biosensor for Saxitoxin Based on Immobilized Sodium Channel Receptors. Anal. Lett. 2010, 43, 476–486. [Google Scholar] [CrossRef]
- Campbell, K.; Haughey, S.A.; Van Den Top, H.; Van Egmond, H.; Vilariño, N.; Botana, L.M.; Elliott, C.T. Single Laboratory Validation of a Surface Plasmon Resonance Biosensor Screening Method for Paralytic Shellfish Poisoning Toxins. Anal. Chem. 2010, 82, 2977–2988. [Google Scholar] [CrossRef] [PubMed]
- Clarck, L.C.; Lyons, C. Electrode Systems for Continuous Monitoring in Cardiovascular Surgery. Ann. N. Y. Acad. Sci. 1962, 102, 29–45. [Google Scholar] [CrossRef] [PubMed]
- McArdle, F.A.; Persaud, K.C. Development of an Enzyme-Based Biosensor for Atrazine Detection. Analyst 1993, 4, 419–423. [Google Scholar] [CrossRef]
- Cheng, T.-C.; DeFrank, J.; Schoeniger, J.; Simonian, A.; Flounders, A.; Wild, J.; Grimsley, J. Enzyme-Based Biosensor for the Direct Detection of Fluorine-Containing Organophosphates. Anal. Chim. Acta 2002, 442, 15–23. [Google Scholar] [CrossRef]
- Milkani, E.; Lambert, C.R.; McGimpsey, W.G. Direct Detection of Acetylcholinesterase Inhibitor Binding with an Enzyme-Based Surface Plasmon Resonance Sensor. Anal. Biochem. 2011, 408, 212–219. [Google Scholar] [CrossRef]
- Maleki, N.; Kashanian, S.; Maleki, E.; Nazari, M. A Novel Enzyme Based Biosensor for Catechol Detection in Water Samples Using Artificial Neural Network. Biochem. Eng. J. 2017, 128, 1–11. [Google Scholar] [CrossRef]
- Raposo, M.; Botelho, M.J.; Costa, S.T.; Gomes, M.T.S.R.; Rudnitskaya, A. A Carbamoylase-Based Bioassay for the Detection of Paralytic Shellfish Poisoning Toxins. Sensors 2020, 20, 507. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
 | Basic Structure | |||||||
| Group | Toxin | R1 | R2 | R3 | R4 | TEF | MBA | KD |
| Carbamoyl | STX | H | H | H |  | 1.0 | 1.0 | 0.5 |
| NeoSTX | OH | H | H | 1.0 | ||||
| GTX1 | OH | H | OSO−3 | 1.0 | ||||
| GTX2 | H | H | OSO−3 | 0.4 | ||||
| GTX3 | H | OSO−3 | H | 0.6 | ||||
| GTX4 | OH | OSO−3 | H | 0.7 | ||||
| M2 | H | OH | H | nk | ||||
| M4 | H | OH | OH | |||||
| Decarbamoyl (dc) | dcSTX | H | H | H |  | 1.0 | ||
| dcNeoSTX | OH | H | H | 0.4 | ||||
| dcGTX1 | OH | H | OSO−3 | 0.5 | ||||
| dcGTX2 | H | H | OSO−3 | 0.2 | ||||
| dcGTX3 | H | OSO−3 | H | 0.4 | ||||
| dcGTX4 | OH | OSO−3 | H | 0.5 | ||||
| ▪ LWTX4 | H | H | H | 0.004 | ||||
| N-sulfocarbamoyl | GTX5 (B1) | H | H | H |  | 0.1 | ||
| GTX6 (B2) | OH | H | H | 0.1 | ||||
| C1 | H | H | OSO−3 | 0.02 | ||||
| C2 | H | OSO−3 | H | 0.1 | ||||
| C3 | OH | H | OSO−3 | 0.01 | ||||
| C4 | OH | OSO−3 | H | 0.1 | ||||
| M1 | H | OH | H | nk | ||||
| M3 | H | OH | OH | |||||
| Mono-hydroxybenzoate | GC1 | H | H | OSO−3 |  | 3.4–4.4 | ||
| GC2 | H | OSO−3 | H | 3.4–4.4 | ||||
| GC3 | H | H | H | 2.2 | ||||
| *GC4 | OH | H | OSO−3 | nk | ||||
| *GC5 | OH | OSO−3 | H | |||||
| *GC6 | OH | H | H | |||||
| Di-hydroxybenzoate | #GC1a | H | H | OSO−3 | Di-hydroxy-benzoate analogue | nk | ||
| #GC2a | H | OSO−3 | H | |||||
| #GC3a | H | H | H | |||||
| #GC4a | OH | H | OSO−3 | |||||
| #GC5a | OH | OSO−3 | H | |||||
| #GC6a | OH | H | H | |||||
| Sulfated-benzoate | #GC1b | H | H | OSO−3 | Sulfated-benzoate-analogue | nk | ||
| #GC2b | H | OSO−3 | H | |||||
| #GC3b | H | H | H | |||||
| #GC4b | OH | H | OSO−3 | |||||
| #GC5b | OH | OSO−3 | H | |||||
| #GC6b | OH | H | H | |||||
| Deoxydecarbamoyl (do) | doSTX | H | H | H | CH3 | nk | ||
| doGTX2 | H | H | OSO−3 | |||||
| doGTX3 | H | OSO−3 | H | |||||
| Acetate | ▪ LWTX1 | H | H | OSO−3 |  | 0.07 | ||
| LWTX2 | H | H | OSO−3 | 0.004 | ||||
| LWTX3 | H | OSO−3 | H | 0.02 | ||||
| LWTX5 | H | H | H | 0.14 | ||||
| ▪ LWTX6 | H | H | H | 0.004 | ||||
| ST Type | N-ST | O-ST | ||
|---|---|---|---|---|
| Dinoflagellate species | G. catenatum | A. catenella | A. tamarense | G. catenatum |
| Optimal activity pH | 6 | 6 | 6 | 6 |
| Optimal activity temperature | 25 °C | 15 °C | 15 °C | 35 °C |
| Molecular mass | 59 kDa | 60 kDa | nk | 65 kDa |
| Structure | monomeric | nk | nk | monomeric |
| Cation requirement | Mg2+, Co2+ (enhanced effects) | none | none | none |
| Sulfate Donor | PAPS | PAPS | PAPS, DMSO, MgSO4 | PAPS |
| Reactions | GTX2+3 → C1+2 STX → GTX5 | GTX2+3 → C1+2 STX → GTX5 | GTX2+3 → C1+2 | 11-α,β-hydroxy STX → GTX2+3 |
| Enzyme/Enzymatic Mediated Transformations | Organism | References |
|---|---|---|
| Carbamoylase | Mactra chinensis | [3,39] |
| Protothaca staminea | [24,25,38] | |
| Spisula solidissima | [46] | |
| Spisula solida | [23,32,41,54] * | |
| Ruditapes decussatus | [44,54] * | |
| Scrobicularia plana | [41,54] * | |
| Paphies donacina | [47] | |
| Paphies subtriangulata | [48] | |
| Panopea globosa | [43] | |
| Tapes japonica | [46,47] | |
| Plactopecten magellanicus | [26,46] | |
| Mercenaria mercenaria | [51] | |
| Cerastoderma edule | [52,54] * | |
| Solen grandis | [39] | |
| Panope japonica | [39] | |
| Patinopecten yessoensis | [39] | |
| Perna viridis | [53] | |
| Chalmys nobilis | [53] | |
| Mytilus galloprovincialis | [54] * | |
| Venerupis pullastra | [54] * | |
| Crassostrea japonica | [54] * | |
| Donax trunculus | [54] * | |
| Chamelea gallina | [54] * | |
| Gram-negative bacteria (isolated from C. edule) | [55] | |
| Humans | [56] | |
| Sulfocarbamoylase | Peronidia venulosa | [3,40] |
| Spisula solida | [32] | |
| Mya arenaria | [32] | |
| N-sulfotransferase-PST synthesis | Gymnodinium catenatum | [3,57,58] |
| Alexandrium catennela | [59] | |
| Alexandrium tamarense | [60] | |
| Cylindrospermopsis raciborskii T3 | [63,65], | |
| Raphidiopsis brookii D9 | [63] | |
| Anabaena circinalis | [67,68] | |
| Aphanizomenon sp. Nostocales | [67,68] | |
| Scytonema crispum | [68] | |
| Microseira wollei | [64] | |
| O-sulfotransferase-PST synthesis | Gymnodinium catenatum | [57,58,61] |
| Cylindrospermopsis raciborskii T3 | [63,65], | |
| Raphidiopsis brookii D9 | [63] | |
| Anabaena circinalis | [67,68] | |
| Aphanizomenon sp. Nostocales | [66,67] | |
| Scytonema crispum | [68] | |
| Microseira wollei | [64] | |
| N-sulfotransferase-PST metabolism | Mytilus edulis | [29] |
| Plactopecten magellanicus | [37] | |
| Panopea globosa | [43] | |
| Chlamys farreri | [71,72,73] | |
| Mytilus galloprovincialis | [72,73] | |
| Patinopecten yessoensis | [74] | |
| Saxidomus purpuratus | [74] | |
| Pseudomonas sp. and Vibrio sp. (isolated from A. floridus and T. argyrostoma) | [75,76] | |
| Rieske oxygenases | Microseira wollei | [31,69] |
| Reduction at N1 position | Placopecten magellanicus | [37] |
| Pseudocardium sachalinensis | [80] | |
| Hiatula rostrata | [81] | |
| Chlamys farreri | [82] | |
| Bacteria (isolated from M. edulis and E. arcuatus) | [55] | |
| Pseudomonas sp. and Vibrio sp. (isolated from A. floridus and T. argyrostoma) | [76] | |
| α- and γ-Proteobacteria (isolated from dinoflagellates) | [83] | |
| Oxidation at N1 position | Alexandrium tamarense | [3] |
| Cylindrospermopsis raciborskii T3 | [64,65,66] | |
| Aphanizomenon flosaquae NH-5 | [64,65,66] | |
| Microseira wollei | [64,65,66] | |
| Ahrensia sp. and Caulobacter sp. (isolated from dinoflagellates) | [83] | |
| Humans | [56,85,86] | |
| Glutathione-S-transferase | Salmo salar | [88] |
| Diplodus sargus | [89,90] | |
| Glucuronosyltransferases | Humans | [84,85] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raposo, M.I.C.; Gomes, M.T.S.R.; Botelho, M.J.; Rudnitskaya, A. Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review. Toxins 2020, 12, 344. https://doi.org/10.3390/toxins12050344
Raposo MIC, Gomes MTSR, Botelho MJ, Rudnitskaya A. Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review. Toxins. 2020; 12(5):344. https://doi.org/10.3390/toxins12050344
Chicago/Turabian StyleRaposo, Mariana I. C., Maria Teresa S. R. Gomes, Maria João Botelho, and Alisa Rudnitskaya. 2020. "Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review" Toxins 12, no. 5: 344. https://doi.org/10.3390/toxins12050344
APA StyleRaposo, M. I. C., Gomes, M. T. S. R., Botelho, M. J., & Rudnitskaya, A. (2020). Paralytic Shellfish Toxins (PST)-Transforming Enzymes: A Review. Toxins, 12(5), 344. https://doi.org/10.3390/toxins12050344